Ассимиляция винограда
- биология
Ассимиляция (от латинского assimilatio — уподобление, отождествление), анаболизм, процесс образования веществ в растениях, при котором организм воспринимает извне различные элементы среды, превращая их в ассимиляты. Реакции процесса ассимиляции тесно связаны и сопряжены с таковыми процесса диссимиляции, составляя две стороны единого процесса обмена веществ и энергии всякого живого организма. Эта связь проявляется не только в слаженности и строгой последовательности слагающих их биохимических реакций, но и в энергии, которая необходима для осуществления первичного синтеза и дальнейшего превращения органических соединений. Одной из важнейших энергетических систем, обусловливающих сопряженность обменных реакций в виноградном растении, является наличие аденозинфосфорных кислот. Различают 3 наиболее важных показателя, характеризующих процесс ассимиляции.
Ассимиляционная депрессия — уменьшение интенсивности ассимиляции под влиянием неблагоприятных факторов внешней среды и внутреннего состояния растения; понижение интенсивности фотосинтеза и повышение интенсивности дыхания, при котором уровень газообмена углекислоты растения на свету близок или ниже компенсационной точки. Частая по периодичности и длительная по продолжительности ассимиляционная депрессия отрицательно сказывается на общей продуктивности виноградного растения. Ассимиляционная депрессия наблюдается в условиях жаркого климата, с резким и сильным повышением и понижением температуры, при атмосферной и почвенной засухе, понижении уровня водообеспеченности растений. У листьев винограда она наблюдается при температуре 30°-35°С, зависит от степени адаптации растения к факторам внешней среды, его физиологического состояния. Ассимиляционная депрессия у старых листьев винограда выражена сильнее, чем у молодых и ксероморфных, у которых она проявляется слабее или может отсутствовать. У виноградных насаждений, культивируемых при орошении, даже кратковременное ухудшение водного режима приводит к сильной ассимиляционной депрессии. Ярковыраженная ассимиляционная депрессия наблюдается при резкой смене светового и температурного режимов, при выносе привитых черенков из условий недостаточного освещения на интенсивный солнечный свет и др.
Частая по периодичности и длительная по продолжительности ассимиляционная депрессия отрицательно сказывается на общей продуктивности виноградного растения. Ассимиляционная депрессия наблюдается в условиях жаркого климата, с резким и сильным повышением и понижением температуры, при атмосферной и почвенной засухе, понижении уровня водообеспеченности растений. У листьев винограда она наблюдается при температуре 30°-35°С, зависит от степени адаптации растения к факторам внешней среды, его физиологического состояния. Ассимиляционная депрессия у старых листьев винограда выражена сильнее, чем у молодых и ксероморфных, у которых она проявляется слабее или может отсутствовать. У виноградных насаждений, культивируемых при орошении, даже кратковременное ухудшение водного режима приводит к сильной ассимиляционной депрессии. Ярковыраженная ассимиляционная депрессия наблюдается при резкой смене светового и температурного режимов, при выносе привитых черенков из условий недостаточного освещения на интенсивный солнечный свет и др.
Ассимиляционная сила — совокупность конечных, устойчивых продуктов световой стадии фотосинтеза: аденозинтрифосфата (АТФ) и никотинамидадениндинуклеотидфосфата (НАДФ . Н2), которые синтезируются в мембранах хлоропластов, диффундируют в матрикс, используются в реакциях усвоения углерода. У виноградного растения на усвоение одной молекулы СО2 затрачиваются 3 молекулы АТФ и 2 молекулы НАДФ . Н2. Количеств, их отношение существенно влияет на соотношение продуктов фотосинтеза. При экстремальных условиях в пластидах виноградного листавозникает дефицит АТФ, что приводит к снижению синтеза высокополимерных и увеличению мономерных соединений. Ассимиляционное число — отношение между интенсивностью фотосинтеза и содержанием хлорофилла в листе. Оно выражается соотношением: мг СО2, поглощенного листом мг хлорофилла в час и используется при оценке фотобиологической активности хлорофилла, особенно если расчет выполнен в молярных показателях (моль СО2/ моль хлорофилла в 1 ч). У виноградного растения ассимиляционное число равно 4-8, величина которого меняется в течение вегетации растений. Экологические факторы и агротехнические мероприятия оказывают влияние на уровень и направленность процессов ассимиляции и на распределение ассимилятов между вегетативными и репродуктивными органами виноградного растения, определяя величину и качество биологического и хозяйственного урожаев.
У виноградного растения ассимиляционное число равно 4-8, величина которого меняется в течение вегетации растений. Экологические факторы и агротехнические мероприятия оказывают влияние на уровень и направленность процессов ассимиляции и на распределение ассимилятов между вегетативными и репродуктивными органами виноградного растения, определяя величину и качество биологического и хозяйственного урожаев.
- Назад
- Вперед
- Вы здесь:
- Главная
- Словарь
- Ассимиляция
Еще почитать:
- Прохождение фаз вегетации столовыми сортами, Новочеркасск, 2010
- Дыхание
- Фотосинтез
- Формирование почек и соцветий винограда
- Лист
Поиск и метки, Контакты, Форум-виноград, Товары по виноградарству.
© Перепечатка и цитирование — только с активной гиперссылкой на сайт о винограде, в бумажных изданиях — только после согласования.
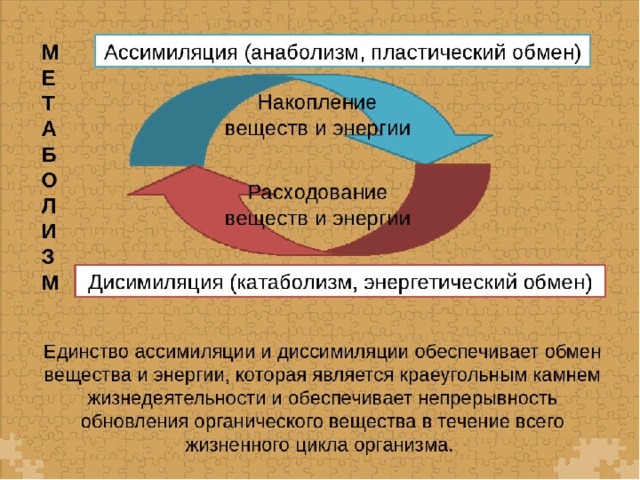
Энергетический обмен веществ и его этапы. Ассимиляция и диссимиляция
Обмен веществ и его типы
Определение 1
Обмен веществ и энергии в живых организмах называется метаболизмом.
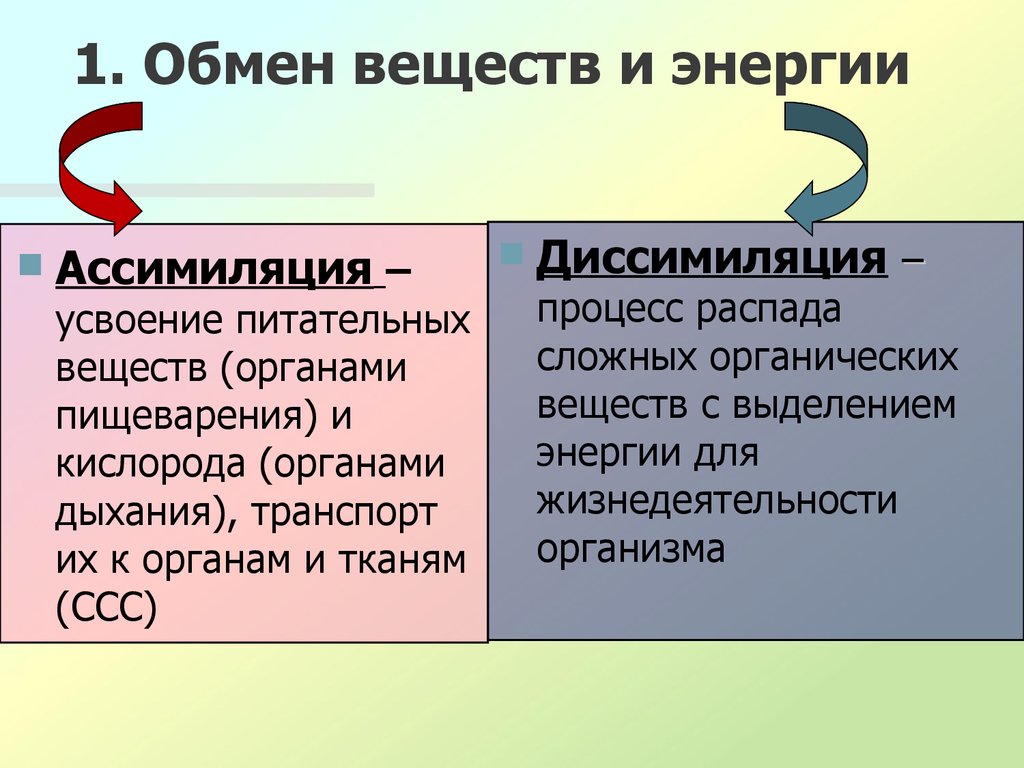
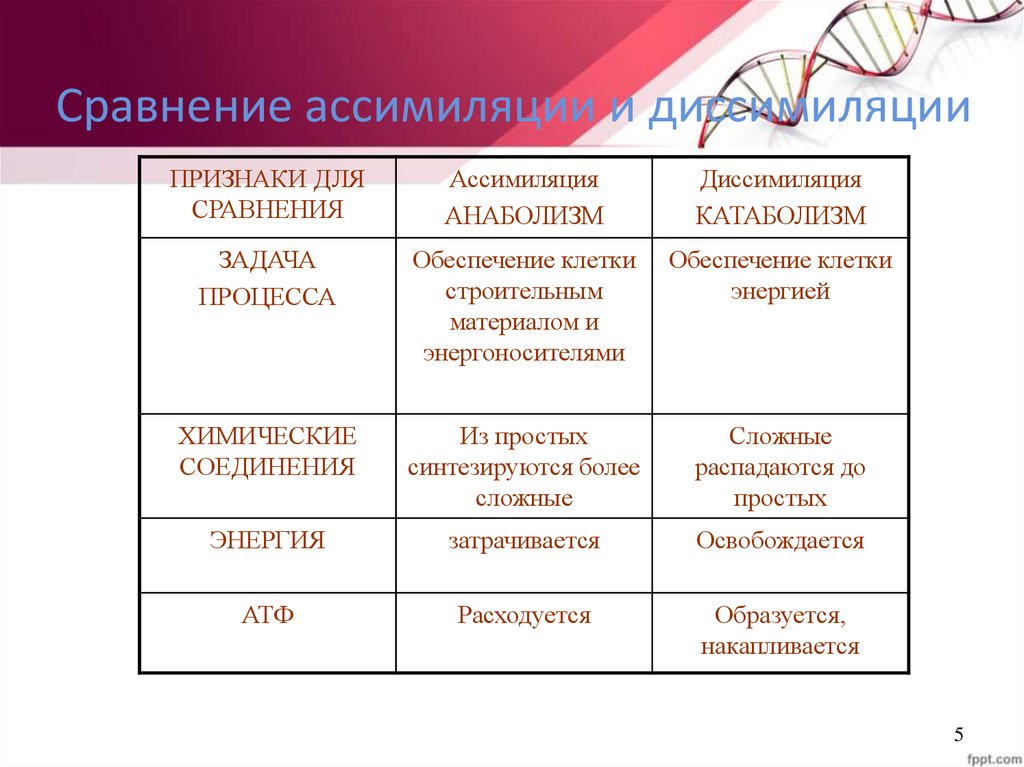
Он обеспечивает постоянство внутренней среды организма в изменяющихся условиях существования – гомеостаз. Обмен веществ слагается из двух взаимосвязанных и взаимопротивоположных процессов. Это процессы диссимиляции, в которых происходит расщепление органических веществ и выделенная энергия используется для синтеза молекул АТФ, и процессы ассимиляции, в которых энергия АТФ используется для синтеза собственных, необходимых организму соединений.
Процессы диссимиляции называют, также, катаболизмом и энергетическим обменом.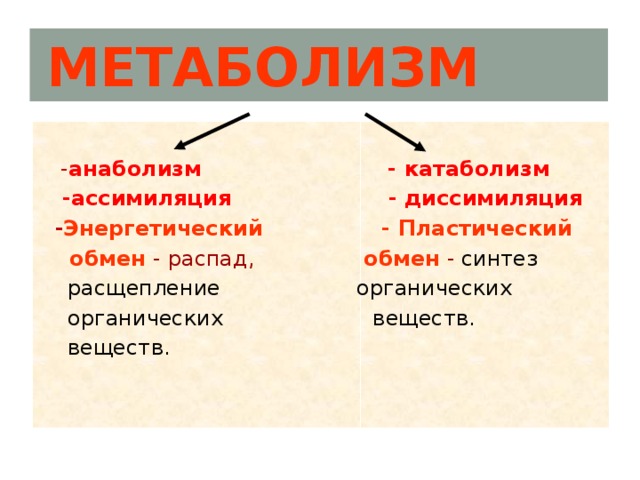 А процессы ассимиляции носят еще названия анаболизма и пластического обмена. Такое обилие синонимов одного и того же понятия возникло потому, что реакции обмена веществ изучали ученые различных специальностей:
А процессы ассимиляции носят еще названия анаболизма и пластического обмена. Такое обилие синонимов одного и того же понятия возникло потому, что реакции обмена веществ изучали ученые различных специальностей:
- биохимики,
- физиологи,
- цитологии,
- генетики,
- молекулярные биологи.
Но все названия и термины прижились и активно используются учеными.
Формы поступления энергии в живые организмы
Для всех живых организмов Земли Солнце является основным источником энергии. Именно благодаря ему организмы удовлетворяют свои энергетические потребности.
Организмы, которые могут синтезировать органические соединения из неорганических, называются автотрофами. Они разделяются на две группы. Одни способны использовать энергию солнечного света. Это – фотосинтетики или фототрофы. В основном это — зеленые растения, цианобактерии (сине-зеленые водоросли).
Другая группа автотрофов использует энергию, которая освобождается во время химических реакций. Такие организмы называются хемотрофами или хемосинтетиками.
Такие организмы называются хемотрофами или хемосинтетиками.
Грибы, большая часть животных и бактерий не могут сами синтезировать органические вещества. Такие организмы называются гетеротрофами. Для них источником энергии служат органические соединения, синтезированные автотрофами. Энергия используется живыми организмами для химических, механических, тепловых и электрических процессов.
Подготовительный этап энергетического обмена
Энергетический обмен принято условно разделять на три основных этапа. Первый этап назвали подготовительным. На этом этапе макромолекулы под воздействием ферментов расщепляются до мономеров. В ходе реакций происходит выделение довольно незначительного количества энергии, которое рассеивается в виде тепла.
Бескислородный этап энергетического обмена
Бескислородный (анаэробный) этап энергетического обмена происходит в клетках. Мономеры, которые образовались на предыдущем этапе (глюкоза, глицерин и т.п.), подвергаются дальнейшему многоступенчатому расщеплению без доступа кислорода.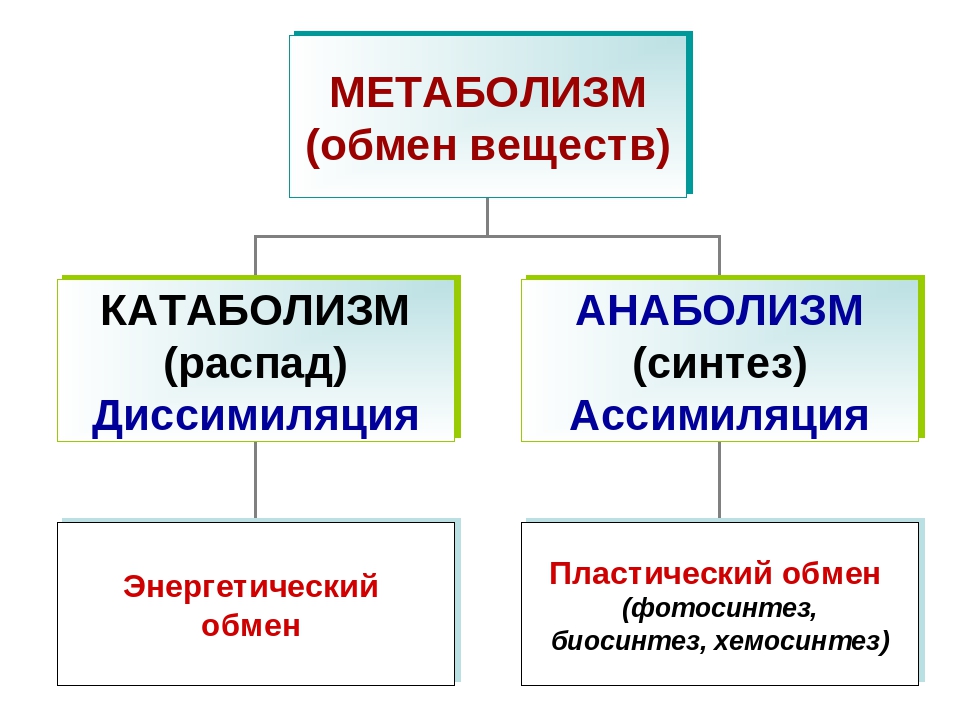 Главным на этом этапе является процесс расщепления молекулы глюкозы на молекулы пировиноградной или молочной кислоты с образованием двух молекул АТФ.
Главным на этом этапе является процесс расщепления молекулы глюкозы на молекулы пировиноградной или молочной кислоты с образованием двух молекул АТФ.
$C_6H_{12}O_6 + 2H_3PO_4 + 2АДФ → 2C_3H_6O_3 + 2АТФ + 2H_2O$
В ходе этой реакции (реакция гликолиза) выделяется около $200$ кДж энергии. Однако она не вся превращается в тепло. Часть ее используется для синтеза двух, богатых на энергию (макроэргических), фосфатных связей в молекулах АТФ. Глюкоза также расщепляется в ходе спиртового брожения.
$C_6H_{12}O_6 + 2H_3PO_4 + 2АДФ → 2C_2H_5OH + 2CO_2 + 2АТФ + 2H_2O$
Кроме спиртового существуют еще такие виды бескислородного брожения, как маслянокислое и молочнокислое.
Кислородный этап энергетического обмена
На этом этапе соединения, образованные на бескислородном этапе, окисляются до конечных продуктов реакции – углекислого газа и воды.
Английский биохимик Адольф Кребс в $1937$ году открыл последовательность превращений органических кислот в матриксе митохондрий.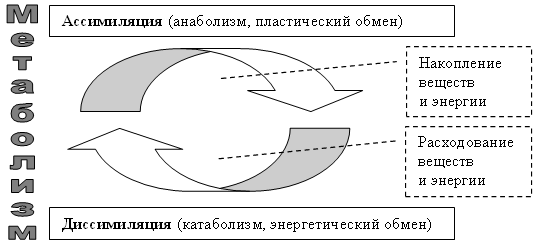 В его честь совокупность этих реакций назвали циклом Кребса.
В его честь совокупность этих реакций назвали циклом Кребса.
Замечание 1
Полное окисление молекул молочной или пировиноградной кислоты, образованных в ходе анаэробного процесса, до углекислого газа и воды сопровождается выделением $2800$ кДЖ энергии. Этого количества хватит на синтез $36$ молекул АТФ (в $18$ раз больше, чем на предыдущем этапе).
Суммарное уравнение кислородного этапа энергетического обмена выглядит так:
$2C_3H_6O_3 + 6O_2 + 36АДФ + 36H_3PO_4 → 6CO_2 + 42H_2O + 36АТФ$
Подводя общий итог, можно записать суммарное уравнение энергетического обмена:
$C_6H_{12}O_6 + 6O_2 + 38АДФ + 38H_3PO_4 → 6CO_2 + 44H_2O + 38АТФ$
На завершающей стадии происходит выведение продуктов метаболизма из организма.
Эффективность ассимиляции — Лаборатория физиологической экологии и биоэнергетики Лаборатория физиологической экологии и биоэнергетики
Ламантины в неволе питаются преимущественно салатом ромен, состав которого не сильно меняется в течение года.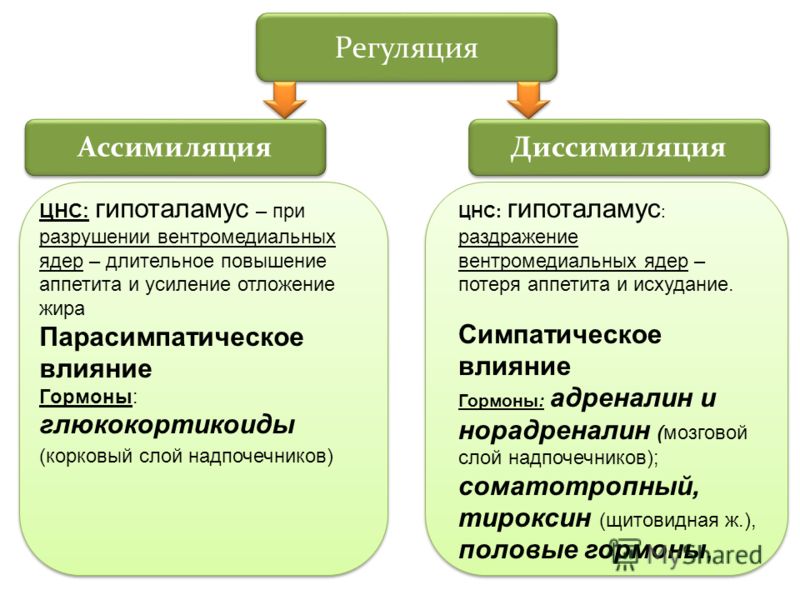 В дикой природе ламантины потребляют многочисленные пресноводные растения и морские травы, причем предпочтительными видами пресноводных растений являются южная наяда, гидрилла и параграс. Черепаховая трава, ламантиновая трава, мелководье и клеверная трава, вероятно, являются основными источниками пищи в морских системах. В меньшей степени ламантины могут также есть беспозвоночных, наземные травы, растущие достаточно близко к берегу, чтобы дотянуться до них, желуди, упавшие в воду, и мясо рыбы, пойманной в жаберные сети.
В дикой природе ламантины потребляют многочисленные пресноводные растения и морские травы, причем предпочтительными видами пресноводных растений являются южная наяда, гидрилла и параграс. Черепаховая трава, ламантиновая трава, мелководье и клеверная трава, вероятно, являются основными источниками пищи в морских системах. В меньшей степени ламантины могут также есть беспозвоночных, наземные травы, растущие достаточно близко к берегу, чтобы дотянуться до них, желуди, упавшие в воду, и мясо рыбы, пойманной в жаберные сети.
Количество питательных веществ, получаемых при употреблении в пищу растений, варьируется в зависимости от того, какая часть растения потребляется, а также зависит от возраста различных частей. На эффективность всасывания питательных веществ влияют морфология пищеварительного тракта, тип и качество пищи, сезонность, размер порции, время между приемами пищи и состояние питания животного. Иногда ламантины поедают только листья растений, а иногда выкапывают корневища и поедают все растение целиком. Когда ламантин концентрирует свое питание в одном месте в течение длительного времени, он может прогрызать туннели сквозь массу растительности.
Когда ламантин концентрирует свое питание в одном месте в течение длительного времени, он может прогрызать туннели сквозь массу растительности.
Мало что известно о том, сколько энергии ламантины усваивают из различных видов диких кормов, которые они потребляют. Встречающиеся в природе, по-видимому, неусвояемые минералы ранее использовались в качестве диетических маркеров у различных ящериц, птиц и млекопитающих. Во всех этих исследованиях было установлено, что марганец является хорошим маркером для использования в диетических исследованиях, поскольку он практически не всасывается через кишечник. Еще одним преимуществом использования марганца является то, что он потенциально полезен для определения эффективности ассимиляции у животных, живущих на свободном выгуле. Этот метод в основном включает сбор образцов пищи и фекалий и сравнение концентрации марганца в каждом из них.
Использование встречающегося в природе марганца в качестве инертного маркера позволяет нам измерить эффективность ассимиляции ламантинов в неволе и, теоретически, в дикой природе, а также оценить влияние различных условий окружающей среды на их пищевой статус. Цель настоящего проекта состояла в том, чтобы определить эффективность ассимиляции и, таким образом, количество получаемой энергии для ламантинов, потребляющих диету из морских водорослей.
Цель настоящего проекта состояла в том, чтобы определить эффективность ассимиляции и, таким образом, количество получаемой энергии для ламантинов, потребляющих диету из морских водорослей.
Долгое время предполагалось, что экология питания мегатравоядных в значительной степени зависит от их огромных размеров тела, и всегда считалось, что более крупные животные могут переносить диеты более низкого качества из-за более высокой эффективности пищеварения, которая является результатом длительного времени удержания ( из-за более длинного кишечника) и меньшей относительной потребности в энергии. О большинстве мегатравоядных имеется ограниченная информация, но, по-видимому, слоны не используют высокий пищеварительный потенциал, который теоретически обеспечивается их огромными размерами тела. У них, как правило, более быстрое время прохождения и более низкая эффективность пищеварения, чем можно было бы предсказать, основываясь исключительно на размере тела. По крайней мере, некоторые из этих признаков, вероятно, связаны с удивительно коротким (для их размера тела) пищеварительным трактом.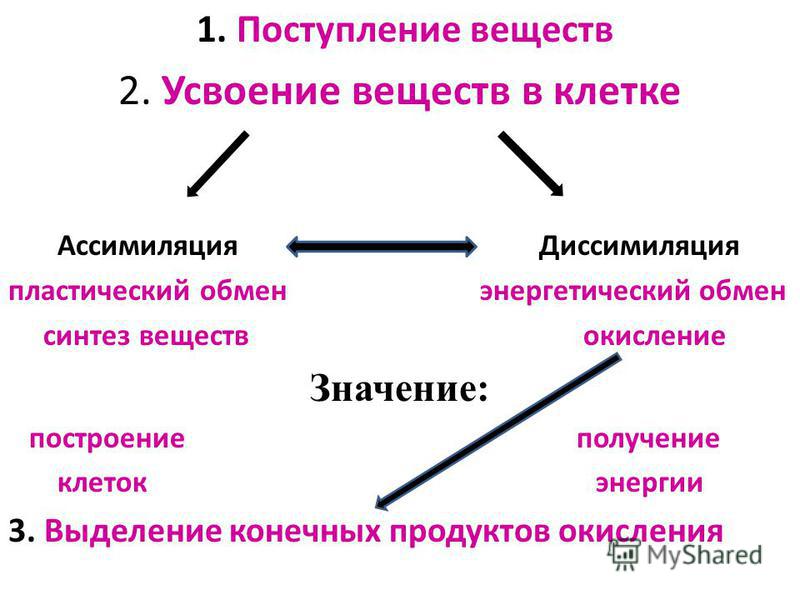 Измеренная эффективность варьируется от 22% для африканских слонов на диете в зоопарке до 73% для азиатских слонов, питающихся пальмовыми листьями. Большинство исследований указывают на эффективность рациона сена в пределах 33-53%. Данные о диких слонах-самцах, питающихся преимущественно травой в Национальном парке Крюгера, показали эффективность 290,7% – 44,7%.
Измеренная эффективность варьируется от 22% для африканских слонов на диете в зоопарке до 73% для азиатских слонов, питающихся пальмовыми листьями. Большинство исследований указывают на эффективность рациона сена в пределах 33-53%. Данные о диких слонах-самцах, питающихся преимущественно травой в Национальном парке Крюгера, показали эффективность 290,7% – 44,7%.
Эффективность ассимиляции (AE%) как содержащихся в неволе, так и находящихся на свободном выгуле западно-индийских ламантинов ( Trichechus manatus latirostris ) оценивали с использованием неассимилируемых косвенных маркеров, марганца и лигнина. AE%, измеренный с использованием марганца, существенно не отличался от AE%, оцененного с использованием лигнина, что подтверждает эффективность подхода с использованием марганца. Ламантины имеют AE%, сравнимый со слонами и лошадьми, но имеют пропорционально больший желудочно-кишечный тракт и значительно более длительное время транзита (7 дней). Образцы желудочно-кишечного тракта, полученные от недавно умерших животных, показали, что концентрации марганца были самыми низкими в желудке, оставались низкими в двенадцатиперстной кишке и тонкой кишке, но увеличивались в слепой кишке, толстой кишке и прямой кишке, что соответствовало пищеварению в толстой кишке. Дикие ламантины, потребляющие морскую растительность, имели значительно более низкий AE% [46,9].± 5,0% (n = 8)], чем те, которые потребляли пресноводную растительность [77,8 ± 6,9% (n = 7)], что, в свою очередь, было значительно ниже, чем содержащиеся в неволе ламантины, потребляющие салат [84,0 ± 4,4%, n = 37]. Дикие ламантины, питающиеся морскими водорослями, имели значительно более высокий процент AE%, чем животные, длительное время содержащиеся в неволе, потребляющие морские водоросли в течение коротких периодов времени (46,5 ± 4,4% против 41,1 ± 2,6% соответственно). Следствием низкой эффективности ассимиляции является то, что ламантинам, потребляющим морские травы, требуется большая стоячая биомасса для удовлетворения своих потребностей, чем это потребовалось бы, если бы они питались пресноводной растительностью. Учитывая большое количество морских водорослей, необходимых для поддержания жизни ламантинов, особенно в холодную погоду, крайне важно обеспечить сохранение и защиту среды обитания, прежде чем рассматривать вопрос о включении ламантинов в список исчезающих видов или исключении их из него.
Дикие ламантины, потребляющие морскую растительность, имели значительно более низкий AE% [46,9].± 5,0% (n = 8)], чем те, которые потребляли пресноводную растительность [77,8 ± 6,9% (n = 7)], что, в свою очередь, было значительно ниже, чем содержащиеся в неволе ламантины, потребляющие салат [84,0 ± 4,4%, n = 37]. Дикие ламантины, питающиеся морскими водорослями, имели значительно более высокий процент AE%, чем животные, длительное время содержащиеся в неволе, потребляющие морские водоросли в течение коротких периодов времени (46,5 ± 4,4% против 41,1 ± 2,6% соответственно). Следствием низкой эффективности ассимиляции является то, что ламантинам, потребляющим морские травы, требуется большая стоячая биомасса для удовлетворения своих потребностей, чем это потребовалось бы, если бы они питались пресноводной растительностью. Учитывая большое количество морских водорослей, необходимых для поддержания жизни ламантинов, особенно в холодную погоду, крайне важно обеспечить сохранение и защиту среды обитания, прежде чем рассматривать вопрос о включении ламантинов в список исчезающих видов или исключении их из него.
Биологическое разнообразие ассимиляции углерода среди изолятов дрожжей Dekkera bruxellensis из винных и топливно-этанольных промышленных процессов
. 2019 1 мая; 19 (3): foz022.
doi: 10.1093/femsyr/foz022.
Жаклин Мария да Силва 1 2 , Жилберто Энрике Телеш Гомеш да Силва 1 , Дениз Кастро Паренте 1 2
, Фернанда Кристина Безерра Лейте 3 , Каролина Сантос Сильва 4 , Патрисия Валенте 5 , Анжелика Мария Ганга 6 , Диогу Ардайон Симойнс 2 , Маркос Антонио де Мораис младший 1Принадлежности
- 1 Факультет генетики, Федеральный университет Пернамбуку, Ресифи, Бразилия.

- 2 Кафедра биохимии, Федеральный университет Пернамбуку, Ресифи, Бразилия.
- 4 Факультет химического машиностроения, Федеральный университет Пернамбуку, Ресифи, Бразилия.
- 5 Кафедра микробиологии, иммунологии и паразитологии, Федеральный университет Риу-Гранди-ду-Сул, Порту-Алегри, Бразилия.
- 6 Факультет пищевой инженерии, Университет Сантьяго, Сантьяго, Чили.

- PMID: 30848782
- DOI:
10. 1093/фемсир/foz022
 1093/фемсир/foz022
1093/фемсир/foz022Жакелин Мария да Силва и др. FEMS Yeast Res. .
. 2019 1 мая; 19 (3): foz022.
doi: 10.1093/femsyr/foz022.
Авторы
Жаклин Мария да Силва 1 2 , Жилберто Энрике Телеш Гомеш да Силва 1 , Дениз Кастро Паренте 1 2 , Фернанда Кристина Безерра Лейте 3 , Каролина Сантос Сильва 4 , Патрисия Валенте 5
Принадлежности
- 1 Факультет генетики, Федеральный университет Пернамбуку, Ресифи, Бразилия.
- 2 Кафедра биохимии, Федеральный университет Пернамбуку, Ресифи, Бразилия.
- 3 Факультет биологии Федерального сельского университета Пернамбуку, Ресифи, Бразилия.
- 4 Факультет химического машиностроения, Федеральный университет Пернамбуку, Ресифи, Бразилия.
- 5 Кафедра микробиологии, иммунологии и паразитологии, Федеральный университет Риу-Гранди-ду-Сул, Порту-Алегри, Бразилия.
- 6 Факультет пищевой инженерии, Университет Сантьяго, Сантьяго, Чили.
- PMID: 30848782
- DOI:
10. 1093/фемсир/foz022
 1093/фемсир/foz022
1093/фемсир/foz022Абстрактный
Dekkera bruxellensis считается дрожжами для порчи в виноделии, пивоварении и производстве топливного этанола. Однако в литературе появляется все больше свидетельств его биотехнологического потенциала. В данной работе мы рассмотрели 29Изоляты D. bruxellensis из трех стран и двух разных промышленных источников (виноделие и производство топливного этанола) для метаболизма промышленно значимых сахаров. Изоляты характеризовались определением их максимальных удельных скоростей роста и тестированием их способности к росту в присутствии 2-дезокси-d-глюкозы и антимицина А. Среди изолятов наблюдалось большое разнообразие, при этом изоляты топливного этанола демонстрировали в целом более высокие удельные темпы роста, чем у винных изолятов. Наблюдались предпочтения в отношении галактозы (три винных изолята) и целлобиозы или лактозы (некоторые изоляты топливного этанола).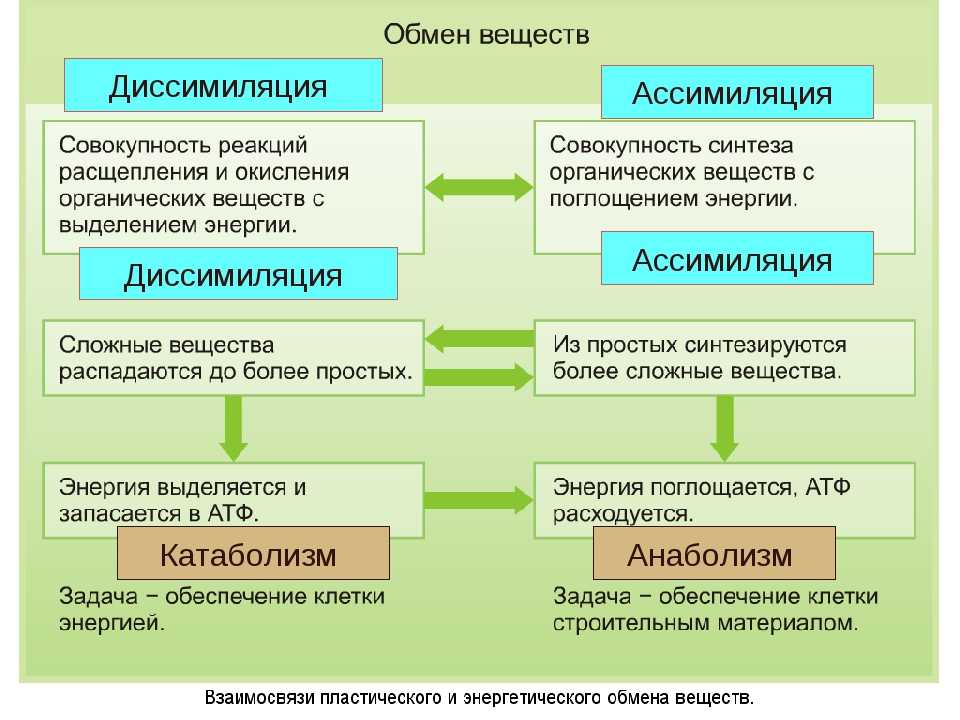
Ключевые слова: эффект Крэбтри; анаэробное культивирование; ассимиляция углерода; дисахариды; подавление катаболита глюкозы; избирательная адаптация.
© ФЭМС 2019.
Похожие статьи
Биотехнологический потенциал дрожжей Dekkera bruxellensis.
де Баррос Пита В., Телес Г.Х., Пенья-Морено И.С., да Силва Х.М., Рибейро К.С., де Мораис Жуниор М.А. де Баррос Пита В. и др. World J Microbiol Biotechnol. 201924 июня; 35(7):103. doi: 10.1007/s11274-019-2678-x. World J Microbiol Biotechnol. 2019. PMID: 31236799 Обзор.
Нитрат повышает анаэробную выработку этанола ацетат-зависимым образом у дрожжей Dekkera bruxellensis.
Пенья-Морено И.С., Кастро Паренте Д., да Силва Х.М., Андраде Мендонса А., Рохас Л.А.В., де Мораис Жуниор М.А., де Баррос Пита В. Пенья-Морено И.С. и соавт. J Ind Microbiol Biotechnol. 2019Фев; 46 (2): 209-220. doi: 10.1007/s10295-018-2118-1. Epub 2018 11 декабря. J Ind Microbiol Biotechnol. 2019. PMID: 30539327
Ферментация лигноцеллюлозного гидролизата альтернативными промышленными этанольными дрожжами Dekkera bruxellensis.
Блумквист Дж., Саут Э., Тиукова И., Момени М.Х., Ханссон Х., Стольберг Дж., Хорн С.Дж., Шнюрер Дж., Пассот В. Блумквист Дж. и соавт. Lett Appl Microbiol. 2011 июль; 53 (1): 73-8. doi: 10.1111/j.1472-765X.2011.03067.x. Epub 2011 31 мая. Lett Appl Microbiol. 2011. PMID: 21535044
Dekkera bruxellensis — дрожжи с биотехнологическим потенциалом и моделью эволюции, физиологии и конкурентоспособности дрожжей.
Блумквист Дж., Пассот В. Блумквист Дж. и соавт. FEMS Yeast Res. 2015 Июн;15(4):fov021. doi: 10.1093/femsyr/fov021. Epub 2015 7 мая. FEMS Yeast Res. 2015. PMID: 25956542 Обзор.
Взаимодействие Saccharomyces cerevisiae-Lactobacillus fermentum-Dekkera bruxellensis и исходного сырья при ферментации топливного этанола.
Bassi APG, Meneguello L, Paraluppi AL, Sanches BCP, Ceccato-Antonini SR. Басси АПГ и др. Антони Ван Левенгук. 2018 сен; 111 (9): 1661-1672. doi: 10.1007/s10482-018-1056-2. Epub 2018 27 февраля. Антони Ван Левенгук. 2018. PMID: 29488182

Посмотреть все похожие статьи
Цитируется
Сравнительный протеомный анализ раскрывает метаболические аспекты и биотехнологический потенциал ассимиляции нитратов у дрожжей Dekkera bruxellensis.
Пенья-Морено IC, Паренте Д.С., да Силва К.М., Пенья Э.П.Н., Сильва Ф.К., Кальса Джуниор Т., де Баррос Пита В., де Морайс М.А. младший. Пенья-Морено И.С. и соавт. Приложение Microbiol Biotechnol. 2021 фев;105(4):1585-1600. doi: 10.1007/s00253-021-11117-0. Epub 2021 4 фев. Приложение Microbiol Biotechnol. 2021. PMID: 33538877
Биотехнологический потенциал дрожжей Dekkera bruxellensis.