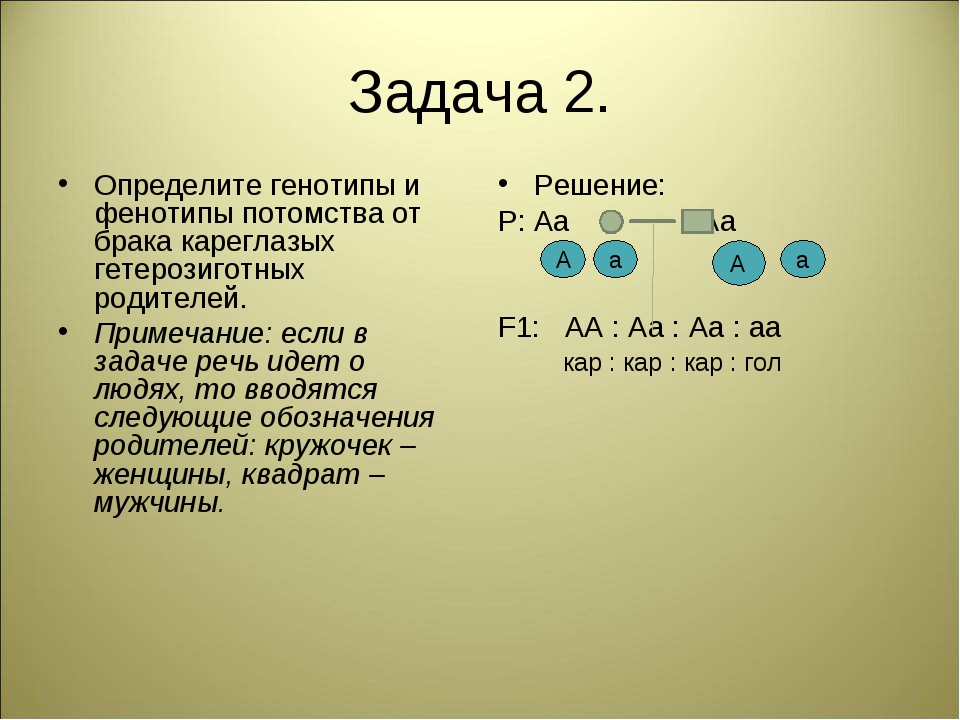
Генотип и фенотип, их изменчивость
Генотип — это совокупность всех генов организма, являющихся его наследственной основой.
Фенотип — совокупность всех признаков и свойств организма, которые выявляются в процессе индивидуального развития в данных условиях и являются результатом взаимодействия генотипа с комплексом факторов внутренней и внешней среды.
Каждый биологический вид имеет свойственный только ему фенотип. Он формируется в соответствии с наследственной информацией, заложенной в генах. Однако в зависимости от изменений внешней среды состояние признаков варьирует от организма к организму, в результате чего возникают индивидуальные различия — изменчивость.
На основе изменчивости организмов появляется генетическое разнообразие форм. Различают изменчивость модификационную, или фенотипическую, и генетическую, или мутационную.
Модификационная изменчивость не вызывает изменений генотипа, она связана с реакцией данного, одного и того же генотипа на изменение внешней среды: в оптимальных условиях выявляется максимум возможностей, присущих данному генотипу. Модификационная изменчивость проявляется в количественных и качественных отклонениях от исходной нормы, которые не передаются по наследству, а носят лишь приспособительный характер, например, усиление пигментации кожи человека под действием ультрафиолетовых лучей или развития мышечной системы под действием физических упражнений и т. д.
Степень варьирования признака у организма, то есть пределы модификационной изменчивости называются нормой реакции. Таким образом, фенотип формируется в результате взаимодействия генотипа и факторов среды, Фенотипические признаки не передаются от родителей к потомкам, наследуется лишь норма реакции, то есть характер реагирования на изменение окружающих условий.
Генетическая изменчивость бывает комбинативной и мутационной.
Комбинативная изменчивость возникает в результате обмена гомологичными участками гомологичных хромосом в процессе мейоза, что приводит к образованию новых объединений генов в генотипе.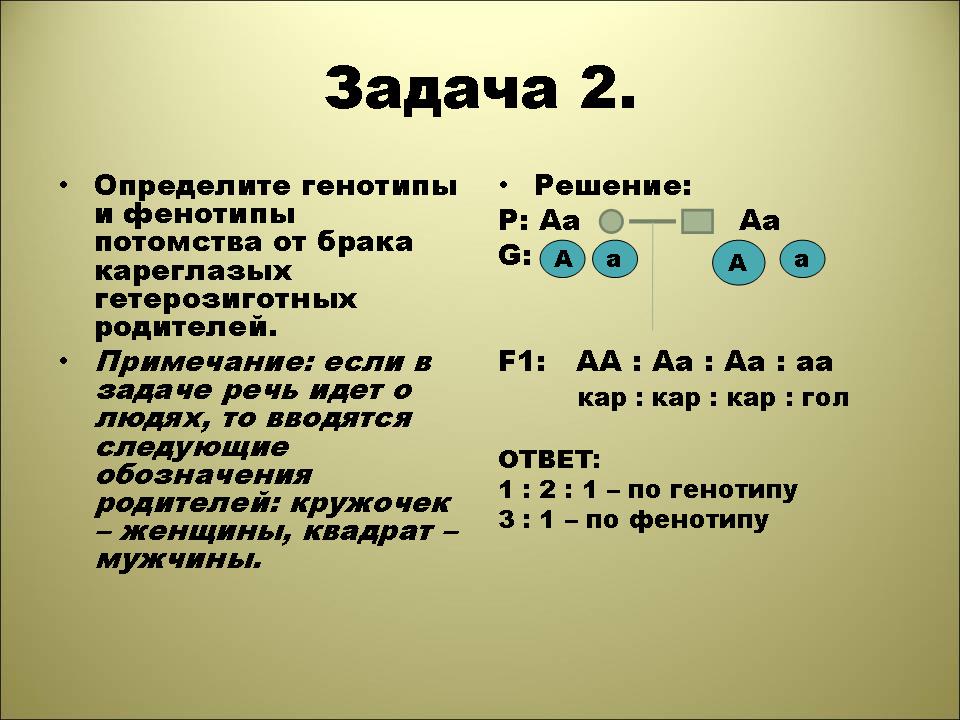
Мутационная изменчивость (мутации). Мутациями называют скачкообразные и устойчивые изменения единиц наследственности — генов, влекущие за собой изменения наследственных признаков. Они обязательно вызывают изменения генотипа, которые наследуются потомством и не связаны со скрещиванием и рекомбинацией генов.
Существуют хромосомные и генные мутации. Хромосомные мутации связаны с изменением структуры хромосом. Это может быть изменение числа хромосом кратное или не кратное гаплоидному набору (у растений — полиплоидия, у человека — гетероплоидия). Примером гетероплоидии у человека может быть синдром Дауна (одна лишняя хромосома и в кариотипе 47 хромосом), синдром Шерешевского — Тернера (отсутствует одна Х-хромосома, 45). Такие отклонения в кариотипе человека сопровождаются расстройством здоровья, нарушение психики и телосложения, снижением жизнеспособности и др.
Генные мутации — затрагивают структуру самого гена и влекут за собой изменение свойств организма (гемофилия, дальтонизм, альбинизм и др.). Генные мутации возникают как в соматических, так и в половых клетках.
Мутации, возникающие в половых клетках, передаются по наследству. Их называют генеративными мутациями. Изменения в соматических клетках вызывают соматические мутации, распространяющиеся на ту часть тела, которая развивается из изменившейся клетки. Для видов, размножающихся половым путем, они не имеют существенного значения, для вегетативного размножения растений они важны.
Генотип и фенотип
Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу
biorepet-ufa.ru.Вот такая получается «петрушка», если не сказать хуже. Очередной раз сталкиваюсь с тем, что основополагающие понятия генетики в учебниках преподносятся так, что разобраться в них бывает трудно.
Эту статью меня так и подмывало назвать сначала «Фенотип и генотип». Понятно, что фенотип вторичен от генотипа. Но если сам термин «генотип» учащиеся чаще всего могут истолковать правильно, то относительно понятия «фенотипа», как выясняется, нет четкого представления.
Да откуда же ему быть «четкому», если определения фенотипа в учебной литературе носят такой расплывчатый характер.
«Фенотип — совокупность всех внешних признаков организма, определяемых генотипом и условиями окружающей среды». Или «Фенотип — совокупность всех внешних и внутренних признаков и свойств организма, зависящих от генотипа и условий внешней среды».
А если действительно и «внешних», и «внутренних», а это на самом деле так, то в чем тогда отличие фенотипа от генотипа?
Всё же придется начинать не с «хвоста», а с «головы». Уверен, пройдет пара минут и вы, несколько уточнив для себя, что такое «генотип организма», сможете получить более четкое представление и о «фенотипе».
Часто термины «признак» и «ген» мы используем как синонимы
Говорят, «генотип — совокупность всех признаков организма». И вот тут то важно понять самое главное — именно к определению генотипа такое определение вносит дополнительную путаницу. Да, действительно, информация о любом признаке закодирована в каком-либо гене (или совокупности генов) организма.
Но всех генов очень много, весь генотип организма огромен, а в течение жизни данной особи или отдельной клетки реализуется (то есть служит образованию каких-либо определенных признаков) лишь незначительная часть генотипа.
Поэтому правильным будет запомнить, что
«генотип — совокупность всех генов организма». А уж какие из этих генов реализуются в течение жизни организма в его фенотипе , то есть послужат образованию каких-либо признаков — это зависит как от взаимодействия множества этих генов, так и от конкретных условий окружающей среды.
Таким образом, если правильно понимать, что собой представляет генотип, то не остается и лазейки для путаницы в терминах, обозначающих, что такое «генотип», а что такое «фенотип».
Понятно, что «фенотип — это совокупность всех реализовавшихся в течение жизни организма генов, послуживших образованию конкретных признаков данного организма в определенных условиях среды».
Поэтому на протяжении жизни организма, под действием меняющихся условий среды, фенотип может изменяться, хотя он и базируется на том же самом неизменном генотипе. А в каких границах может меняться фенотип?
Эти границы для фенотипа четко очерчены генотипом и носят название «нормы реакции». В фенотипе ведь не может проявиться ничего того, чего бы не было уже «записано» ранее в генотипе.
Чтобы лучше понять, что вкладывается в понятие «нормы реакции», разберем на конкретных примерах возможного проявления «широкой» или «узкой» нормы реакции.
Вес (масса) коровы и удойность коровы, какой признак имеет более широкую, а какой более узкую норму реакции?
Понятно, что вес взрослой коровы определенной породы как ее хорошо ни корми не может превысить, к примеру, 900 кг, а при плохом содержании — не может быть меньше 600 кг.
А удойность? При оптимальном содержании и кормлении удойность может меняться от каких-то максимально возможных для данной породы величин, она может упасть до 0, при неблагоприятных условиях содержания. Значит масса коровы имеет довольно узкую норму реакции, а удойность — очень широкую.
Пример с картофелем. Любому очевидно, что «вершки» имеют довольно узкую норму реакции, а масса клубней — очень широкую.
Думаю, теперь всё «устаканилось». Генотип — множество всех генов организма, это весь его потенциал на что он может быть способен в жизни. А фенотип — лишь проявление небольшой части этого потенциала, реализация лишь части генов организма в ряд конкретных признаков в течение его жизни.
Наглядным примером реализации в течение жизни организма части его генотипа в фенотип, являются однояйцевые близнецы. Имея абсолютно одинаковый генотип, в первые годы жизни они почти неотличимы друг от друга фенотипически. Но взрослея, имея сначала незначительные отличия в поведении, в каких-то привязанностях, отдавая предпочтение тому или иному роду деятельности, эти близнецы становятся довольно отличимыми и фенотипически: по выражению лица, строению тела.
В конце этой заметки, я бы хотел вот на что ещё обратить ваше внимание. Слово генотип для изучающих основы генетики имеет как бы два смысла. Выше мы разобрали значение «генотипа» в широком его понимании.
Но для уяснения законов генетики, при решении генетических задач, под словом генотип подразумевают лишь сочетание каких-то конкретных отдельных аллелей одной (моногибридное скрещивание) или двух (дигибридное скрещивание) пар определенных генов, контролирующих проявление конкретного одного или двух признаков.
То есть, и фенотип то у нас при этом какой-то усеченный, говорим «фенотип организма», а сами изучили механизм наследования лишь одного, двух его признаков. В широком же смысле термин «фенотип» относится к любым морфологическим, биохимическим, физиологическим и поведенческим характеристикам организмов.
P.S. В связи с характеристиками понятий «генотип» и «фенотип», уместным было бы здесь разобрать вопрос о наследственной и ненаследственной формах изменчивости организмов. Ну да ладно, об этом как раз и поговорим в следующей статье.
***************************************************************
У кого есть вопросы по статье к репетитору биологии по Скайпу, замечания, пожелания — прошу в писать комментарии.
Генотип — все статьи и новости
Генотип — совокупность генов организма и их отношений между собой, в том числе аллели и сцепление генов в хромосоме. В самом узком смысле это понятие может подразумевать все аллели гена или группы генов, контролирующих какой-либо признак организма, а в более широком — совокупность всех наследственных признаков организма, записанных в его ДНК (включая некодирующие последовательности), которые определяют его фенотип — внешний вид и внутреннее строение.
Сам термин «генотип» вместе с терминами «ген» и «фенотип» ввел немецкий генетик Вильгельм Людвиг Иогансен в 1909 году в своей работе «Элементы точного учения наследственности». Генотип не следует путать с геномом: в первом случае речь идет о наследственной информации конкретного организма, во втором — о наборе признаков, свойственном целому виду.
На ранних этапах изучения генотипа считалось, что все гены в нем действуют отдельно и каждый из них соответствует одному белку, выполняющему определенную функцию в организме и проявляющемуся как конкретный признак, который может быть качественным или количественным (например, рост, умение E.coli расщеплять какое-то питательное вещество, цвет лепестков ночной красавицы или гладкость или морщинистость горошины в опытах Менделя). Еще в первой половине XX века этот принцип — «один ген — один фермент» — считался передовым. Благодаря ему у ученых появилась сама возможность заняться расшифровкой генетического кода. Однако в дальнейшем было доказано, что генотип — это единая система элементов, которые взаимодействуют на разных уровнях. Например, один ген может кодировать более одного белка, не вся ДНК состоит из кодирующих последовательностей (экзонов), не все некодирующие последовательности бесполезны. Тот же рост контролируется вовсе не одним геном. Кроме того, гены влияют друг на друга, в результате чего несколько генотипов могут давать один и тот же фенотип или же действие нескольких аллелей или разных генов суммируется и дает один признак. Среди этих механизмов: неполное доминирование, как у ночной красавицы, когда при скрещивании пурпурного и белого цветка мы получаем «среднее арифметическое» — розовые цветки у потомства; эпистаз, когда один ген подавляет другой; кодоминирование, определяющее наследование агглютининов и агглютиногенов, на основании которых впервые было выделено четыре группы крови человека.
Также у особей с одинаковым генотипом в ходе индивидуального развития организма могут проявиться разные фенотипы, так как некоторые гены проявляются только в определенных условиях. Кроме того, если у двух организмов одинаковый фенотип, это не значит, что у них идентичные генотипы (при полном доминировании мы получаем доминантный фенотип и у гетерозигот, и у доминантных гомозигот). При этом фенотип может меняться на протяжении жизни организма, а генотип в целом остается неизменным (не считая ошибок при копировании ДНК и других мутаций).
Кроме того, если у двух организмов одинаковый фенотип, это не значит, что у них идентичные генотипы (при полном доминировании мы получаем доминантный фенотип и у гетерозигот, и у доминантных гомозигот). При этом фенотип может меняться на протяжении жизни организма, а генотип в целом остается неизменным (не считая ошибок при копировании ДНК и других мутаций).
Изображение: OpenClipart-Vectors/Pixabay
Генотипы и фенотипы дигетерозигот. Задачи 100
Определение генотипов и фенотипов при скрещивании дигетерозигот
Задача 100.
У томатов цвет красной окраски плода и округлой формы — доминантные, а жёлтый и продолговатая форма плода — рецессивные.
1) Определить генотип и фенотип F1, если скрестили между собой гомозиготные растения, имеющие красный цвет и округлую форму плода.
2) Определить генотип F1, если скрещивамые гомозиготные растения имели жёлтый цвет и округлую форму плода.
3) Определить генотип и фенотип F1, если скрестили растения имеющие красный цвет и продолговатую форму плода.
Задачи на дигибридное скрещивание с применением третьего закона Менделя.
Дано:
А — ген красной окраски плодов томата;
а — ген жёлтой окраски плодов томата;
В — ген округлой формы плодов томата;
в — ген продолговатой формы плодов томата;
АаВв — дигетерозиготное растение томата;
А_вв — растение, имеющее красный цвет и продолговатую форму плода.
Решение:
1. Определение генотипов и фенотипов F1, если скрестили между собой растения, имеющие красный цвет и округлую форму плода
Схема скрещивания
P: ААВВ х ААВВ
G: АВ АВ
F1: ААВВ — 100%.
Наблюдается один тип генотипа. Расщепления по генотипу нет.
Фенотип:
ААВВ — красный цвет и округлая форма плода — 100%.
Наблюдается один тип фенотипа. Расщепления по фенотипу нет.
Ответ: в данном случае расщепления по генотипу и фенотипу нет, т. е, с вероятностью 100% образуются плоды ААВВ (красные округлые).
2. Определение генотипов F1, если скрещиваемые гомозиготные растения томата имели жёлтый цвет и округлую форму плода.
Схема скрещивания
P: ааВВ х ааВВ
G: аВ аВ
F1: ааВВ — 100%.
Наблюдается один тип генотипа. Расщепления по генотипу нет.
Фенотип:
ааВВ — желтый цвет и округлая форма плода — 100%.
Наблюдается один тип фенотипа. Расщепления по фенотипу нет.
Ответ: в данном случае расщепления по генотипу и фенотипу нет, т. е, с вероятностью 100% образуются плоды ааВВ (желтые округлые).
3. Определение генотипов F1, если скрещиваемые гомозиготные растения томата имели красный цвет и продолговатую форму плода.
Схема скрещивания
P: ААвв х ААвв
G: Ав Ав
F1: ААвв — 100%.
Наблюдается один тип генотипа. Расщепления по генотипу нет.
Фенотип:
ААвв — красный цвет и продолговатая форма плода — 100%.
Ответ: в данном случае расщепления по генотипу и фенотипу нет, т. е, с вероятностью 100% образуются плоды ААвв (красные продолговатые).
Выводы:
1. Во всех трех случаев скрещиваний расщеплений по генотипу и фенотипу нет.
Задача 101.
У человека сложные формы близорукости доминируют над нормальным зрением, карий цвет глаз – над голубым. Кареглазый близорукий мужчина, мать которого имела голубые глаза и нормальное зрение, женился на голубоглазой женщине с нормальным зрением. Какова вероятность в % рождения ребенка с признаками матери?
Решение:
Так как у кареглазого близорукого мужчины, мать имела голубые глаза и нормальное зрение, то мужчина является дигетерозиготой по обоим признакам. Голубоглазая женщина с нормальным зрением является дигомозиготой по обоим рецессивным признакам.
Голубоглазая женщина с нормальным зрением является дигомозиготой по обоим рецессивным признакам.
А — карий цвет глаз;
а — голубой цвет глаз;
В — близорукость;
в — нормальное зрение;
АаВв — генотип мужчины;
аавв — генотип женщины.
Схема скрещивания:
Р: аавв х АаВв
Г: ав АВ; Ав; аВ; ав.
F1: АаВв — 25%; Аавв — 25%; ааВв — 25%; аавв — 25%.
Фенотипы:
АаВв — карие глаза, близорукость — 25%;
Аавв — карие глаза, нормальное зрение — 25%;
ааВв — голубые глаза, близорукость — 25%;
аавв — голубые глаза, нормальное зрение — 25%.
Вывод:
Вероятность рождения ребенка с признаками матери составляет 25%.
Genetic Lingo | Protocol (Translated to Russian)
12.1: Генетический язык
Обзор
Организм диплоидный, если он наследует два варианта, или аллели, каждого гена, по одному от каждого родителя. Эти два аллеля представляют собой генотип данного гена. Термин генотип также используется для обозначения полного набора генов организма. Диплоидный организм с двумя одинаковыми аллелями имеет гомозиготный генотип, в то время как два разных аллеля указывают на гетерозиготный генотип. Наблюдаемые черты, вытекающие из генотипов, называются фенотипами, на которые также могут влиять факторы окружающей среды. Аллель доминирует, если только одна копия необходима для проявления связанного фенотипа и рецессивного, если для фенотипического выражения требуется две копии.
Аллели могут влиять на фенотипы
Диплоидные организмы, в том числе люди, большинство других животных, и многие растения, имеют дубликат набора хромосом в соматических (неполовых) клетках. Эти хромосомные дубликаты гомологичны, имеют сходную длину, положения центромера и расположение генов. Диплоидные организмы наследуют по две версии каждого гена, по одному от каждого родителя. Эти два варианта генов, или аллели, расположены в одном относительном локусе, или положении, на двух гомологичных хромосомах. Каждая хромосома содержит много генетических локусов, и часто существует несколько возможных аллелей данного гена.
Диплоидные организмы наследуют по две версии каждого гена, по одному от каждого родителя. Эти два варианта генов, или аллели, расположены в одном относительном локусе, или положении, на двух гомологичных хромосомах. Каждая хромосома содержит много генетических локусов, и часто существует несколько возможных аллелей данного гена.
Два аллеля, унаследованные диплоидным организмом, составляют его. генотип в локусе. Термин генотип также относится к общему набору генов организма. Различные генотипы могут приводить к различным фенотипам или наблюдаемым характеристикам (например, цвет глаз). Фенотипы возникают в результате генотипов, хотя несколько генотипов могут вызывать один и тот же фенотип. Фенотипы также часто находятся под влиянием факторов окружающей среды.
Генотип может состоять из двух одинаковых или различных аллелей
Для данного гена два унаследованных аллеля могут иметь идентичные или различные нуклеотидные последовательности. Организм с двумя идентичными аллелями имеет гомозиготный генотип (или гомозигот). Организм с двумя разными аллелями имеет гетерозиготный генотип (т.е. гетерозигот). Если два аллеля отличаются (гетерозиготный генотип), но только один влияет на фенотип, — доминирует аллель, влияющий на фенотип. Другой аллель, который не влияет на фенотип, является рецессивным.
Первоначально считалось, что цвет глаз определяется одним геном, и поэтому он часто используется для иллюстрации генетического доминирования (коричневые глаза) и рецессивности (голубые глаза). Тем не менее, ученые обнаружили по крайней мере восемь генов, которые регулируют цвет глаз. Хотя ген OCA2 отвечает за почти три четверти вариации в сине-коричневом цветовом спектре глаз, другие гены иногда изменяют или переопределяют эти эффекты. Тем не менее, цвет глаз может быть полезным примером для иллюстрации доминирующих и рецессивных аллелей.
В следующем примере мы используем упрощенный пример цвета глаз, чтобы проиллюстрировать связь между генотипом и фенотипом. Аллель для карих глаз обозначается B, а аллель для голубых глаз b. Гетерозигота будет иметь генотип Bb в результате получения доминирующей аллель от одного родителя и рецессивный аллель от другого. Этот человек будет иметь карие глаза, доминирующий фенотип. Доминирующий аллель(B)переопределяет или «скрывает» инструкции от рецессивного аллеля для голубых глаз. Таким образом, человек с голубыми глазами обязательно будет иметь генотип bb, в то время как человек с карими глазами может иметь либо генотип BB или Bb.
Аллель для карих глаз обозначается B, а аллель для голубых глаз b. Гетерозигота будет иметь генотип Bb в результате получения доминирующей аллель от одного родителя и рецессивный аллель от другого. Этот человек будет иметь карие глаза, доминирующий фенотип. Доминирующий аллель(B)переопределяет или «скрывает» инструкции от рецессивного аллеля для голубых глаз. Таким образом, человек с голубыми глазами обязательно будет иметь генотип bb, в то время как человек с карими глазами может иметь либо генотип BB или Bb.
Доминирующие и рецессивные модели наследования заболеваний связаны с риском заболевания
Хотя большинство признаков контролируются несколькими генами, а не одним причинным геном, эти принципы могут быть полезны для прогнозирования вероятности важных результатов. Например, болезнь Хантингтона (HD) является прогрессирующим, нейродегенеративным состоянием, которое вызывает неконтролируемые движения и когнитивные и эмоциональные нарушения. HD считается одним геном расстройства, поскольку она вызвана мутацией в одном гене (HTT), хотя другие гены могут изменить течение заболевания. HD вызвано доминирующей мутацией, а это означает, что один мутировавший аллель приводит к болезни. Если родитель имеет нормальный и мутировавший вариант гена, потомство имеет 50% шанс получить мутировавший аллель, и заболевание разовьётся. HD также обычно не проявляется до среднего возраста. Таким образом, хотя реальность неприятна, человек с пострадавшим родителем может пройти генетическое тестирование, чтобы определить, есть ли у них причинно-следственная аллель и, в свою очередь, информировать их планирования семьи.
Литература для дополнительного чтения
Fay, David, and Andy Spencer. Dominant Mutations. WormBook, 2006. [Source].
Генотип и фенотип
|
Любая черта организма, то есть любое его свойство или особенность, по которым он отличается от других экземпляров, называется признаком в генетике и селекции. Определение понятийКак же определить генотип и фенотип? Ниже приводится понятие о генотипе и фенотипе. Комплекс всех признаков экземпляра растения, включая внешние черты и особенности структуры, функционирования клеток и тканей, называется фенотипом. Данное обозначение можно употреблять и относительно одного из альтернативных (противоположных) признаков. Генотипом регулируется развитие всех наследственных признаков и свойств организма. Таким образом, генотип – это совокупность всех носителей наследственной информации данной клетки и организма в целом. При скрещивании экземпляров растений наблюдается расщепление как по генотипу, так и по фенотипу. При становлении генетических принципов о связи между геном и признаком считалось, что в соответствии с каждым признаком имеется специальный наследственный фактор. Но позднее было выяснено, что прямые однозначные связи гена с признаком обычно являются исключением, а не правилом. Ученые доказали, что имеется множество свойств и признаков растений, развитие которых контролируется двумя, тремя либо даже несколькими парами генов. Может быть и наоборот, когда одним геном определяется ряд признаков. Поэтому четкое соотношение генотипа и фенотипа установить сложно. Также воздействие гена может подвергаться изменениям в связи с соседством с другими генами и влиянием факторов внешней среды. Так, в онтогенезе работают не отдельные гены, а весь геном как цельная исторически сложившаяся система, характеризующаяся сложными связями и взаимоотношениями между генами и окружающей средой. Биологическое значение кроссинговера невозможно переоценить. Генетическая рекомбинация дает возможность производить новые, не существовавшие ранее комбинации генов. Это позволяет обеспечить увеличение выживаемости организмов в ходе эволюции. Похожие статьи: |
 Примером признака у растений может служить форма венчика (симметричный, асимметричный), окраска венчика цветка (розовый, белый), время созревания плодов у растений (скороспелые, позднеспелые). Помимо этого, признаком является способность или неспособность растения к продуцированию определенного вещества, к примеру, крахмала, масла, фитонцидов, а также стойкость к каким-либо болезням или восприимчивость к ним и т.п.
Примером признака у растений может служить форма венчика (симметричный, асимметричный), окраска венчика цветка (розовый, белый), время созревания плодов у растений (скороспелые, позднеспелые). Помимо этого, признаком является способность или неспособность растения к продуцированию определенного вещества, к примеру, крахмала, масла, фитонцидов, а также стойкость к каким-либо болезням или восприимчивость к ним и т.п. Кроме того, данная система непрерывно изменяется и совершенствуется с течением времени.
Кроме того, данная система непрерывно изменяется и совершенствуется с течением времени.47. Генотип и фенотип программных систем | by Michael Dubakov
Полгода назад, читая Лема, я набрел на генотипы и фенотипы. И конечно в голову полезли мысли, как это распространить на программные продукты.
Генотип и ФенотипЕсли очень кратко, то генотип закладывает какие-то свойства в систему, а внешняя среда накладывает свой отпечаток и формирует фенотип на основе этих свойств и контекста. То есть отдельная живая особь содержит больше информации, чем генотип, потому что к информации генотипа добавляется внешняя информация среды. Под влиянием внешней среды происходит экспрессия генов. К примеру, под воздействием антибактериального мыла и заботливой мамы у вас развивается склонность к аллергии. В генах это заложено, но вот Васе мама разрешала копаться в грязи, и у него нет аллергии. А вы ненавидите апрель.
Возьмем для конкретики любой продукт. Ну, например, Targetprocess. Он имеет генотип (заложенные в него функции), но каждый клиент настраивает систему под себя, так что каждый отдельный аккаунт Targetprocess — это фенотип. Мы сейчас можем смотреть на ситуацию с эволюционной точки зрения. Генотип Targetprocess порождает множество своих экземпляров, которые сталкиваются с внешней средой (клиентами). В этой внешней среде экземпляры либо выживают (компания пользуется продуктом и все хорошо), либо умирают (компания отказывается от продукта).
Внешняя среда (контекст использования) может оказывать существенное влияние на конкретный экземпляр системы. Фенотип накапливает в себе информацию: домен компании, данные, недовольные пользователи и их фидбек, настройки, расширения, интеграции, и тп. В будущем эта накопленная информация по идее может вызывать мутации в генотипе. Фактически, это и есть задача Product Owner —собирать данные о фенотипах и закладывать нужные мутации в генотип продукта. От того, что заложено в генотипе, зависит выживаемость каждого конкретного экземпляра системы, который попал в свой специфический внешний контекст.
В будущем эта накопленная информация по идее может вызывать мутации в генотипе. Фактически, это и есть задача Product Owner —собирать данные о фенотипах и закладывать нужные мутации в генотип продукта. От того, что заложено в генотипе, зависит выживаемость каждого конкретного экземпляра системы, который попал в свой специфический внешний контекст.
Дальше самое интересное. При однообразии контекстов генотип может быть довольно однозначный, потому что ясно к чему стремиться и что оптимизировать. Продукт используют похожим образом и Product Owner будет получать большое количество одинаковых сигналов от разных экземпляров, поэтому легко понять, что нужно изменить в системе. В предельном случае, если вы делаете систему под одного заказчика, генотип = фенотипу. Вы просто делаете все, что нужно этому заказчику.
Чуть более сложный случай, когда все ваши клиенты очень похожи. Например, это софт для ведения бухгалтерии ИП. Единый стандарт с минимальными изменениями контекста использования делает жизнь PO довольно простой. Вам достаточно иметь несколько клиентов и поддерживать с ними контакт, чтобы реализовать всю нужную функциональность.
Разнообразный контекст
При разнообразии контекстов все становится гораздо сложнее. Каждый экземпляр имеет уникальный фенотип. Клиенты просят довольно разные вещи, и вы не можете делать однозначные выводы на основе фидбека даже сотни клиентов. Жизнь PO становится невыносимой. Принимать правильные решения невероятно сложно, потому что фича, решающая проблему 10% клиентов, совершенно не нужна оставшимся 90%. Что самое хреновое, очень мало таких фич, которые нужны почти всем. Каждая фича нужна немногим. И вы попадаете в ад.
Фактически вы приходите к тому, что при разнообразии контекстов генотип должен быть вариативный. Вы должны обеспечить возможность экспрессии генов. Это значит, что отдельные экземпляры системы должны иметь возможность приспособиться к очень разным контекстам и выжить в непредсказуемой среде. В софте эта вариативность реализуется через обширность настроек или платформу с приложениями. Если вы решили делать софт на рынке с большим разнообразием контекстов — нужно делать систему гибкой. Другого пути просто нет. Сразу скажу, что обширность настроек — очень стремный путь. Рано или поздно продукт просто умрет из-за внутренней сложности.
Это значит, что отдельные экземпляры системы должны иметь возможность приспособиться к очень разным контекстам и выжить в непредсказуемой среде. В софте эта вариативность реализуется через обширность настроек или платформу с приложениями. Если вы решили делать софт на рынке с большим разнообразием контекстов — нужно делать систему гибкой. Другого пути просто нет. Сразу скажу, что обширность настроек — очень стремный путь. Рано или поздно продукт просто умрет из-за внутренней сложности.
Если взять Fibery, то мы пошли от предпосылки большого разнообразия контекстов. На самом деле за годы эксплуатации Targetprocess мы выяснили, что компании работают очень разнообразно. Их процессы часто уникальны и выдвигаемые требования хрен реализуешь в рамках одного целостного продукта. Поэтому, если мы хотим быть на рынке систем управления проектами, мы должны заложить в генотип Fibery значительную гибкость.
Что интересно, сам факт закладывания этой гибкости приведет к тому, что Fibery будут чаще использовать в самых непредсказуемых и разнообразных контекстах. Мы получаем очень веселый positive feedback loop:
Гибкая система → неожиданное использование → требование еще большей гибкости.
Пока сложно представить, к чему это все приведет. Но кажется впереди у нас много веселых бесед об эволюции, сложности и качестве 🌶.
Генотип против фенотипа: примеры и определения
Любой организм является побочным продуктом как его генетической структуры, так и окружающей среды. Чтобы понять это в деталях, мы должны сначала оценить некоторые основные генетические термины и концепции. Здесь мы даем определения терминов генотип и фенотип , обсуждаем их взаимосвязь и рассматриваем, почему и как мы могли бы их изучить.
В биологии ген — это часть ДНК, которая кодирует признак.Точное расположение нуклеотидов (каждый из которых состоит из фосфатной группы, сахара и основания) в гене может различаться в разных копиях одного и того же гена. Следовательно, у разных организмов ген может существовать в разных формах. Эти разные формы известны как аллели. Точное фиксированное положение на хромосоме, содержащее конкретный ген, называется локусом.
Следовательно, у разных организмов ген может существовать в разных формах. Эти разные формы известны как аллели. Точное фиксированное положение на хромосоме, содержащее конкретный ген, называется локусом.
Диплоидный организм наследует либо две копии одного и того же аллеля, либо одну копию двух разных аллелей от своих родителей. Если человек наследует два идентичных аллеля, его генотип считается гомозиготным по этому локусу.
Однако, если они обладают двумя разными аллелями, их генотип классифицируется как гетерозиготный для этого локуса. Аллели одного и того же гена могут быть аутосомно-доминантными или рецессивными. Аутосомно-доминантный аллель всегда будет предпочтительно выражаться по сравнению с рецессивным аллелем.
Последующая комбинация аллелей, которыми обладает человек для определенного гена, — это его генотип .
Рассмотрим классический пример — цвет глаз.
- Ген кодирует цвет глаз.
- В этом примере аллель либо коричневый, либо синий, причем один унаследован от матери, а другой — от отца.
- Коричневый аллель является доминантным (B), а синий аллель — рецессивным (b). Если ребенок наследует два разных аллеля (гетерозиготных), у него будут карие глаза. Чтобы у ребенка были голубые глаза, они должны быть гомозиготными по аллелю голубого глаза.
Рис. 1. Диаграмма наследования, в которой подробно показано, как индивид может унаследовать голубые или карие глаза в зависимости от аллелей, принадлежащих их родителям, причем аллель цвета карих глаз является доминантным, а аллель цвета голубых глаз — рецессивным.
Другие примеры генотипа включают:
- Цвет волос
- Рост
- Размер обуви
Сумма наблюдаемых характеристик организма — это их фенотип. Ключевое различие между фенотипом и генотипом состоит в том, что генотип наследуется от родителей организма, а фенотип — нет.
Хотя фенотип влияет на генотип, генотип не равен фенотипу.На фенотип влияет генотип и факторы, включая:
- Эпигенетические модификации
- Факторы окружающей среды и образа жизни
Рисунок 2: Фламинго от природы имеют белый цвет, их вызывают только пигменты организмов, которые они едят чтобы стать ярко-розовым.
Примеры фенотипов
Факторы окружающей среды, которые могут влиять на фенотип, включают питание, температуру, влажность и стресс.Фламинго — классический пример того, как окружающая среда влияет на фенотип. Несмотря на то, что они известны своим ярко-розовым цветом, их естественный цвет — белый — розовый цвет вызван пигментами организмов в их рационе.
Второй пример — цвет кожи человека. Наши гены контролируют количество и тип меланина, который мы производим, однако воздействие ультрафиолетового света в солнечном климате вызывает потемнение существующего меланина и способствует усилению меланогенеза и, следовательно, более темной коже.
Генотип против фенотипа: наблюдениеНаблюдать за фенотипом просто — мы смотрим на внешние особенности и характеристики организма и делаем выводы о них.Однако наблюдение за генотипом немного сложнее.
Генотипирование — это процесс, при котором различия в генотипе человека анализируются с помощью биологических анализов. Полученные данные затем можно сравнить либо с последовательностью второго человека, либо с базой данных последовательностей.
Ранее генотипирование позволяло получить только частичные последовательности. Теперь, благодаря крупным технологическим достижениям последних лет, мы получили самое современное секвенирование всего генома.
Рисунок 3: Рабочий процесс, изображающий различные этапы полногеномного секвенирования (WGS).
(WGS) позволяет получать целые последовательности. WGS — это эффективный процесс, который становится все более доступным и включает использование высокопроизводительных методов секвенирования, таких как секвенирование одной молекулы в реальном времени (SMRT), для идентификации исходной последовательности нуклеотидов, составляющих ДНК организма.
WGS — это не единственный способ анализа генома организма — доступны различные методы.
Почему важно изучать генотип по сравнению с фенотипом?Понимание взаимосвязи между генотипом и фенотипом может быть чрезвычайно полезным в различных областях исследований.
Особенно интересная область — фармакогеномика. Генетические вариации могут возникать в ферментах печени, необходимых для метаболизма лекарств, таких как CYP450. Следовательно, фенотип человека, то есть его способность метаболизировать конкретное лекарство, может варьироваться в зависимости от того, какой формой гена, кодирующего фермент, он обладает. Для фармацевтических компаний и врачей эти знания являются ключевыми для определения рекомендуемых дозировок лекарств для разных групп населения.
Тандемное использование методов генотипирования и фенотипирования оказывается лучше, чем использование только генотипических тестов.В сравнительном клиническом исследовании фармакогеномики подход мультиплексирования выявил большие различия в способности метаболизма лекарств, чем это было предсказано только генотипированием. Это имеет важное значение для персонализированной медицины и подчеркивает необходимость проявлять осторожность, полагаясь исключительно на генотипирование.
Как мы можем изучить взаимосвязь между генотипом и фенотипом?Используя модели животных, например мышей, ученые могут генетически модифицировать организм так, чтобы он больше не экспрессировал определенный ген — известные как «нокаутные мыши».Сравнивая фенотип этого животного с фенотипом дикого типа (то есть фенотипом, который существует, когда ген не был удален), мы можем изучить роль определенных генов в обеспечении определенных фенотипов.
Инициатива в области информатики генома мышей (MGI) собрала базу данных тысяч фенотипов, которые могут быть созданы и изучены, а также генов, которые должны быть выбиты для получения каждого конкретного фенотипа.
| | Генотип | набор генов в нашей ДНК, которые отвечают за определенный признак | Наблюдаемые характеристики и признаки организма | |
| Характеризуется | Методы генотипирования, такие как WGS | Наблюдение за внешними характеристиками организма | ||
| Зависит от | Последовательности генов, которыми обладает организм | Генотип, ПЛЮС эпигенетика и факторы окружающей среды | ||
| Унаследовано? | Да | Нет | ||
| Пример | Гены, кодирующие цвет глаз | Человек с карими глазами |
примеров генотипа и фенотипа: выявленные различия
Генотип и фенотип — два фундаментальных термина в науке генетики. Эти два термина часто используются одновременно для описания одного и того же организма, но между генотипом и фенотипом есть разница:
Эти два термина часто используются одновременно для описания одного и того же организма, но между генотипом и фенотипом есть разница:
- Генотип организма — это набор генов в его ДНК, ответственных за определенный признак.
- Фенотип организма — это физическое выражение этих генов.
Например, две практически идентичные мыши могут иметь разные генотипы. Но если у них явно разные черты — скажем, у одного белый мех, а у другого черный мех, — тогда у них разные фенотипы.
Объяснение генотипа
Как генотип влияет на фенотип? Простой. Генотип — это совокупность генов, отвечающих за различные генетические особенности данного организма. Генотип относится конкретно к генам, а не к признакам; то есть необработанная информация в ДНК организма.
Генотип определяется составом аллелей, пар генов, отвечающих за определенные признаки. Аллель может состоять из двух доминантных генов, доминантного и рецессивного гена или двух рецессивных генов.Комбинация двух и того, какой из них является доминантным, определяет, какой признак будет выражать аллель.
Генотип просто означает, какие аллели содержатся в ДНК конкретного организма. Это не может быть определено простым наблюдением; это требует биологического тестирования. Генотип передается по наследству от родителей организма и выражает всю генетическую информацию о нем.
Примеры генотипа
Трудно привести конкретные примеры генотипа. Этот термин просто означает «гены, которые есть у конкретного организма.«Любой пример генотипа будет просто диаграммой хромосом конкретного живого существа или молекул ДНК, ответственных за различные генетические признаки. Однако наличие определенных генов действительно дает заметные результаты.
Например, если вы встретили человека, страдающего альбинизмом, вы бы знали, что у него, скорее всего, есть мутировавший ген TYR, потому что это наиболее частая причина альбинизма. Этот мутировавший ген TYR является частью их генотипа. Альбинизм — часть их фенотипа.
Альбинизм — часть их фенотипа.
Или, допустим, у вас карие глаза.Видимый цвет глаз — это ваш фенотип, но он ничего не говорит нам о вашем генотипе. Множество разных генов влияют на цвет глаз у людей, и любой из них может проявлять доминантные или рецессивные черты вашего фенотипа, то есть уникальный оттенок коричневого в ваших глазах.
Объяснение фенотипа
Фенотип — это то, что вы видите — видимое или наблюдаемое выражение результатов генов в сочетании с влиянием окружающей среды на внешний вид или поведение организма. Все, от формы птичьего крыла до пения горбатого кита, можно рассматривать как часть фенотипа: наблюдаемые аспекты этого животного, которые, по крайней мере частично, определяются его генами.
Примеры фенотипов
Фенотип означает «конкретные результаты генотипа организма». Фенотип, таким образом, можно наблюдать от природы. Каждый признак, определяемый геном, даже частично, является частью его фенотипа. Ниже приводятся лишь некоторые из бесчисленных примеров фенотипов живых существ.
- Цвет глаз
- Цвет волос
- Рост
- Звук вашего голоса
- Отдельные виды болезней
- Размер птичьего клюва
- Длина лисьего хвоста
- Цвет полос на кошке
- Размер и форма пятен на спине собаки
- Размер обуви человека
Мир генов
Гены — это основная структура жизни.Основа биологии — это генетика, а на самом простом уровне генетика — это вопрос генотипа — имеющихся у вас генов — и фенотипа — черт, которые эти гены дают вам. Между ними генотип и фенотип определяют большую часть организма.
Чтобы узнать больше о генетике, ознакомьтесь с нашей статьей о неполном доминировании. Если после всей этой биологии вы настроены на химию, посетите наши страницы, посвященные гетерогенным смесям.
Деконструкция источников ассоциаций генотип-фенотип у людей
Резюме
В последние десятилетия попытки связать вариации генома человека с фенотипами достигли огромных успехов.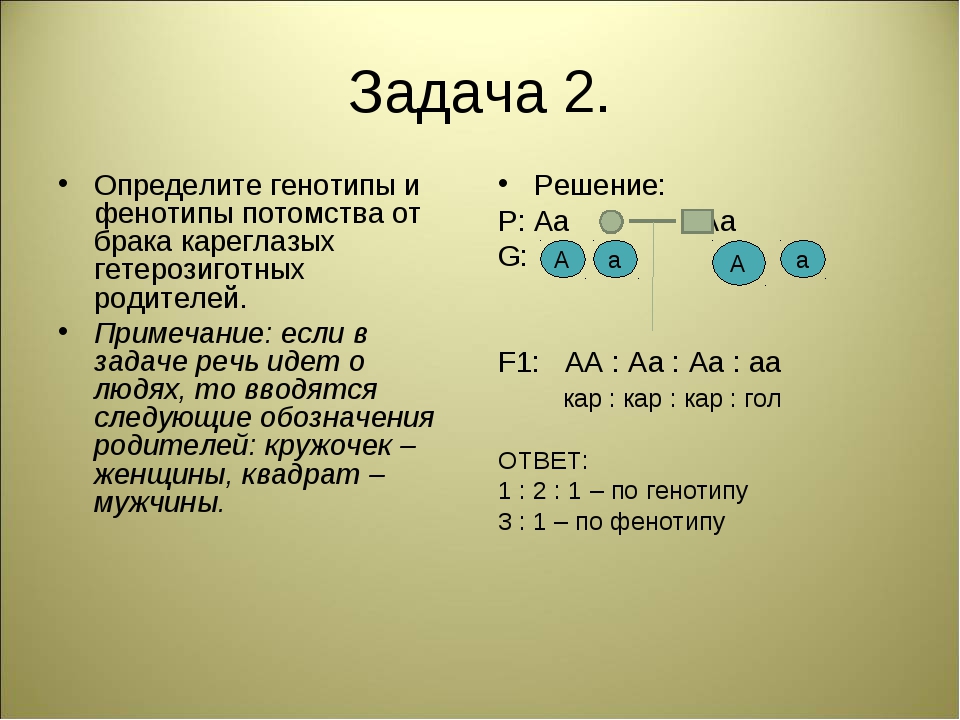 Было показано, что на большинство человеческих черт влияет большое количество генетических вариантов в геноме. Для интерпретации этих ассоциаций и их надежного использования — в частности, для фенотипического предсказания — необходимо лучшее понимание многих источников ассоциаций генотип-фенотип. Мы резюмируем прогресс, достигнутый в этом направлении у людей, особенно в разложении прямых и косвенных генетических эффектов, а также в искажении структуры популяции. Мы обсуждаем естественные следующие шаги в сборе данных и разработке методологии, уделяя особое внимание тому, что можно получить, анализируя данные генотипа и фенотипа от близких родственников.
Было показано, что на большинство человеческих черт влияет большое количество генетических вариантов в геноме. Для интерпретации этих ассоциаций и их надежного использования — в частности, для фенотипического предсказания — необходимо лучшее понимание многих источников ассоциаций генотип-фенотип. Мы резюмируем прогресс, достигнутый в этом направлении у людей, особенно в разложении прямых и косвенных генетических эффектов, а также в искажении структуры популяции. Мы обсуждаем естественные следующие шаги в сборе данных и разработке методологии, уделяя особое внимание тому, что можно получить, анализируя данные генотипа и фенотипа от близких родственников.
Не так давно генетический анализ проводился с использованием значений признаков (фенотипов) в семьях, без генетических данных. Открытие легко измеряемых геномных маркеров позволило идентифицировать гены болезни с помощью анализа сцепления без предварительного знания основных механизмов ( 1 ). Это привело к идентификации гена, ответственного за Х-сцепленное фагоцитарное расстройство хронической гранулематозной болезни в 1986 году, за которым последовали гены других менделевских заболеваний, таких как муковисцидоз ( 2 ) и болезнь Хантингтона ( 3 ), а также гены рака груди ( 4 , 5 ).Этот подход также применялся для изучения общих сложных заболеваний, включая диабет 2 типа, но не смог предоставить воспроизводимых результатов.
Вторая крупная разработка — это массивы однонуклеотидных полиморфизмов (SNP) с высокой пропускной способностью, которые позволили одновременно генотипировать сотни тысяч SNP, что привело к исследованию общегеномных ассоциаций (GWAS) ( 6 ) . GWAS проверяет каждый SNP на связь с фенотипом без данных о семье. Успех GWAS начался с открытия, что CFH способствует возрастной дегенерации желтого пятна; этот анализ был основан на 96 случаях и 50 контрольных ( 7 ).Последующее увеличение размера выборки, которая сейчас составляет более 2 миллионов ( 8 ), привело к открытию тысяч генетических вариантов, влияющих на сотни человеческих черт. Результаты GWAS обещают выявить новые лекарственные цели ( 9 , 10 ), среди других приложений.
Способность GWAS идентифицировать SNP, влияющий на признак, зависит от доли вариации признака, объясняемой SNP, которая увеличивается пропорционально квадрату размера эффекта и гетерозиготности.Поскольку гетерозиготность выше для более распространенных вариантов, первоначальные успехи были в основном для вариантов восприимчивости с частотой минорных аллелей выше 5%. Даже если общий вариант не анализируется напрямую, он, вероятно, сильно коррелирует с генотипированным рядом SNP из-за отсутствия событий наследственной рекомбинации между ними. Эта корреляция называется «локальным» неравновесием по сцеплению (LD). Нелокальная LD — корреляция между вариантами, которые не являются физически близкими — может быть результатом неслучайного спаривания.В результате локальной LD GWAS обычно не идентифицирует непосредственно конкретный причинный вариант, а только определяет его приблизительное геномное положение. Для выявления причинных вариантов требуется мелкомасштабное картирование, которое часто требует функционального анализа и экспериментов ( 11 ).
«Для многих сложных признаков GWAS изменил ландшафт генетических исследований и наше понимание генетической архитектуры»
Большинство распространенных вариантов, обнаруженных GWAS, влияющих на риск заболевания, имеют эффекты от низких до умеренных (увеличивая вероятность заболевания менее чем в 1 раз.5 на аллель риска) ( 12 , 13 ). Применение GWAS для секвенирования всего экзома и всего генома, наряду со статистическим вменением вариантов на уровне последовательности в образцы, генотипированные с помощью массивов SNP, привело к открытию некоторых более редких вариантов с большим эффектом ( 14 ). Хотя дисперсия признаков, объясняемая общегеномными значимыми (GWS) локусами, увеличилась, для большинства сложных признаков дисперсия, объясняемая локусами GWS, составляет лишь часть предполагаемой наследуемости.Этот пробел, названный «отсутствующей наследуемостью», обсуждается ниже ( 12 , 15 ).
Для сложных признаков идентификация всех причинных вариантов и выяснение лежащих в их основе механизмов остается далекой целью. Однако данные GWAS можно использовать для прогнозирования генотипов, особенно с помощью полигенных оценок (PGS). PGS объединяет предполагаемые эффекты нескольких генетических вариантов, чтобы обеспечить прогнозируемое значение признака для человека. Были исследованы многие применения PGS, такие как идентификация людей с существенно повышенным генетическим риском сердечных заболеваний ( 16 ).Несмотря на продемонстрированную ценность PGS, начали появляться вопросы относительно надежности и интерпретации (то есть, что движет предсказательной силой) ( 17 , 18 ).
В GWAS широко признано, что ассоциации могут быть искажены стратификацией населения, в первую очередь ассоциацией между происхождением и влиянием окружающей среды. Методы, корректирующие происхождение, вместе с репликацией ( 19 ), дают уверенность в том, что большинство ассоциаций GWS с общими SNP являются истинно положительными.Однако это не означает, что систематическая ошибка устранена, а природа ассоциаций генотип-фенотип не охарактеризована должным образом. Мы стремимся изложить здесь различные вклады в ассоциацию генотип-фенотип, объяснить трудности, которые они создают, и предложить возможные решения.
Эффекты, фиксируемые ассоциациями GWAS
Связь между генетическим вариантом и фенотипом можно разложить на прямой эффект варианта, косвенный генетический эффект варианта и смешанные эффекты (рис.1). Примером может служить вариант, который оказывает прямое влияние на уровень образования (ОА) при наследовании, а также косвенный эффект через поведение / воспитание родителей ( 20 ). Тот же вариант может иметь косвенное влияние на здоровье через родительское воспитание, но практически не влияет. Прямые эффекты включают в себя широкий спектр причинно-следственных связей, некоторые из которых непростые и не «прямые»; например, варианты в CHRNA5 влияют на риск рака легких через их связь с количеством выкуриваемых ( 21 ).Кроме того, прямой эффект здесь может включать эффекты других вариантов локальной LD. Обратите внимание, что типичный GWAS, проводимый без данных о семье, позволяет оценить только сумму прямого и косвенного воздействия (комбинированный эффект), а не их по отдельности.
Рис. 1 Сигналы, захваченные GWAS отдаленно связанных людей и семей.При оценке степени влияния SNP на признак, основанного на отдаленных родственниках, включаются прямые генетические эффекты (черный цвет), а также ряд других эффектов, в том числе искажение из-за структуры популяции (серый цвет), выборочное спаривание по признаку или коррелированный (бордовый), косвенные генетические эффекты от родителей (синий) и сиб-эффекты (персик).GWAS на основе семьи (например, использование трио) использует родительские генотипы в качестве контроля для разделения прямых и косвенных генетических эффектов и других искажающих эффектов ( 20 ), как показано в разложении справа. На этом рисунке мы игнорируем эффекты локального LD.
КРЕДИТ: KELLIE HOLOSKI / SCIENCE
В рамках аддитивной модели для совместных эффектов вариантов мы определяем генетический компонент как линейную комбинацию генотипов всех причинных вариантов с весами, пропорциональными истинным (прямым, косвенным , или комбинированные) эффекты (рис.2). Генетические компоненты прямого и косвенного воздействия различны, но они могут коррелировать с силой, которая зависит от генетической корреляции между интересующим фенотипом пробанда и фенотипами родственников, через которые опосредуются косвенные эффекты. Например, эта корреляция, вероятно, сильна для EA и слабая для индекса массы тела (ИМТ) ( 20 ). Относительные силы этих двух генетических компонентов и их корреляция определяют корреляции с генетическим компонентом комбинированного эффекта.Поскольку PGS, построенный на основе типичного GWAS, использует оценки комбинированных эффектов, его предсказательная сила иногда может быть значительно сильнее, чем то, что можно объяснить только прямыми эффектами ( 20 ).
Рис. 2 Генетические модели с двумя признаками с объединенными или разделенными прямыми и косвенными эффектами.Для признака, предполагая аддитивную модель, генетический компонент, сочетающий прямые и косвенные эффекты, равен g δ + η = ∑ i (δ i + η i ) g i , где g i , δ i и η i обозначают генотип, прямое и косвенное влияние варианта i соответственно.Вверху: с двумя характеристиками (1 и 2) есть две величины и одна корреляция. Для каждого признака комбинированный генетический компонент может быть разделен на компонент прямого действия, g δ = ∑ i δ i g i , и компонент косвенного воздействия, g η = ∑ i η i g i . Внизу: модель с двумя признаками становится одной с четырьмя генетическими компонентами и шестью попарными корреляциями между ними.Для проиллюстрированного здесь канонического примера, где признак 1 может быть EA, а признак 2 может быть BMI, размер точки указывает величину компонента, а толщина соединительной линии указывает силу корреляции.
Генетические эффекты могут способствовать ассоциации между чертами через плейотропию. Модель с двумя признаками плейотропии (рис. 2, вверху) комбинированных эффектов имеет три параметра: дисперсии, объясняемые генетическими компонентами комбинированного эффекта двух признаков, и корреляцию между ними.Эта корреляция была оценена для многих пар признаков с использованием данных GWAS ( 22 ). Разделив компоненты прямого и косвенного воздействия, модель (рис. 2, внизу) имеет 10 параметров, включая величины четырех генетических компонентов прямого и косвенного воздействия и шесть корреляций. Полная модель не может быть оценена с помощью стандартного GWAS, поэтому в настоящее время у нас мало понимания того, в какой степени прямые и косвенные генетические эффекты по-разному влияют на плейотропию.
Смешивающие эффекты
Связь между генетическим вариантом и фенотипом может частично отражать корреляцию с каким-либо другим причинным феноменом (средовым или генетическим), а не истинным причинным эффектом SNP на фенотип.Этот тип смешения возникает из-за наличия неслучайного спаривания, приводящего к структуре населения. В GWAS есть по крайней мере три источника искажения: (i) искажение окружающей среды, когда частоты аллелей и эффекты среды коррелированно варьируются в разных географических регионах или субпопуляциях; (ii) генетическое смешение, когда различия частот аллелей между субпопуляциями коррелируют с различиями частот других аллелей с причинными эффектами; или (iii) смешивание ассортативного спаривания, которое происходит, когда есть ассортативное спаривание по признаку (или коррелированному признаку), вариант с причинным эффектом на признак становится коррелированным с другими вариантами с причинными эффектами, и его связь с признаком затем фиксирует свой собственный причинный эффект плюс часть других вариантов.Эти формы смешения концептуально различны, но на практике они часто взаимосвязаны.
Поправка на искажение в GWAS
Корректировка на главный компонент (PC) — это распространенный метод, используемый для устранения некоторых искажающих эффектов, связанных со структурой популяции ( 23 ). В идеале основные компоненты, используемые для корректировок, сильно коррелируют с компонентом, влияющим на окружающую среду, и не коррелируют с компонентом прямого генетического воздействия. Если компонент прямого воздействия существенно коррелирует с искажающими компонентами, корректировка ПК удалит некоторые прямые генетические эффекты, а также смешанные эффекты.
Смешивающий компонент ассортативного спаривания (iii) по своей природе почти идеально коррелирует с суммой прямых и косвенных компонентов. Ассортативное спаривание по таким признакам, как рост и EA ( 24 ) приводит к нелокальному LD вариантов с прямыми и косвенными эффектами, которые улавливаются PC. Таким образом, теоретически настройка ПК может отрегулировать большую часть компонента прямого воздействия. На практике этого не происходит. Даже при очень большом размере выборки предполагаемые ПК, скорее всего, будут в основном шумными, помимо нескольких сильных (часто географических) сигналов.Результаты выборки белых британцев (WB) UKB подчеркивают этот момент (рис.3): помимо первых восьми самых сильных ПК, ПК, вычисленные из выборки из 272 519 человек ( 25 ), по-видимому, в основном обусловлены шумом выборки и локальным LD. в хромосомах. Шум может маскировать тонкую структуру населения, что может привести к искажению GWAS даже после настройки ПК ( 26 ).
Рис. 3 Поведение основных компонентов 272 519 образцов биобанка Великобритании.Мы исследуем степень, в которой основные компоненты отражают реальную структуру популяции, исследуя, воспроизводится ли генетическая дисперсия (собственные значения), объясняемая 40 основными компонентами, выведенными из 146082 SNP в 272 519 британских образцах биобанка белых британцев (WB), в независимой выборке. ВБ.Собственное значение репликации выше 1 указывает на то, что предполагаемый главный компонент фиксирует воспроизводимые корреляции между SNP, либо локальной LD (в пределах хромосомы), либо структурой популяции (в основном между хромосомами). Исходный (черные кружки): собственные значения основных компонентов в исходном наборе из 272 519 человек из ВБ. Репликация (синие треугольники): эквиваленты собственных значений, то есть дисперсии линейных комбинаций генотипов SNP с использованием весов, выведенных из исходного набора, и стандартизованных генотипов в наборе репликации из 64 969 индивидуумов WB.Репликация (только между хромосомами) (красные кресты): с использованием того же набора репликации, но эквиваленты собственных значений, вычисленные путем игнорирования ковариаций пар SNP в одних и тех же хромосомах и подсчета только ковариаций пар SNP на разных хромосомах, что включает 94,8% все пары SNP. Среднее собственное значение для последних 32 ПК снижается с 4,37 для исходного набора до 2,61 для набора репликации и далее до 1,03 для набора между хромосомами, указывая на то, что эти ПК в основном улавливают шум и локальную LD, а не структуру популяции.
Фитинг линейных смешанных моделей (LMM) является альтернативой настройке с помощью ПК. Эти методы выполняют тип регрессии для набора SNP, где эффект каждого SNP моделируется как «случайный эффект», полученный из нормального распределения ( 27 ). LMM уже давно используются для прогнозирования признаков в селекции животных ( 28 ). В исследованиях на людях тестирование ассоциации LMM обычно состоит из оценки эффекта фокусного SNP как «фиксированного эффекта» при моделировании случайных эффектов для набора других SNP.Наивные вычисления LMM масштабируются с кубом размера выборки, и поэтому были разработаны альтернативные вычислительные подходы для обработки больших наборов выборок GWAS ( 29 ).
Привлекательность LLM заключается в том, что они позволяют улучшить моделирование стратификации населения и родства выборки ( 27 ). LMM часто используются в сочетании с корректировкой PCA и могут учитывать более сложные модели стратификации путем моделирования эффектов (почти) всех измеренных SNP, фиксируя как реальные генетические эффекты, так и эффекты стратификации ( 27 ).Кроме того, методы LMM могут улучшить оценку эффектов SNP и их ошибок выборки по сравнению с линейной регрессией при наличии родства выборки ( 27 ). LMM могут также уменьшить смещение в оценках эффекта SNP из-за ассортативного спаривания ( 30 ). Однако современные методы LMM GWAS не устраняют косвенных генетических эффектов.
Использование данных генотипа семьи
Учитывая родительские генотипы, генотип потомства определяется путем случайной сегрегации генетического материала во время мейоза.Эта случайная сегрегация не коррелирует с косвенными генетическими эффектами от родственников и другими смешивающими эффектами. Таким образом, родительские генотипы можно использовать в качестве контроля для получения объективных оценок прямых генетических эффектов ( 20 , 31 ) (рис. 1). Точно так же генетические различия между братьями и сестрами являются результатом случайной менделевской сегрегации родителей во время мейоза. Следовательно, генетические различия между братьями и сестрами не смешиваются с косвенными генетическими эффектами родителей, стратификацией популяции и ассортативным спариванием.Однако методы, использующие различия в генотипах братьев и сестер, оценивают прямой эффект за вычетом косвенного эффекта от брата или сестры, и, следовательно, дают объективные оценки прямого эффекта только в том случае, если косвенный генетический эффект брата или сестры равен нулю.
Изучение косвенных генетических эффектов имеет долгую историю в животноводстве ( 13 ). У людей в большинстве исследований косвенных генетических эффектов использовались PGS, полученные из GWAS, которые не различают прямые и косвенные генетические эффекты (рис.4) ( 20 ). Однако, когда прямые и косвенные генетические эффекты не полностью коррелированы, этот подход не может дать полной картины ( 32 ). В идеале GWAS следует проводить с генотипами родителей или братьев и сестер в качестве контроля и с использованием моделей с косвенными генетическими эффектами. Однако возможности этого подхода в настоящее время ограничены, поскольку большие выборки с генотипированными братьями и сестрами и / или родителями встречаются редко. Кроме того, поскольку только около половины генетической изменчивости в популяции является внутрисемейной, для получения такой же исследовательской мощности, как и при стандартном GWAS-анализе, требуются значительно более крупные выборки семей.Следовательно, необходимы методы, объединяющие информацию из стандартных GWAS и из анализа семейств.
Рис. 4 Сокращение полигенного прогноза и оценок наследуемости с использованием внутрисемейных планов с использованием исландских данных.( A ) Оценка наследуемости SNP с использованием переданных аллелей дается hSNP2; оценка наследуемости SNP с использованием внутрисемейного метода, регрессии неравновесия по родству (RDR) ( 31 ), дается с помощью hRDR-SNP2.Статистически значимые различия ( P <0,05, односторонний z-тест) наблюдались для EA hSNP2 / hRDR-SNP2 = 1,72 ( P = 7,6 × 10 −3 ) и роста hSNP2 / hRDR-SNP2 = 1,24 ( P = 0,015). ( B ) Дисперсия, объясняемая регрессией признака на полигенную оценку, дается Rpoly2; дисперсия, объясняемая полигенной оценкой, когда ее эффект оценивается с использованием внутрисемейного (трио) дизайна, определяется как Rpoly: δ2 ( 20 ). Мы подчеркиваем относительный размер оценок от внутрисемейных методов (hRDR-SNP2 и Rpoly: δ2) до межсемейных методов (hSNP2 и Rpoly2).Межсемейные методы фиксируют косвенные генетические эффекты от родственников и, возможно, стратификацию популяции и ассортативное спаривание в дополнение к наследуемости, зафиксированной внутрисемейными методами. Сокращения для признаков: ИМТ, индекс массы тела; EA, уровень образования (лет).
Наследственность
Традиционно наследуемость оценивалась путем сравнения корреляций между однояйцевыми и неидентичными близнецами. В дополнение к идентификации конкретных причинных локусов можно использовать данные GWAS для оценки фенотипической изменчивости, объясняемой генетической изменчивостью, захваченной SNP (и вариантами в LD с ними) на массиве генотипирования, называемом «наследуемость SNP» или hSNP2 ( 33 ).Оценки hSNP2 подразумевают, что общие генетические варианты, проанализированные на типичном массиве генотипов, в совокупности объясняют значительно большую фенотипическую дисперсию, чем варианты GWS. Однако оценки hSNP2, как правило, значительно ниже оценок наследуемости из исследований близнецов ( 15 ), что является частью «проблемы отсутствия наследственности». Некоторые, но далеко не все, этот разрыв объясняется эффектами вмененных вариантов, которые не находятся в сильной LD с маркерами на типичном массиве генотипов ( 13 , 34 ).Одна из возможностей состоит в том, что большая часть оставшейся недостающей наследуемости объясняется очень редкими вариантами ( 35 ).
Широко используемый метод GREML оценивает hSNP2 путем измерения силы взаимосвязи между фенотипическим сходством и геномным генетическим сходством (оценивается по SNP), которая варьируется даже для отдаленных родственников, обычно используемых в GWAS ( 36 ) . Этот подход обеспечивает оценку общей дисперсии, объясняемой комбинированными прямыми и косвенными эффектами аллелей пробандов ( 20 , 31 ).Степень, в которой косвенные генетические эффекты и стратификация популяции повлияли на оценки hSNP2 (рис. 4), не известна, равно как и систематическая ошибка, вызванная ассортативным скрещиванием при оценке наследуемости как внутри семьи, так и между семьей.
Также важно отметить, что общая дисперсия, объясняемая комбинированными прямыми и косвенными эффектами, отличается от традиционно определенной наследуемости, которая касается только прямых эффектов. Тем не менее, это интересный параметр, поскольку он определяет верхнюю границу генетического предсказания на основе аллелей пробандов.Подразумевается, что верхний предел генетического предсказания признака часто может быть больше, чем наследуемость ( 18 ).
Некоторые недавние методологические разработки
Регрессия оценки LD
С появлением GWAS были разработаны подходы, позволяющие лучше использовать и интерпретировать их результаты. Примечательно, что регрессия по шкале LD (LDSC) была разработана для того, чтобы отличить эффекты смешения из-за стратификации населения от причинно-следственных генетических эффектов на статистику теста GWAS ( 37 ).Предполагая высокополигенную архитектуру, ожидается, что статистика теста GWAS для отдельного SNP будет увеличиваться с увеличением его показателя LD (мера генетической вариации, помеченной SNP через локальную LD) из-за увеличения корреляции с причинными вариантами. Однако средняя статистика теста по всем SNP повышается при стратификации популяции из-за корреляции между аллелями и различий в средних значениях признаков между субпопуляциями ( 37 — 39 ). Оценивая, насколько искажение, вызванное стратификацией населения, увеличивает среднюю статистику теста, точку пересечения LDSC можно использовать для корректировки статистики теста GWAS.LDSC также можно использовать для оценки корреляции между эффектами SNP на разные признаки ( 22 ), для разделения вкладов в наследуемость SNP от различных функциональных категорий вариантов ( 40 ) и для облегчения многопоточного метаанализа ( 41 ). ).
Ключевое предположение LDSC состоит в том, что различия частот аллелей между субпопуляциями не зависят от баллов LD ( 37 ). Однако корреляция между оценками LD и различиями частоты аллелей может быть вызвана формами связанного отбора, такими как фоновый отбор ( 26 ).Таким образом, остаются вопросы о надежности меры смещения стратификации населения LDSC.
Менделирующая рандомизация
Менделирующая рандомизация (MR) использует генетические данные для улучшения причинно-следственных связей в эпидемиологии ( 42 ). Если генетический вариант влияет на признак A, а признак A влияет на признак B, то ожидается, что варианты, которые влияют на признак A, будут влиять на признак B. Генетические варианты, которые влияют на признак A, могут быть использованы для определения того, отражает ли ассоциация между признаком A и признаком B причинное влияние признака A на признак B, учитывая, что генетические варианты влияют на признак B только через свое влияние на признак A, и что генетические варианты не коррелируют с какими-либо смешивающими факторами.MR доказала свою эффективность в опровержении ложных причинно-следственных гипотез, основанных на данных наблюдений, таких как связь между уровнем холестерина ЛПВП и сердечно-сосудистыми заболеваниями ( 43 ) и сниженным риском сердечно-сосудистых заболеваний у умеренно пьющих в западных обществах ( 44 ).
MR обычно полагается на оценки эффекта SNP из GWAS без семей, которые могут быть искажены стратификацией популяции, косвенными генетическими эффектами от родственников и ассортативным спариванием ( 45 ).Для решения этих проблем были предложены внутрисемейные методы МРТ, которые показали, что предыдущие оценки МРТ причинных эффектов роста и ИМТ на ЭА были ложными ( 45 ).
Еще одной проблемой для MR-анализа является широко распространенная плейотропия: если SNP влияет на признак B через признак, отличный от признака A, то он не является действенным инструментом для вывода о причинном влиянии признака A на признак B. разработаны для решения этой проблемы, их эффективность может зависеть от предварительных знаний о путях смешивания ( 46 ).
Взаимодействие генов с окружающей средой
Взаимодействие генов с окружающей средой (GxE) происходит, когда влияние генетического варианта на признак различается в разных средах ( 47 ). Такие взаимодействия GxE отличаются от корреляции ген-среда, которая может быть результатом, например, косвенных генетических эффектов от родственников. У людей хорошо реплицированные примеры взаимодействий GxE редко встречаются вне фармакогеномики ( 48 , 49 ). Единственным исключением является взаимодействие между вариантами в локусе FTO и физической активностью, влияющей на ИМТ ( 50 , 51 ).
Возможности обнаружения взаимодействий GxE в исследованиях GWA, вероятно, были низкими из-за небольшого размера эффекта и нагрузки на несколько тестов. Один из способов увеличить мощность обнаружения GxE — это поиск взаимодействий между факторами окружающей среды и PGS ( 52 , 53 ). Этот метод эффективен, когда генетические варианты, влияющие на признак, аналогичным образом взаимодействуют с факторами окружающей среды, но не могут идентифицировать взаимодействия между факторами окружающей среды и конкретными генетическими вариантами.LMM могут применяться для обнаружения компонента фенотипической изменчивости, возникающего в результате взаимодействия между геномными генетическими вариантами и фактором окружающей среды ( 54 ), но не могут точно определить взаимодействия с конкретными генетическими вариантами. Генетические варианты, участвующие во взаимодействиях GxE, влияют на изменчивость признака ( 55 , 56 ), что может быть использовано для сокращения пространства поиска потенциальных взаимодействий путем ограничения вариантов с доказательствами влияния на фенотипическую изменчивость.Однако остаются методологические проблемы: эффекты взаимодействия и генетические эффекты на фенотипическую изменчивость чувствительны к масштабу измерения ( 56 , 57 ), а влияние стратификации популяции на оценки GxE недостаточно охарактеризовано. Более того, причинность эффектов взаимодействия GxE установить сложно, поскольку взаимодействие может быть связано с неизмеряемым фактором окружающей среды, который коррелирует с измеренным фактором (факторами) окружающей среды, а более широкие социально-экологические факторы, которые могут структурировать воздействие окружающей среды, часто неизвестны.
Переносимость фенотипического предсказания
Точность предсказания на основе PGS зависит от наследуемости признака и мощности существующего GWAS (особенно от размера выборки и генетической архитектуры) ( 28 ). Для нескольких признаков [таких как рост, для которого текущая точность прогноза составляет ~ 25% ( 58 )], существующие оценки уже являются информативными для групп людей, аналогичных тем, для которых проводился GWAS.
Полигенные оценки не работают также при прогнозировании фенотипов людей, которые отличаются от тех, которые включены в набор GWAS.Некоторые причины понятны и возникают из-за различий в происхождении. Примечательно, что поскольку PGS состоит из взвешенной суммы количества аллелей и поскольку частоты аллелей варьируются по всему миру (из-за генетического дрейфа и естественного отбора), аллели, которые вносят вклад в вариацию признаков в GWAS, с меньшей вероятностью присутствуют или могут даже отсутствовать. у более дальних родственников. Также ожидается, что точность предсказания PGS будет снижаться по группам предков, потому что GWAS идентифицирует не причинные сайты, а наборы возможных причинных сайтов в локальной LD; поскольку локальные паттерны LD зависят от истории популяции, ассоциации, наблюдаемые в одной популяции, будут иметь тенденцию хуже улавливать причинные SNP в других.Как и ожидалось, недавние исследования сообщают, что инкрементный R 2 для широкого диапазона признаков ниже у людей, чьи предки отличаются от предков из набора GWAS ( 59 , 60 ).
Помимо частоты аллелей и различий LD, другие факторы могут способствовать снижению прогностической способности PGS: степень дисперсии окружающей среды может различаться среди групп разнородных предков или выбираться по отдельным критериям включения ( 18 ), а измерение фенотипа может отличаться. по группам.Более того, величина эффекта вариантов может различаться в результате взаимодействий ген-ген (GxG) и GxE. Изменения в величине эффекта могут быть особенно важны для признаков, в которые косвенные эффекты или ассортативное спаривание вносят большой вклад, поскольку такие факторы могут зависеть от культурных и экологических факторов. Здесь становится важным разложить природу сигналов, идентифицированных в GWAS, чтобы определить, какие компоненты (например, прямые и косвенные эффекты) обеспечивают более легко обобщенные прогнозы.
Outlook
Для многих сложных признаков GWAS изменил ландшафт генетических исследований и наше понимание генетической архитектуры. Если раньше не было ни одной надежно воспроизводимой ассоциации, теперь существуют тысячи вариантов с надежными ассоциациями. Примечательно, что GWAS не требует данных о семье, что облегчает сбор больших размеров выборки. Однако в последнее время на первый план снова выдвигаются уникальные свойства семейных данных. Во-первых, некоторые редкие варианты с сильными эффектами существуют только в расширенных семьях.Наиболее важно то, что для более глубоких и тонких вопросов могут потребоваться семейные данные, такие как тройки родителей и потомков и пары братьев и сестер, чтобы отличить прямые эффекты от косвенных и других смешивающих факторов. Статистически одно естественное расширение — это расширение исследуемой единицы от индивидуума до нуклеарной семьи. В этой связи стоит отметить, что по мере увеличения размера выборки неизбежно будут собираться близкие родственники, поскольку в выборку входят более крупные фракции населения.
Остающейся проблемой является проблема систематической ошибки при установлении, которая возникает, когда исследуемые выборки систематически отличаются от совокупности.Большинство наборов выборок смещено в сторону лиц европейского происхождения ( 60 ), а также лиц с более высоким социально-экономическим статусом и более здоровым ( 61 ), наряду с другими неизвестными предубеждениями. Хотя эти предубеждения при установлении не обязательно приводят к ложноположительным результатам, они ограничивают переносимость результатов GWAS ( 18 , 60 ). Особенно заметными в этом отношении являются взаимодействия GxE не только в пространстве, то есть между популяциями в данный момент времени, но и во времени, учитывая массивные вековые тенденции в окружающей среде, которые произошли и продолжают происходить.Это соображение относится к чертам здоровья, чертам, связанным с образованием, и чертам плодородия, которые влияют на давление отбора. В связи с этим важно не только собирать образцы разных предков и текущей среды, но, где это возможно, также собирать данные по нескольким поколениям.
Box 1Глоссарий.
Ассортативное спаривание: Когда пары, производящие потомство, выбирают друг друга на основе определенных фенотипов.
Картирование в мелком масштабе: Относится к подходам, которые направлены на определение того, какой вариант или варианты могут быть причинными среди набора связанных вариантов, идентифицированных в GWAS.
Наследственность: Измеряет долю фенотипической изменчивости, объясняемую прямым воздействием всех генетических вариантов в популяции в данный момент времени.
Гетерозиготность: Вероятность того, что два аллеля на сайте различаются; в предположении равновесия Харди-Вайнберга и с учетом двуаллельного сайта эта мера генетического разнообразия определяется как 2 p (1 — p ), где p — частота аллеля.
Вменение: Статистический метод, позволяющий вывести генотипы людей в вариантах, не измеренных напрямую в массиве генотипов, путем ссылки на полные данные последовательности генома.
Косвенный генетический эффект: Влияние генетического варианта у одного человека на черты другого человека через окружающую среду.
Общегеномные значимые ассоциации (GWS): Варианты, связанные с фенотипом на уровне значимости, выбранном для преодоления бремени множественного тестирования, обычно устанавливается на P <5 × 10 –8 .
Анализ сцепления: Тесты на совместную сегрегацию фенотипов и генотипов внутри семей.
Плейотропия: Распространенное наблюдение, что многие SNP, связанные с одним признаком, также связаны с другими признаками. Относится к концепции генетической корреляции.
Главный компонент: Главный компонент — это предполагаемая ось генетической изменчивости в выборке. Главный компонент — это линейная комбинация генотипов SNP, где каждый SNP имеет «нагрузку», дающую свой вклад в главный компонент.
Полигенная оценка (PGS): Взвешенная сумма аллелей, переносимых индивидуумом, где веса даны по величине эффекта, оцененной в GWAS.
Благодарности: Спасибо Д. Конли, А. Харпаку и Дж. Притчарду за комментарии к черновику рукописи. При поддержке Фонда Ли Ка Шинга (A.I.Y., S.B. и A.K.) и гранта NIH R01 GM121372 (M.P.). Это исследование было проведено с использованием ресурса UK Biobank Resource под номером заявки 11867.
Генотип против фенотипа | Биологический словарь
Генетический состав организма называется его генотипом, а наблюдаемые признаки — его фенотипом. Другими словами, то, как человек выглядит, является результатом его или ее ДНК. Прекрасным примером, объясняющим эту концепцию, являются эксперименты Грегора Менделя с чистыми растениями желтого и зеленого гороха (P 1 ). Фенотип растений — их цвет, желтый и зеленый. Когда Мендель скрестил эти два растения, все потомство (F 1 ) было желтым.Но зеленый цвет не был потерян, так как он снова появился в следующем поколении (F 2 ). В то время Мендель этого не знал, но лежащие в основе генотипы зеленых и желтых растений были ответственны за черты, наблюдаемые в поколении F 1 .
И зеленые, и желтые растения гороха P 1 имели идентичные аллели гена, контролирующего их цвет, то есть они были гомозиготными. Когда два зеленых или два желтых растения были скрещены вместе, все потомки зеленых растений были зелеными, а все потомки желтых растений были желтыми.Однако при скрещивании зеленого растения с желтым все потомство F 1 было желтым. Это связано с тем, что поколение F 1 было гетерозиготным по цветному признаку, то есть у них были разные аллели по цвету. И из двух аллелей гена окраски желтый был доминирующим, что делало все растения F 1 желтыми. Аллели гороха в F 2 должны быть гомозиготными рецессивными, чтобы растение было зеленым
Сравнительная таблица
| Генотип | Фенотип | |
|---|---|---|
| Наблюдаемые признаки организма? | Нет | Да |
| Генетический состав организма? | Да | Нет |
| Как это наблюдается? | Секвенирование ДНК, анализ ПЦР, если необходимо (см. Изображение ниже) | Посмотрев на организм |
| Относится к информации, закодированной по аллелям | Да | Нет |
| Примеры включают цвет волос, цвет глаз , высота | Нет | Да |
| По наследству? | Частично, по одному аллелю от каждого родителя | Нет |
| Что он содержит (показать)? | Вся информация о наследственности организма, даже о невыраженных генах. | Только экспрессируемые гены. |
На изображении выше показано, как работает метод полимеразной цепной реакции (ПЦР) для амплификации небольших образцов ДНК для генетического анализа. Генетический анализ выявляет генотип человека, который кодирует наблюдаемые характеристики, то есть фенотип.
Ссылки
- Генотип против фенотипа. (нет данных). В Diffen.com . Получено с https://www.diffen.com/difference/Genotype_vs_Phenotype
- OpenStax.(20 мая 2013 г.). Эксперименты Менделя в наследственности. В Биология . (Глава 12). Получено с http://cnx.org/content/col11448/latest/
Подпишитесь на нашу рассылку новостей
Генотип и фенотип — Центр научного обучения
Мы все уникальны. Даже у монозиготных близнецов, которые генетически идентичны, всегда есть некоторые различия в том, как они выглядят и действуют. Эта уникальность является результатом взаимодействия нашей генетической структуры, унаследованной от наших родителей, и влияния окружающей среды с момента нашего зачатия.
Понимание генотипа и фенотипа
Вильгельм Йоханссен был ученым, работавшим в Дании в конце 19 — начале 20 веков. В ходе серии экспериментов он наблюдал вариации генетически идентичных бобов. Он пришел к выводу, что вариация должна быть вызвана факторами окружающей среды, и ввел термины «генотип» и «фенотип» в 1911 году.
Генотип
Генотип — это генетическая структура отдельного организма. Ваш генотип функционирует как набор инструкций для роста и развития вашего тела.Слово «генотип» обычно используется, когда говорят о генетике определенного признака (например, цвета глаз).
Фенотип
Фенотип — это наблюдаемые физические или биохимические характеристики отдельного организма, определяемые как генетическим составом, так и влиянием окружающей среды, например ростом, весом и цветом кожи.
Как генотип влияет на фенотип
Термин «генотип» обычно используется для обозначения определенных аллелей. Аллели — это альтернативные формы одного и того же гена, которые занимают одно и то же место на хромосоме.В любом данном локусе есть 2 аллеля (по 1 на каждой хромосоме в паре) — вы получаете 1 аллель от матери и 1 от отца. 2 аллеля могут быть одинаковыми или разными. Различные аллели гена обычно выполняют одну и ту же функцию (например, они кодируют белок, влияющий на цвет глаз), но могут давать разные фенотипы (например, голубые или карие глаза) в зависимости от того, какой набор из двух аллелей у вас есть.
Например, способность ощущать вкус PTC (горького соединения) контролируется одним геном.У этого гена как минимум 7 аллелей, но обычно встречаются только 2 из них.
Заглавная буква «T» представляет доминантный аллель, который дает способность ощущать вкус — «доминантный» означает, что любой, у кого есть 1 или 2 копии этого аллеля, сможет попробовать PTC. Аллель без дегустации является рецессивным и обозначается строчной буквой «t» — «рецессивный» означает, что человеку потребуется 2 копии аллеля, чтобы не тестировать.
Каждая пара аллелей представляет генотип конкретного человека, и в этом случае существует 3 возможных генотипа: TT (тестер), Tt (тестер) и tt (не тестер).
Если аллели совпадают (TT или tt), генотип гомозиготный. Если аллели различны (Tt), генотип гетерозиготен.
На самом деле редко, когда 1 ген определяет 1 признак, как в случае дегустации PTC (моногенный признак). Большинство признаков сложны и имеют гены, влияющие на них более чем в 1 локусе (полигенный).
Как факторы окружающей среды влияют на фенотип
Ваши гены несут в себе инструкции для роста и развития вашего тела.Однако на ваш фенотип во время эмбрионального развития и на протяжении всей вашей жизни влияют факторы окружающей среды. Факторов окружающей среды много и они разнообразны и включают диету, климат, болезни и стресс.
На примере дегустации PTC ученые подсчитали, что ген контролирует около 85% способности ощущать вкус. Факторы окружающей среды, которые играют роль, включают сухость во рту и время, когда вы ели.
Степень, в которой ваш фенотип определяется вашим генотипом, называется «фенотипической пластичностью».Если факторы окружающей среды имеют сильное влияние, фенотипическая пластичность высока. Если генотип можно использовать для надежного прогнозирования фенотипа, фенотипическая пластичность низкая.
В целом, степень влияния факторов окружающей среды на ваш конечный фенотип — это горячо обсуждаемая научная проблема. Это часто называют спорами «природа (гены) против воспитания (окружающей среды)». Ученые обычно изучают монозиготных (однояйцевых) близнецов, чтобы исследовать взаимосвязь генотип / фенотип.
В заключение скажу, что ваш генотип или генетический состав играет решающую роль в вашем развитии. Однако факторы окружающей среды влияют на наши фенотипы на протяжении всей нашей жизни, и именно постоянное взаимодействие между генетикой и окружающей средой делает всех нас уникальными.
Фенотип и генотип — определение, 10 основных различий, примеры
Главная »Разница между» Фенотип и генотип — определение, 10 основных различий, примеры
Последнее обновление: 30 декабря 2020 г., автор: Sagar Aryal
Изображение создано с помощью biorender.com Определение фенотипа- Фенотип — это термин, используемый в генетике для обозначения всех наблюдаемых признаков у организмов в результате взаимодействия генотипа с окружающей средой.
- Термин «фенотип» в «фенотипе» относится к «наблюдаем», и, таким образом, фенотип используется для обозначения наблюдаемых характеристик организмов, таких как его рост и цвет.
- Фенотип организма охватывает морфологию, физическую форму и структуру организма, его развитие и поведение, его биологические и физиологические свойства и даже продукты организма.
- Таким образом, фенотипы используются для различения различий в последовательностях ДНК между людьми с различиями в таких признаках, как рост.
- Фенотип организма зависит от двух факторов; экспрессия генома или его генотипа и его взаимодействие с факторами окружающей среды.
- Один или оба фактора могут влиять на фенотип организма.
- Изменения фенотипа наблюдаются даже у индивидуума из-за экологических, физиологических и морфологических изменений, связанных с возрастом.
- Вариация фенотипа формирует основу естественного отбора, при котором окружающая среда благоприятствует выживанию более приспособленных особей по сравнению с другими.
- Этот феномен четко прослеживается в случае близнецов, когда люди с одинаковыми генотипами могут выражать разные фенотипы, если каждый человек сталкивается с разной средой.
- Таким образом, без фенотипической изменчивости эволюция путем естественного отбора была бы невозможна.
- Концепция вариации фенотипа может действовать ниже генетической структуры человека.Примером этого являются молчащие мутации, которые не изменяют аминокислотные последовательности, но изменяют частоту пар оснований гуанин-цитозин.
- Это изменение влияет на содержание G-C в геноме, что, в свою очередь, вызывает повышение термической стабильности, позволяя организму жить в высокотемпературной среде.
- Примеры фенотипов, наблюдаемых у различных организмов, включают группу крови, цвет глаз и текстуру волос, а также генетические заболевания у людей, размер стручка и цвет листьев, клюв птиц и т. Д.
- Генотип — это термин, используемый в генетике для обозначения генетического состава человека, состоящего из наследуемых генов.
- Термин «генотип» также относится к двум аллелям, которые наследуются для определенного гена.
- Генотипы представлены буквенными буквами, например Bb, где B обозначает доминантный аллель, а b обозначает рецессивный аллель.
- Физические характеристики человека, называемые фенотипами, являются результатом экспрессии этих генов.
- Однако генотип — не единственный фактор, ответственный за различие в фенотипе, и на фенотип также влияют другие факторы, такие как наследственная эпигенетика и факторы окружающей среды.
- Таким образом, ни все люди с одним и тем же генотипом не выглядят одинаково, ни все люди, которые похожи друг на друга, не имеют одинаковый генотип.
- На генотипы организмов влияют различные факторы, такие как мутации, которые изменяют частоту пар оснований или структуру последовательностей ДНК.
- Однако гены, образованные в результате соматических мутаций, которые приобретаются, а не наследуются, не рассматриваются как генотип человека. Эти мутации также не вызывают никаких изменений генотипов.
- Экспрессия генотипов в организмах зависит от состава генотипа. При наличии доминантного аллеля потомство обязательно наследует доминантные признаки независимо от другого аллеля.
- Генотип персонажа полностью зависит от последовательностей генов, за исключением случайных наследственных мутаций.
- Таким образом, генотип остается неизменным на протяжении всей жизни человека, если он не затронут мутациями.
- Точно так же фенотипические изменения у индивидуума не вызывают никаких изменений генотипа индивидуума, поскольку на генотип не влияет фенотип.
- Генотипы, как и фенотипы, не наблюдаются извне. Вместо этого должны выполняться такие процессы, как генотипирование.
- Процесс определения генотипа человека называется генотипированием, которое может использоваться для различных целей, таких как ПЦР, секвенирование ДНК и полиморфизм длины рестрикционных фрагментов (RFLP).
- Примеры генотипов, наблюдаемых у разных животных, включают TT как гомозиготный аллель роста, Tt как гетерозиготный аллель роста и BB как гомозиготный аллель цвета глаз.
| Основа для сравнения | Фенотип | Генотип |
| Определение | Фенотип — термин, используемый в генетике для обозначения всех наблюдаемых признаков у организмов в результате взаимодействия генотипа с окружающей средой. | Генотип — это термин, используемый в генетике для обозначения генетического состава человека, состоящего из наследуемых генов. |
| Наблюдаемый | Фенотипы — это наблюдаемые черты, которые можно увидеть на теле организма. | Генотипы присутствуют в хромосомах индивидов и поэтому не наблюдаются. |
| Унаследовано | Фенотипы не передаются по наследству. | Генотипы частично передаются от человека к потомству как один из двух аллелей во время полового размножения. |
| Состоит из | Фенотип состоит из различных черт, таких как физическая форма и структура, его развитие и поведение, его биологические и физиологические свойства и даже продукты организма. | Генотип состоит из наследственных характеристик организма, которые могут или не могут быть выражены в следующем поколении. |
| Затронуты | На фенотип влияет генотип организма и другие условия окружающей среды. | На генотип влияет генетический состав особи, который, в свою очередь, зависит от полового размножения. Наследственные мутации также могут повлиять на генотип организма. |
| Отношение | Один и тот же фенотип может или не может быть выражен из одного и того же генотипа. | Один и тот же генотип всегда приводит к одному и тому же фенотипу, за исключением наследственных мутаций. |
| Определяется по | Фенотипы можно легко определить, наблюдая за организмом. | Генотипы определяются в процессе генотипирования с помощью различных научных методов, таких как полимеразная цепная реакция (ПЦР) и ПДРФ. |
| Факторы окружающей среды | На фенотипы влияют факторы окружающей среды. | Генотипы не подвержены влиянию факторов окружающей среды. |
| Изменения | Фенотипы могут меняться в течение жизни человека, например, цвет волос. | Генотипы остаются неизменными на протяжении всей жизни человека. |
| Примеры | Примеры фенотипов, наблюдаемых у различных организмов, включают группу крови, цвет глаз и текстуру волос, а также генетические заболевания у людей, размер стручка и цвет листьев, клюв птиц и т. Д. | Примеры генотипов, наблюдаемых у разных животных, включают TT как гомозиготный аллель роста, Tt как гетерозиготный аллель роста и BB как гомозиготный аллель цвета глаз. |
- Производство меланина в организме человека контролируется определенными генами; таким образом, различие в производстве меланина связано с различиями в генотипах организмов.
- Хотя разные гены контролируют распределение меланина по телу, существует единственный ген, который влияет на его выработку.
- Таким образом, люди с определенным генотипом могут вообще не страдать от выработки меланина, что приводит к альбинизму.
- Таким образом, у людей с альбинизмом обычно белая или розовая кожа.
- Альбинизм — это фенотип, являющийся результатом определенного генотипа, и поскольку генофонд альбинизма довольно велик, случаи альбинизма наблюдаются во многих популяциях.
- Альбинизм может даже быть распространенным у некоторых животных.
- В рамках своих исследований Мендель изучил ряд фенотипов гороха. Почти все сделанные им открытия были основаны на этих фенотипах.
- В случае цвета гороха исследовали зеленый и желтый горох. Он обнаружил, что селекция гороха желтого и зеленого цвета привела к получению половины желтого и половины зеленого горошка с некоторыми вариациями в некоторых поколениях.
- На основании этого он определил соотношение разных фенотипов в разных поколениях.
- Окраска гороха кодируется определенным геном, который приводит к желтому цвету. В отсутствие этого гена цвет гороха становился зеленым.
- Таким образом, желтый цвет является доминантным аллелем, а зеленый — рецессивным аллелем.
- Доминирующим генетическим признаком цвета глаз является коричневый, который представлен генотипом BB.
- Таким образом, у гомозиготных людей генотип цвета глаз будет BB или bb.
- Все остальные цвета глаз от синего до зеленого и серого являются рецессивными признаками, которые проявляются только тогда, когда генотип гомозиготно-рецессивный, bb.
- Если генотип гомозиготный, гены цвета глаз в локусах двух хромосом должны быть идентичными.
- Фенотип индивида, то есть цвет глаз у индивида, зависит от результирующего генотипа этого персонажа.
- Если мы рассматриваем генотип для типа волос как H для доминантного признака и h для рецессивного признака, генотип для вьющихся волос будет HH или Hh.
- Этот генотип кодирует белок, который делает волосы вьющимися.
- Если генотип HH, у человека будут вьющиеся волосы, но при наличии гетерозиготного генотипа волосы будут волнистыми, то есть между вьющимися и прямыми волосами.
- Чтобы волосы были прямыми, генотип должен быть hh, поскольку прямые волосы являются рецессивным признаком.
- Полное преобладание вьющихся волос наблюдается у лиц с гетерозиготным состоянием.
- 2% — https://biodifferences.com/difference-between-phenotype-and-genotype.html
- 1% — https://www.britannica.com/science/phenotype
- 1% — https://www.biologyonline.com/dictionary/phenotype
- 1% — https://en.wikipedia.org/wiki/Phenotype
- 1% — https: //en.wikipedia.org / wiki / Генотип
- 1% — https://byjus.com/biology/genotype-and-phenotype-difference/
- <1% - https://www.spectrumnews.org/news/genetics-identical-twins-not-so-similar/
- <1% - https://www.slideshare.net/SRMegh/silent-mutation-2
- <1% - https://www.researchgate.net/publication/329214501_Biparental_Inheritance_of_Mitochondrial_DNA_in_Humans
- <1% - https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3563493/
- <1% - https: // www.ncbi.nlm.nih.gov/books/NBK7908/
- <1% - https://quizlet.com/361810829/chapter-22-and-23-darwin-flash-cards/
- <1% - https://quizlet.com/35
- 78/genetics-ch-1-flash-cards/
- <1% - https://quizlet.com/25549586/genetics-flash-cards/
- <1% - https://quizlet.com/114950720/ch-15-ch-16-study-guide-flash-cards/
- <1% - https://curlybetty.wordpress.com/2011/05/10/curly-hair-genetics-heredity/
- <1% - https: // biologywise.com / гомозиготный-против-гетерозиготный
границ | Дифференциальный взгляд на отношения генотип – фенотип
Введение
Иногда кажется, что мы забываем, что исходный вопрос в генетике заключался не в том, что делает белок, а в том, «что делает собаку собакой, а человека человеком».
(Благородный, 2006)
Один из фундаментальных вопросов биологии — понять, что отличает людей, популяции и виды друг от друга.Концепция фенотипа , который соответствует наблюдаемым признакам индивидуума, была придумана в противовес генотипу , унаследованному материалу, передаваемому гаметами. С момента первоначального предположения, что генотипы и фенотипы образуют два принципиально разных уровня биологической абстракции (Johannsen, 1911), проблема заключалась в том, чтобы понять, как они соединяются друг с другом, как генотипы отображаются на фенотипах. За последние 15 лет более 1000 примеров изменений последовательности ДНК были связаны с естественными не вредными фенотипическими различиями между людьми или видами у эукариот (Martin and Orgogozo, 2013b).У человека каталог OMIM ® (онлайн-менделевское наследование у человека, http://omim.org/), в котором собраны генетические детерминанты связанных с заболеванием фенотипов, насчитывает более 4300 статей и в общей сложности 2493 опубликованных исследования геномных ассоциаций. (GWAS) обнаружили множество участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в скоплении данных.Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые хотят попытаться оспорить свое нынешнее понимание фенотипов. Выделим одну полезную точку зрения, дифференциальную точку зрения. Затем мы показываем, что эта простая структура остается проницательной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
Гены как факторы различия
Мутации, выделенные из лабораторных штаммов, сыграли важную роль в понимании карты GP.По классической схеме мутация сравнивается с эталоном дикого типа, и ее фенотипические эффекты используются для определения функции гена. Эта структура часто приводит к семантическому сокращению: из генетического изменения, вызывающего вариацию в фенотипе, часто удобно ассимилировать соответствующий ген в качестве причинной детерминанты признака (Keller, 2010; Рисунок 1A). Часто можно встретить заголовки, выражающие эти упрощения, трубящие перед широкой аудиторией об открытии гена «долголетия» или «благополучия», которые жертвуют научной точностью ради психологического воздействия.Исходя из этого, следует ли ген, чья мутация является летальной, называть «геном жизни»? Эти чрезмерно упрощенные формулировки на самом деле означают, что вариация в данном гене вызывает вариацию в данном фенотипе (Dawkins, 1982; Schwartz, 2000; Waters, 2007). Фактически, сам по себе ген не может ни вызвать наблюдаемого фенотипического признака, ни быть необходимым и достаточным для появления наблюдаемых признаков. Гены нуждаются в клеточной среде, комбинированном действии множества других генов, а также в определенных физико-химических условиях, чтобы оказывать заметное влияние на организмы (рис. 1B).Например, каштановая пигментация волос у одного человека является продуктом не только генов, кодирующих ферменты, синтезирующие пигмент, но и присутствия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, тщетен, чтобы полностью ответить на широкий вопрос о том, что делает волосы коричневыми, что вызывает конкретную биологическую структуру или процесс в целом.Тем не менее, генетический редукционизм может быть совершенно подходящим для идентификации генетических локусов, изменение которых вызывает фенотипические различия (рис. 1C). Разница в цвете волос между двумя людьми в некоторых случаях может быть связана с их генетическими различиями. Отметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Разница в цвете волос также может быть вызвана негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окрашивание волос, или сочетанием генетических и негенетических различий.
РИСУНОК 1. Схематическое изображение каналов GP. (A) Традиционное представление в классическом генетическом редукционизме. (B) Интегративный взгляд на биологию развития. (C) Схема экспериментального подхода в генетике. (D) Схема экспериментального подхода в эволюционной генетике. (E) Один пример таксономически устойчивой взаимосвязи GP: SLC45A2 и внутривидовые различия в пигментации у тигров и кур.Связи GP обозначены пунктирными линиями на панелях (D – E) .
Пока современная генетика находилась в зачаточном состоянии, Альфред Стертевант сформулировал вопрос о карте GP простыми словами: «Одна из центральных проблем биологии — это проблема дифференциации — как яйцо превращается в сложный многоклеточный организм? Это, конечно, традиционная главная проблема эмбриологии; но это также проявляется в генетике в форме вопроса: «Как гены производят свои эффекты?» (Стертевант, 1932).Долгое время некоторые генетики могли думать, что они анализируют морфогенетические механизмы, лежащие в основе формирования фенотипических признаков, в то время как их экспериментальный подход на самом деле обнаруживал гены, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. Д.) Приводят к фенотипические различия (сравните фиг. 1A с фиг. 1C). Фактически, предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и бледного пигмента феомеланина), либо как неявную ссылку на другие возможности. (он коричневый, а не другого цвета).Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наш разум и наш язык часто путают объекты, вариации которых рассматриваются, с самими вариациями (Keller, 2010), и важно напомнить, что в генетике интересующие объекты (например, данный генотип, аллель или фенотип) заслуживают определения относительно другого эталонного состояния.
Таким образом, классический генетический редукционистский подход по своей сути неспособен выяснить все факторы, ответственные за наблюдаемые характеристики в живом мире (Stotz, 2012), но является мощным и актуальным методом анализа генетических рычагов наследственной фенотипической изменчивости.Сосредоточение внимания на фенотипических вариациях между людьми, а не на абсолютных признаках, присутствующих в отдельных организмах, является ключом к лучшему пониманию генетических причин фенотипического разнообразия.
Связь GP находится между двумя уровнями вариации
Размышление о различиях делает очевидной абстрактную сущность, которая включает в себя как генетический, так и фенотипический уровни. Этот объект состоит из вариации в генетическом локусе (два аллеля), связанных с ним фенотипических изменений (два различных фенотипических состояния) и их взаимоотношений (рис. 1D).Мы втроем называем совокупность этих элементов «гефе», но здесь мы просто называем это «отношениями генотип-фенотип» (отношения GP). Мы покажем, что отношения GP — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия.
Генетическая часть взаимоотношений с терапевтом
В современных базах данных аннотаций генома ген обычно определяется как участок нуклеиновых кислот, который транскрибируется и кодирует РНК или полипептид с известной или предполагаемой функцией (Gerstein et al., 2007). Генетический локус, лежащий в основе фенотипических различий, не обязательно является геном в строгом смысле слова; он может охватывать конкретную пару оснований, кодирующую область, регуляторную область цис- или распространяться на весь ген с его регулирующими областями цис- или даже на кластер генов (Таблица 1). Как ранее отмечалось другими (Falk, 1984; Gilbert, 2000; Stern, 2000; Moss, 2003; Griffiths and Stotz, 2013), концепция гена в биологии развития и в современных базах данных аннотаций генома отличается от концепции гена в эволюционная биология.Здесь упор делается не на сам ген, как это определено в базах данных геномов, а на индивидуальное функциональное разделение генома на локусы, создающие различия. Генотипическая часть родства GP может принимать форму различных аллелей: отдельные кодоны, кодирующие разные аминокислоты, вставки / делеции в кодирующей последовательности белка, расходящиеся версии определенного регуляторного элемента cis-, наличие / отсутствие вставок транспозонов. , количество копий гена в генном кластере, подверженном структурным изменениям и т. д.В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, включая так называемую мусорную ДНК (Graur et al., 2015), присутствие которой не влияет на наблюдаемые характеристики организма, кроме репликации и, возможно, транскрибирования. Есть также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и которые больше не связаны с фенотипической изменчивостью.Например, генетические вариации в кодирующих областях связывания гистоновой ДНК могли быть важны на ранней стадии эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, внутри генома есть нуклеотидные участки, которые абсолютно необходимы для жизни, но сами не содержат жизнеспособных фенотипически значимых вариаций.
ТАБЛИЦА 1. Несколько примеров отношений GP.
Фенотипическая часть семейных отношений
Фенотипический аналог отношения GP относится к разновидности вариации (цвет волос, уровень устойчивости к токсинам и т. Д.)), а не до состояния (светлые волосы, дегустатор фенилтиокарбамида и др .; Таблица 1).
Фенотип, связанный с генетическим изменением, не обязательно ограничивается организмом, который несет генетическую мутацию. Например, у улитки Lymnaea peregra различие между левыми и правыми спиралями раковин определяется одним генетическим локусом с материнским эффектом: генотип матери, но не самой особи, отвечает за направление наматывание оболочки (Boycott et al., 1931). В других случаях причинное генетическое изменение лежит в пределах бактерий-симбионтов: термостойкость тли может варьироваться от человека к человеку из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты также могут проявляться на более высоком уровне, чем организм, у которого есть генетическое изменение (Dawkins, 1982), одним из примеров является социальная организация колонии муравьев (Wang et al., 2013).
Дифференциальная часть отношений с GP
Как определено выше, отношение GP включает генетическое различие и фенотипическое различие.Взаимосвязь различия как на генетическом, так и на фенотипическом уровне довольно абстрактна и может соответствовать трем отчетливым различиям в живом мире: (# 1) различие между двумя репродуктивно изолированными таксонами (живыми или вымершими), (# 2) различие, сегрегирующее внутри популяции, и (# 3) различие, впервые появившееся во время эволюции, между организмом, несущим предковый аллель / признак, и его прямым потомком, который развил новый аллель / признак. Следует отметить, что изменение фенотипа не всегда следует сразу же за появлением новой вызывающей мутации, но может появиться позже из единственного набора аллелей, сегрегированных в популяции.Например, новый фенотип редуцированных пластин брони появился в популяции пресноводной колюшки, когда рецессивный аллель EDA , уже присутствующий на скрытых уровнях, оказался в гомозиготном состоянии у одного человека (Colosimo et al., 2005; Jones et al., 2012). Главный концептуальный прогресс, сделанный Чарльзом Дарвином, состоял в том, чтобы связать вариации между особями внутри скрещивающейся группы (различие № 2) с вариациями между таксономическими группами в пространстве и времени (различие № 1; Lewontin, 1974a).
Отметим также, что определенные фенотипические изменения могут проявляться на уровне всего организма, когда «причинная» мутация сопровождается дополнительными соматическими мутациями, которые весьма вероятны. Например, у женщин, несущих аллель дикого типа и мутантный аллель BRCA1 , клетки могут продуцировать белки BRCA1 дикого типа, поскольку они несут одну копию аллеля BRCA1 дикого типа. Тем не менее, эти женщины имеют до 80% риска развития рака груди или яичников к 70 годам по сравнению с женщинами, несущими две копии BRCA1 дикого типа, из-за появления дополнительных вредных мутаций в гене BRCA1 дикого типа. аллель в их соматических клетках молочной железы (Narod and Foulkes, 2004).
Важно отметить, что разница GP всегда определяется относительно представляющей интерес популяции или таксона (Sober, 1988). В менее развитых с медицинской точки зрения странах люди, несущие две дефектные копии гена фенилаланингидроксилазы, имеют серьезные проблемы со здоровьем, включая судороги и умственную отсталость. Напротив, в большинстве развитых стран у таких людей диагностируется при рождении, и они имеют нормальную продолжительность жизни с нормальным умственным развитием благодаря диете с ограничением фенилаланина (Armstrong and Tyler, 1955).Таким образом, связь между GP, включающая дефектную мутацию фенилаланингидроксилазы, зависит от контекста: мутация связана с проблемами со здоровьем в менее развитых с медицинской точки зрения странах, но не в других странах. Этот пример показывает, что причинно-следственная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика для случая фенилаланингидроксилазы) в рамках предположения ceteris paribus о «при прочих равных условиях».”
Таким образом, отношения GP лучше всего рассматривать как отношения между двумя вариациями, одна на генотипическом уровне, а другая на фенотипическом уровне. Человеческий разум может разработать концепции возрастающей абстракции: концепции вещей (например, клетки), концепции изменения (например, эволюции) и концепции отношений (например, гомология; Cassirer, 1910; Simondon, 1968). Здесь концепция отношения GP устанавливает связь между двумя изменениями (генетическими и фенотипическими). В следующих параграфах мы покажем, что, по сравнению с использованием интуитивных представлений о вещах, этот обходной путь через усиление абстракции может оказаться более эффективным для лучшего понимания фенотипического разнообразия.
Несколько текущих представлений о связи между генотипом и фенотипом неявно отклоняют дифференциальную точку зрения
Выше мы утверждали, что всегда следует помнить о дифференциальной точке зрения, думая о связи между генотипами и фенотипами. GWAS, которые представляют собой наиболее популярный метод обнаружения геномных локусов, связанных со сложными признаками в популяциях, основаны на анализе различий (Visscher et al., 2012). Тем не менее, в текущих исследованиях дифференциальная точка зрения иногда неявно отвергается.Когда наблюдаются множественные факторы, влияющие на фенотипические признаки (рис. 1B), дифференциальная точка зрения считается слишком упрощенной, и исследователи часто предпочитают снова сосредоточиться на фенотипах отдельных людей, не связывая их явно с фенотипической ссылкой.
В большинстве современных статей проблема связи генотипа с фенотипом формулируется в терминах карт генотипа и фенотипа. Первая карта GP была представлена Ричардом Левонтином в его книге «Генетическая основа эволюционных изменений» (Lewontin, 1974a; рис. 2A).Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разделен на четыре этапа: (1) средний фенотип является производным от развития отдельных генотипов в различных средах; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, у которых будет потомство; (3) личность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутации и рекомбинация, изменяют положение в G-пространстве.
В другом распространенном графическом представлении (рис. 2B) точка в пространстве G и соответствующая ей точка в пространстве P соответствуют генотипу и фенотипу отдельного человека (Fontana, 2002; Landry and Rifkin, 2012). При таком представлении абстрактный объект, который мы определили выше как отношение GP, будет соответствовать «движению» в пространстве генотипа, связанному с «движением» в пространстве фенотипа (или, лучше, сумме нескольких «ходов» в генотипе, и пространства фенотипов, потому что несколько различных геномов могут нести два альтернативных аллеля данного отношения GP).В третьем представлении, выдвинутом Вагнером (1996; рис. 2C), отдельные гены связаны с отдельными признаками.
Хотя эти три графических представления карт GP могут облегчить наше понимание определенных аспектов биологии, во всех из них связь GP и дифференциальная точка зрения понять непросто. Вызывает недоумение то, что первым, кто нарисовал такую карту GP, был Ричард Левонтин, красноречивый сторонник дифференциальной точки зрения (см., Например, его предисловие к Oyama, 2000, шедевр убеждения).Поскольку эти графики сосредоточены на отдельных, а не на различных объектах, мы считаем, что эти три представления неявно побуждают нас вернуться к более интуитивному представлению об одном генотипе, связанном с одним фенотипом. Упущение из виду дифференциальной точки зрения может также происходить с точки зрения молекулярной биологии, где белки рассматриваются как имеющие причинные эффекты сами по себе, такие как фосфорилирование субстрата или связывание с последовательностью ДНК. Из-за двух запутанных определений гена, кодирующего белок или вызывающего фенотипические изменения (Griffiths and Stotz, 2013), легко перейти от дифференциальной точки зрения к недифференциальной точке зрения на отношения GP.
Таким образом, многие современные ментальные представления о связи между генотипом и фенотипом неявно отвергают дифференциальную точку зрения. Теперь мы покажем, что дифференциальная точка зрения совместима с тем фактом, что на фенотипические черты влияет сложная комбинация множества факторов, и что мы можем найти соответствующее схематическое представление отношений GP.
Проблема плейотропии
Разложение организма на элементарные единицы, такие как анатомические структуры, сыграло важную роль во многих биологических дисциплинах, таких как физиология, палеонтология и эволюция.Однако проблема состоит в том, чтобы определить декомпозицию на символы, наиболее подходящую для интересующего вопроса. Для вопросов, связанных с взаимоотношениями между органами разных людей или видов (например, гомология), может быть целесообразно сохранить традиционную декомпозицию на анатомические структуры (Wagner, 2014). Ричард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют квазинезависимые единицы эволюционных изменений (Левонтин, 1978; Вагнер, 2000).Их определение касается абсолютных черт, наблюдаемых у отдельных организмов (например, формы крыла или количества пальцев у особи), и, таким образом, далеко от дифференциальной точки зрения. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые атрибуты организма на множество элементарных вариаций GP, которые накапливались в течение нескольких поколений, начиная с начального состояния. Мы настаиваем на том, что с этой точки зрения персонажи — это не конкретные объекты (например, кожа), а абстрактные сущности, определяемые существованием различий между двумя возможными наблюдаемыми состояниями (например, цветом кожи).В качестве аналогии можно представить два способа изготовления поношенной кожаной обуви определенной формы. Можно либо собрать разные атомы в одну и ту же организацию, либо можно купить обувь в магазине и затем подвергнуть ее воздействию ряда механических сил. Мы естественно склонны сравнивать организмы с машинами и мыслить категориями частей, которые необходимо собрать, чтобы образовать функциональное целое. Однако безудержная метафора дизайнера или изготовителя неадекватна для понимания происхождения современных организмов (Coen, 2012).Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые произошли последовательно в течение эволюционного времени, а не во время развития. Исходное состояние является гипотетическим предком исследуемого организма.
Определенные мутации (квалифицируемые как плейотропные) наблюдаются одновременно в нескольких органах, в то время как другие изменяются только по одному (Paaby and Rockman, 2013; Zhang and Wagner, 2013). Для плейотропных мутаций мы считаем, что отношения GP должны включать все фенотипические изменения (в разных органах, на разных стадиях и т. Д.)) связано с генетическим различием. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменениями плотности потовых и молочных желез в азиатских популяциях (Kamberov et al., 2013). В таких случаях отношения GP являются взаимно-множественными. Рассмотрение кожи и глаза как независимых анатомических модулей человеческого тела может показаться подходящим для многих эволюционных изменений, но это несколько неадекватно в тех случаях, когда эти два органа развили новую черту пигментации одновременно через единственную мутацию в гене SLC45A2 (Лю и другие., 2013). Рассуждения в терминах отношений GP снимают проблему нахождения релевантной декомпозиции на элементарные анатомические структуры. Сами элементарные отношения GP выглядят как адекватные полунезависимые модули, комбинация которых может объяснить наблюдаемые характеристики организма.
Проблема непрерывных сложных черт
Согласно дифференциальной концепции отношений GP, одним из важнейших моментов является разложение наблюдаемых признаков на серию полунезависимых фенотипических вариаций, то есть выявление элементарных изменений, произошедших в ходе эволюции.Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разными формами листьев дает потомство, которое демонстрирует составной диапазон промежуточных форм листьев. Анализ главных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и которые, по-видимому, вызваны разными геномными регионами (Langlade et al., 2005). Это в какой-то мере говорит о том, что «сумма затемняет части.«То, что мы традиционно считаем сложными, можно сделать из более простых, более поддающихся генетическому анализу. Другой яркий пример — пигментация живота в группе Drosophila dunni . Взятые как единственная переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение паттернов взрослых на анатомические субъединицы нарушает дискретный генетический контроль для каждого суб-признака (Hollocher et al., 2000). Более известный случай — это эволюция цвета тела у пляжных мышей.Разница в цвете между светлыми пляжными мышами и темными мышами может быть разложена на различные фенотипы (спинной оттенок, дорсальная яркость, ширина полосы на хвосте и дорсовентральная граница), которые все связаны с различными мутациями в гене Agouti ( Linnen et al., 2013; рисунок 3). Кажется, что каждый генетический локус Agouti предназначен для спецификации пигментации в данной части тела. Вместе они образуют группу тесно связанных локусов, которые связаны с изменениями пигментации шерсти.
РИСУНОК 3. Эволюция светлых пляжных мышей вызвана несколькими мутациями с отчетливыми эффектами пигментации в локусе Agouti . Темный и светлый фенотипы можно разделить на четыре фенотипических признака, которые связаны с различными однонуклеотидными полиморфизмами (SNP, цветные точки), локализованными в гене Agouti . Показаны только SNP с предполагаемым коэффициентом отбора легкого аллеля выше 0,1. Кодирующие экзоны представлены темными прямоугольниками, а непереведенные экзоны — белыми.По материалам Linnen et al. (2013).
Хотя сложные черты не всегда могут быть сведены к набору простых взаимосвязей GP, вполне возможно, что такие черты, как рост взрослого человека, наиболее символический количественный признак, который, по прогнозам, состоит из множества генетических эффектов небольшого размера (Fisher, 1930), могут также можно разложить на элементарные вариации, каждая из которых объясняет более дискретные суб-черты. В то время как некоторые детерминанты роста человека, такие как LIN28B , были связаны с ростом взрослого человека в разном возрасте, другие гены достигли статистической значимости только в исследованиях, посвященных конкретным стадиям, с упором на рост плода и скорость роста в период полового созревания (Lettre, 2011).Другими словами, эти данные предполагают, что рост человека может быть составной чертой, которая модулируется несколькими отношениями GP, каждая из которых действует на разных фазах развития.
Проблема эпистаза и GxE
Взаимодействие «ген за окружающей средой» (GxE) происходит, когда фенотипический эффект данного генетического изменения зависит от параметров окружающей среды. Сходным образом эпистаз или взаимодействие GxG происходит, когда фенотипический эффект данного генетического изменения зависит от аллельного состояния по крайней мере одного другого локуса (Phillips, 2008; Hansen, 2013).Появляется все больше свидетельств того, что взаимодействия GxG и GxE имеют фундаментальное значение для понимания эволюции и наследования сложных признаков (Gilbert and Epel, 2009; Hansen, 2013). Мы предполагаем, что оба явления могут быть интегрированы в базовую дифференциальную структуру GP, где взаимодействия GxG и GxE создают слой зависимости от контекста и приводят к различиям, заключенным в различиях.
Разница в цветовой пигментации между темными и светлыми пляжными мышами, упомянутая ранее (рис.3), связана не только с мутациями в Agouti , но и с кодирующей мутацией в гене MC1R , который уменьшает пигментацию (Steiner et al., 2007; Рисунок 4B). Эффект мутации MC1R виден только в присутствии связанного со светом производного гаплотипа Agouti . Здесь локус Mc1R считается эпистатически взаимодействующим с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два возможных фенотипических различий (изменение пигментации шерсти или отсутствие изменений вообще). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь в локусе Agouti ).Таким образом, дифференциальная точка зрения остается относительно простой для двухлокусных взаимодействий: контекстная зависимость фенотипа транслируется в выбор между двумя возможными фенотипическими различиями. Мы предлагаем, чтобы отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, включали все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах. Среди всех возможных фенотипических вариаций наблюдаемые фенотипические различия определяются другими генетическими локусами.В общем, взаимодействия GxG включают несколько сайтов, которые рассредоточены по геному (Bloom et al., 2013).
РИСУНОК 4. Взаимодействия между генами (GxE) и GxG. (A) Кодирующая мутация npr-1 влияет на агрегацию нематод при 21% -ном уровне кислорода, но не при 10% (Andersen et al., 2014). (B) Кодирующая мутация Mc1R влияет на пигментацию тела мыши в присутствии доминантных светлых аллелей Agouti , но не на гомозиготном фоне Agouti по рецессивному темному аллелю (Steiner et al., 2007).
Примером взаимодействия GxE (см. Также рисунок 4A) является встречающийся в природе аллель потери функции brx у растений Arabidopsis , который связан с ускоренным ростом и повышенной приспособленностью в кислых почвах, а также с сильно уменьшенными корнями. рост по сравнению с диким типом в нормальных почвах (Gujas et al., 2012). Взаимодействия GxE обычно анализируются в форме нормы реакции , которая представляет все наблюдаемые признаки единственного генотипа в различных средах (Johannsen, 1911; Sarkar, 1999).В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Связанное с этим фенотипическое изменение, таким образом, представляет собой разницу между двумя нормами реакции. Примером из учебника является правило изменения температуры и размера в C. elegans . Как и большинство других животных, нематоды C. elegans растут крупнее при низкой температуре, но являются лабораторным штаммом дикого типа C.elegans , происходящего с Гавайев, не показывает изменений в размере тела при различных температурах. Аминокислотная замена в кальций-связывающем белке ответственна за снижение способности гавайского штамма расти при низкой температуре (Kammenga et al., 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанная с ней взаимосвязь GP включает разницу между этими двумя наклонами.
Диапазон фенотипических вариаций, воплощенных в отношениях GP, подверженных взаимодействиям GxG и GxE, может быть довольно огромным, особенно в случаях, когда несколько тканей затронуты одной и той же мутацией, и когда на фенотипические вариации каждой ткани влияют другие геномные локусы и условия окружающей среды.Фактически, фенотипические эффекты мутации всегда зависят от других фрагментов ДНК из того же генома, так что любые отношения GP могут рассматриваться как переживание эпистаза. Другими словами, генетический локус, влияющий на фенотип, никогда не действует независимо от других последовательностей ДНК. Например, данный аллель опсина будет приводить к определенным свойствам цветового зрения только в том случае, если глаз сформирован и если этот глаз получает свет во время своего развития, что позволяет формировать эффективные нейронные цепи зрения. Чтобы дифференцированный взгляд был податливым, мы советуем не рассматривать все возможные генетические фоны и условия окружающей среды, а ограничить возможности потенциальной средой и сегрегацией аллелей, относящихся к интересующей популяции (Sober, 1988).
Таким образом, при наличии эпистаза или взаимодействий GxE генетическое изменение связано не с одним фенотипическим различием, а с множеством возможных фенотипических различий, одно из которых будет достигнуто в зависимости от окружающей среды и генетического фона. Зависимость от контекста может быть схематически представлена как различия GP, встроенные в различия других генотипов и среды.
Дифференциальный взгляд на генетические и экологические эффекты на фенотипы
Как подчеркнуто несколькими авторами (прежде всего Waddington, 1957; Oyama, 2000; Keller, 2010), гены и окружающая среда действуют совместно на фенотип, и в большинстве случаев невозможно отделить эффект одного от другого.Здесь мы показываем, что рассуждения о различиях помогают прояснить сравнение между генетическим и средовым влиянием на фенотипы. Однако мы выявляем определенные случаи, когда сравнение остается затруднительным.
По аналогии с отношениями GP, мы можем определить отношения между средой и фенотипом как средовую вариацию (две среды), связанные с ней фенотипические изменения (различные фенотипические состояния) и их отношения. Например, у многих видов черепах изменение температуры во время развития яйца связано с различием пола самец / самка (Рисунок 5A), и по крайней мере шесть переходов от экологического к генетическому определению пола (Рисунок 5B) произошли в филогении черепах (Pokorná и Кратохвил, 2009).В этом случае можно сравнить экологические и генетические эффекты: половые хромосомы и температура имеют одинаковый фенотипический эффект на черепах. Такие наблюдения привели Вест-Эберхарда (2003, 2005) к предложению гипотезы «гены как последователи», которая предполагает, что новые фенотипические состояния с большей вероятностью возникают сначала из-за изменения окружающей среды, чем из-за генетической мутации, и что мутации происходят только позже, при изменении порога проявления новой черты. Вест-Эберхард (2003, 2005) экстраполировал от различий, сегрегированных внутри популяций (различие № 2), до различий, которые возникли во времени во время эволюции популяции (различие № 1).
РИСУНОК 5. Отношения между средой и фенотипом и отношения GP для определения пола у черепах. (A) У некоторых видов температура во время эмбрионального развития определяет пол взрослой особи. (B) У других пол определяется половыми хромосомами.
Независимая эволюция направленной лево-правой асимметрии от симметричных предков во множественных ветвях предоставила главный аргумент в пользу гипотезы «ген как последователь» (Palmer, 2004).Согласно этой схеме, направленная асимметрия, при которой все индивиды односторонние, часто эволюционировала из состояния «случайной асимметрии», где направленность будет зависеть от факторов окружающей среды и, таким образом, будет варьироваться между генетически идентичными индивидами. Например, самая сильная клешня омара будет развиваться в зависимости от использования и имеет априори равную вероятность развития левой или правой стороны. Мы можем видеть, как здесь применяется формула «гены как последователи»: среда вызывает асимметрию, и позже в эволюции некоторые генетические эффекты могут изменить ее направленность в ту или иную сторону.Но в то время как асимметрия «возникает до того, как существует генетическая изменчивость, которая ее контролирует», дифференциальная точка зрения проясняет, что генетическое влияние на направленность несопоставимо с влиянием окружающей среды, которое запускает асимметрию. Генетическое изменение вызывает переключение между окончательным 100% односторонним состоянием и начальным состоянием, при котором 50% случаев являются правосторонними и 50% левосторонними. Напротив, два альтернативных фенотипических состояния, возникающих в результате изменения окружающей среды, считаются 100% правым и 100% левым.Этот пример показывает, что для точности важно явно указать различия, которые учитываются в отношениях GP.
Дифференциальный взгляд обеспечивает теоретическую основу, которая может помочь в разработке экспериментов по исследованию правильных переменных: можно сравнивать разные генотипы в фиксированной среде (классические отношения GP), сравнивать реакцию фиксированного генотипа на две разные среды (фенотипическая пластичность) , или сравнить чувствительность двух разных генотипов к двум различным средам (при этом фенотипическая вариация становится различием в различии ; см., например, Engelman et al., 2009; Томас, 2010).
Были разработаны различные количественные методы, позволяющие отделить генетические эффекты от воздействия окружающей среды и количественно оценить взаимодействия GxE (Lynch and Walsh, 1998). Тем не менее, в определенных ситуациях может оказаться невозможным биологически значимым образом отделить генетические эффекты от воздействия окружающей среды, даже если рассуждать с точки зрения различий (Lewontin, 1974b). Популяции жука Calathus melanocephalus включают две морфы: длиннокрылые и короткокрылые (Schwander, Leimar, 2011).Длиннокрылые морфы развиваются только от гомозиготных особей по рецессивному аллелю, сегрегационному в популяции, и только при хороших условиях питания. В этом случае генетические эффекты и эффекты окружающей среды переплетаются (Рисунки 6A, B). В теоретическом случае популяции, состоящей только из короткокрылых гетерозиготных животных, выращенных в условиях голода, и длиннокрылых, и гены, и окружающая среда ответственны за различие крыльев между особями, и невозможно оценить долю окружающей среды и генетические эффекты, потому что гены и окружающая среда действуют на разных уровнях по сложной причинной связи между генотипами и фенотипами.
РИСУНОК 6. Взаимосвязь «окружающая среда-фенотип» и перспективы взаимосвязи GP для полиморфизма длины крыла у жука Calathus melanocephalus . (A) С точки зрения взаимосвязи окружающая среда-фенотип, изменение условий питания связано с изменением размера крыла, но только с гомозиготным фоном для рецессивного аллеля (-1 ) локуса размера крыла. (B) С точки зрения взаимоотношений GP, генетическое изменение в локусе размера крыла связано с изменением размера крыла, но только в хороших условиях питания.
Другой случай, который ставит под сомнение классическое различие между окружающей средой и генетикой, — это когда добавление определенных симбиотических бактерий изменяет фенотип хозяина. Мыши, которых кормили штаммом бактерий Lactobacillus , демонстрируют сниженное поведение, связанное с тревогой, по сравнению с контрольными мышами, которых кормили бульоном без бактерий (Bravo et al., 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного кишечного симбионта. Причиной фенотипических различий является не простое изменение последовательности ДНК или простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между наличием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — которые содержат ДНК, мутации которой также могут изменять фенотип хозяина.
В заключение, рассуждения о различиях могут помочь прояснить сравнение между генетическим воздействием и влиянием окружающей среды на фенотипы. Однако проблемы здесь очень простые. Поскольку гены и окружающая среда действуют на разных уровнях сложной причинной связи между генотипами и фенотипами, в некоторых случаях невозможно разделить обе причины.
Разъяснение терминологии «прибыль / убыток» и «Разрешительный» / Инструкция
Фенотипические различия, по-видимому, подпадают под две основные категории: либо наличие / отсутствие чего-либо (например, волос на теле или способность переваривать молоко), либо сдвиг между двумя альтернативами, которые обе присутствуют (например, два цвета волос).Точно так же на стороне генотипа мутация может соответствовать наличию / отсутствию соответствующей последовательности ДНК или полиморфизму нуклеотидов. Дифференциальная точка зрения делает очевидным, что потеря фенотипа не обязательно связана с потерей генетического материала, и наоборот. Например, эволюционное увеличение темных пигментов, покрывающих всю шерсть животных, часто было связано с потерей гена Mc1R (Gompel and Prud’homme, 2009). Более того, как один из нас ранее отмечал (приложение Stern and Orgogozo, 2008), выигрыш или потеря для фенотипа субъективны.Например, выпадение волос можно рассматривать как прирост голого эпидермиса. Большинство эпидермальных клеток насекомых дифференцируются в одно из этих двух альтернативных состояний, и в обоих состояниях задействованы большие сети регуляции генов. Неясно, какое фенотипическое состояние представляет собой выигрыш или потерю по сравнению с другим. Даже на генотипической стороне определение потерь и прибылей может быть трудным. Вставка мобильного элемента может сбивать ген, тогда как делеция может иногда создавать новый сайт связывания для активатора транскрипции.Фактически, эволюционное усиление экспрессии desatF в D. melanogaster происходит посредством серии из трех делеций, каждая из которых создает мотив гексамера, который необходим для экспрессии desatF (Shirangi et al., 2009).
Точно так же дифференциальный взгляд на воздействие окружающей среды подчеркивает ошибочность различия между разрешающими и поучительными сигналами. Пермиссивный сигнал связан с наличием / отсутствием фенотипа, а инструктивный сигнал — со сдвигом между двумя альтернативами, которые обе присутствуют.Как утверждалось выше, эти различия на фенотипическом уровне нечеткие.
В заключение мы предлагаем использовать терминологию «прибыль / убыток» и поучительную / разрешительную терминологию с осторожностью.
Таксономически устойчивые отношения с GP
Ожидается, что мутация вызовет в некоторой степени воспроизводимые фенотипические вариации в популяции. Такая воспроизводимость фенотипических результатов необходима для генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998).В самом деле, новообразованный аллель, который будет генерировать еще один фенотип каждый раз, когда он окажется в другом организме, не будет подвергнут естественному отбору. Рассуждения с точки зрения вариации, а не рассмотрение аллелей как изолированных сущностей, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор действует непосредственно на аллельную изменчивость, которая последовательно связана с данной фенотипической изменчивостью, которая сама по себе является отношением GP. Таким образом, отношения GP являются базовой единицей эволюционных изменений, на которую действует естественный отбор.
Важным открытием последних 20 лет является то, что вариации в определенных генетических локусах вызывают сравнимые фенотипические вариации не только у разных особей одной популяции, но и у чрезвычайно разных таксонов (Martin and Orgogozo, 2013b). Другими словами, определенные отношения GP устойчивы с таксономической точки зрения и присутствуют в большом диапазоне видов. Это означает, что генетический и экологический фон оставался относительно постоянным или неоднократно появлялся на протяжении эволюции, чтобы позволить генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах.Об этом важном открытии почти 50 лет назад никто не подозревал. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сопоставимую сингулярность на генетическом уровне, включая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 1963). Как однажды предложил Майр (1963) в 1963 году: «Многое из того, что было изучено о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» так же верна как в эволюции, так и в повседневной жизни »(Mayr, 1963).Другими словами, генетические локусы, которые делают человека мужчиной, должны были отличаться от локусов, которые делают собаку или рыбу. Позже, в 80–90-х годах, несколько исследователей предположили совершенно противоположное, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 1978; Perutz, 1983; Stewart et al., 1987; Carroll et al. ., 2005), но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). На сегодняшний день накапливающиеся данные о мутациях, ответственных за естественную изменчивость, ясно показывают, что разнообразие живых организмов имеет общую генетическую основу как минимум по трем пунктам.Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с консервативными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых генетическая основа была хотя бы частично идентифицирована (154 случая из 160), вызваны мутациями в гомологичных генах, и очень немногие (6/160) связаны с новыми генами, которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b).В-третьих, было показано, что множественные случаи сходных фенотипических изменений включают мутации одних и тех же гомологичных генов в независимых клонах (Таблица 1), иногда на больших филогенетических расстояниях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-переносчика SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментированными глазами, кожей , волосы и перья у людей и кур (Xu et al., 2013; Рисунок 1E). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад — моллюска, змеи и иглобрюха — через одну и ту же аминокислотную замену в консервативном гене (Bricelj et al., 2005 ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения сейчас обнаружены для более чем 100 генов у животных и растений (Martin and Orgogozo, 2013b). Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере определенных фенотипических различий существует относительно мало генетических путей. ведут в Рим (Stern, 2013).В настоящее время не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев у одного вида бабочек Heliconius , обеспечивает аналогичные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой, а человека мужчиной, теперь частично объясняется уникальным набором таксономически устойчивых отношений GP, которые обнаруживаются в нескольких ветвях родословной.
Определенные отношения между средой и фенотипом также устойчивы с таксономической точки зрения.Например, у большинства таксонов размер тела зависит от питания; дефицит железа может вызвать анемию, а некоторые токсичные соединения могут быть смертельными. У эктотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, являются ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип более распространенными. Кроме того, является предметом споров, представляют ли таксономически устойчивые отношения GP исключительную и небольшую часть или значительную долю всех отношений GP.В любом случае существование таксономически устойчивых взаимоотношений с терапевтами теперь очевидно и должно быть широко принято исследовательским сообществом.
Некоторые из наиболее ярких учений современной биологии включают открытие того, что живые существа имеют один и тот же генетический материал (ДНК или РНК), один и тот же генетический код (за некоторыми исключениями) и один и тот же базовый клеточный аппарат. Таким образом, совсем не парадоксально, что индивидуальные различия строятся на сходствах, и открытие того, что определенные отношения GP сохраняются в течение длительного времени эволюции, дополняет картину.
Точная предсказательная сила, являющаяся результатом существования таксономически устойчивых взаимосвязей GP, редко встречается в биологии и только начинает использоваться в полной мере. Сохранение связей GP на больших расстояниях теперь полностью оправдывает использование подходов сравнительной генетики для решения прагматических проблем. Например, одомашнивание сельскохозяйственных культур принимало форму аналогичного давления отбора у многих видов, и теперь у нас есть экспериментальные доказательства того, что этот процесс неоднократно включал мутации в одном и том же наборе консервативных генов (Paterson et al., 1995; Мартин и Оргогозо, 2013b). Это наблюдение открывает интересные приложения, поскольку мы можем использовать этот появляющийся массив генетических знаний для помощи в одомашнивании будущих культур или использовать стратегии с использованием маркеров для создания и поддержания биоразнообразия сельскохозяйственных культур (Lenser and Theißen, 2013). Предсказуемость GP уже используется при идентификации штаммов, у которых развилась устойчивость к различным стратегиям борьбы с вредителями, причем в крайних случаях нацелена на устойчивость к противомалярийным препаратам у паразитов Plasmodium (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более важно, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статуса вредителей (Ffrench-Constant et al., 2004; Martin, Orgogozo, 2013б).
Более того, повторяемость генетической основы фенотипической изменчивости предполагает, что клинические исследования также могут выиграть от генетических исследований большого диапазона модельных видов (Robinson and Webber, 2014).Например, естественные вариации толерантности к метотрексату, химиотерапевтическому препарату, были сопоставлены у плодовых мух Drosophila с генами, чьи человеческие ортологи также связаны с реакцией пациентов на это лекарство (Кислухин и др., 2013), таким образом увеличивая использование модельных организмов в качестве моделей болезней.
К классификации фенотипов на основе генов
Одним из оригинальных аспектов определения связи GP с точки зрения индивидуальных отношений GP является то, что он позволяет классифицировать фенотипы в соответствии с их лежащей в основе генетической основой.На первом уровне взаимосвязи GP, затрагивающие разные области одного и того же гена и приводящие к сопоставимым фенотипическим результатам, могут быть сгруппированы вместе. Простые случаи взаимодействия GxG были обнаружены между прочно сцепленными мутациями, как правило, внутри кодирующей последовательности или внутри регуляторного элемента cis-, когда они производят неаддитивный эффект на фенотип. Например, наблюдали, что конкретная мутация в энхансере вызывает различные сдвиги в паттерне экспрессии нижележащего кодирующего гена в зависимости от соседней последовательности ДНК (Frankel et al., 2011; Роджерс и др., 2013). Аналогичным образом, аминокислотные мутации в гене гемоглобина, как было обнаружено, увеличивают или уменьшают сродство к кислороду в зависимости от аллельного состояния других сайтов (Natarajan et al., 2013). В таких случаях интуитивно легко сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же фенотипический признак.
Отсутствие пигментов меланина у животных было связано с мутациями в нескольких генах, включая OCA2 , kit-лиганд или Mc1R (обзор в Gompel and Prud’homme, 2009; Liu et al., 2013). Принимая во внимание, что отсутствие меланина традиционно считается одним из характерных состояний, альбинизмом, независимо от лежащей в основе генетической основы, мы предлагаем здесь отличать OCA2 -ассоциированный альбинизм от Mc1R -ассоциированного альбинизма или от альбинизма, ассоциированного с любым другим геном. Один интерес в разложении вариаций в живом мире на эти множественные элементарные отношения GP состоит в том, что эти элементы затем могут быть сгруппированы вместе в последовательно увеличивающиеся группы.Элементарные фенотипические изменения с участием разных генов, которые являются частью одного и того же генетического пути, также могут быть сгруппированы вместе как сопутствующие компоненты одного и того же механизма модуляции фенотипа. Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9 и рецептору TGF-β BMPR1B, которые все неоднократно были связаны с вариациями функции яичников у людей и домашних пород овец (обзор в Luong et al., 2011).
Другим важным следствием перспективы взаимоотношений GP является то, что очевидно различные фенотипические изменения, вызванные сходными генетическими локусами у разных организмов, могут быть исследованы дополнительно, чтобы выявить то, что могло бы быть общим основным фенотипическим изменением (Deans et al., 2015). Например, личинки мух и черви нематоды имеют разное поведение в поисках пищи, но мутации в одном и том же ортологическом гене ( для / egl-4 ), как было показано, изменяют интенсивность поведения в поисках пищи у обоих организмов (Osborne et al. , 1997; Mery et al., 2007; Hong et al., 2008). Таким образом, вероятно, что базовое изменение поведения, лежащее в основе, казалось бы, отличительных изменений в поиске пищи мухами и нематодами, представляет собой сохраненные отношения GP между нематодами и мухами.Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к заболеванию человека? Можем ли мы считать фенотип мыши похожим на состояние человека, если его генетическая основа иная? Мы и другие предсказываем, что поиск ортологичных фенотипов или «фенологов» (McGary et al., 2010) станет важной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
Заключение
В этой статье мы возвращаем дифференциальную концепцию гена (Schwartz, 2000) в нашу структуру для понимания карты GP. Дифференциальный взгляд на отношения GP помогает прояснить генетическое и экологическое влияние на фенотипы и их связь. Это также открывает новые пути мышления, в частности, в отношении декомпозиции наблюдаемых функций внутри организма и представления карт GP. Кроме того, наличие таксономически устойчивых взаимоотношений GP побуждает беззастенчиво использовать сравнительную генетику для прогнозирования генетической основы фенотипической изменчивости в различных группах организмов, и эта прогностическая сила имеет важный потенциал для трансляционных исследований в агрономии и клинических исследованиях.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы глубоко признательны Мари-Анн Феликс за полезные обсуждения и Томасу Прадо за то, что он обратил наше внимание на несколько важных статей. Мы также благодарим Джузеппе Бальдаччи, Мари-Анн Феликс, Пьера-Анри Гуйона, Александра Пелуффо, Марка Сигала, Дэвида Стерна и рецензентов за их проницательные комментарии к рукописи.Исследование, приведшее к этой статье, получило финансирование от Европейского исследовательского совета в рамках Седьмой рамочной программы Европейского сообщества (FP7 / 2007-2013 грантовое соглашение № 337579) и от Фонда Джона Темплтона (грант № 43903).
Сокращения
GP, генотип – фенотип.
Список литературы
Андерсен, Э. К., Блум, Дж. С., Герке, Дж. П., и Кругляк, Л. (2014). Вариант нейропептидного рецептора npr-1 является основным детерминантом роста и физиологии Caenorhabditis elegans . PLoS Genet. 10: e1004156. DOI: 10.1371 / journal.pgen.1004156
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bloom, J. S., Ehrenreich, I. M., Loo, W. T., Lite, T.-L. В., Кругляк Л. (2013). Выявление источников недостающей наследственности при скрещивании дрожжей. Природа 494, 234–237. DOI: 10.1038 / природа11867
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бойкот, А. Э., Дайвер, К., Гарстанг, С. Л., и Тернер, Ф.М. (1931). Наследование левосторонности у limnæa peregra (Mollusca, Pulmonata). Philos. Пер. R. Soc. Лондон. Сер. В 219, 51–131. DOI: 10.1098 / rstb.1931.0002
CrossRef Полный текст | Google Scholar
Браво, Дж. А., Форсайт, П., Чу, М. В., Эскараваж, Э., Савиньяк, Х. М., Динан, Т. Г. и др. (2011). Проглатывание штамма Lactobacillus регулирует эмоциональное поведение и экспрессию центрального рецептора ГАМК у мыши через блуждающий нерв. Proc.Natl. Акад. Sci. США 108, 16050–16055. DOI: 10.1073 / pnas.1102999108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bricelj, V.M., Connell, L., Konoki, K., Macquarrie, S.P., Scheuer, T., Catterall, W.A., et al. (2005). Мутация натриевых каналов, приводящая к устойчивости к сакситоксину у моллюсков, увеличивает риск PSP. Природа 434, 763–767. DOI: 10.1038 / nature03415
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кэрролл, С.Б., Гренье, Дж., И Уэтерби, С. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных . Молден, Массачусетс: John Wiley & Sons.
Google Scholar
Кассирер, Э. (1910). Substance et Fonction , 2010 Edn. Париж: Nabu Press.
Google Scholar
Коэн, Э. (2012). От клеток к цивилизациям: принципы перемен, формирующих жизнь . Принстон, Нью-Джерси: Издательство Принстонского университета. DOI: 10.1515 / 9781400841653
CrossRef Полный текст | Google Scholar
Колозимо, П.Ф., Хосеманн, К. Э., Балабхадра, С., Вильярреал, Дж. Мл., Диксон, М., Гримвуд, Дж. И др. (2005). Распространенная параллельная эволюция колюшек путем многократной фиксации аллелей эктодисплазина. Наука 307, 1928–1933. DOI: 10.1126 / science.1107239
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Докинз Р. (1982). Расширенный фенотип: долгая досягаемость гена . Оксфорд: Издательство Оксфордского университета.
Google Scholar
Динс, А.Р., Льюис, С. Е., Хуала, Э., Анзалдо, С. С., Эшбернер, М., Балхофф, Дж. П. и др. (2015). Пробираемся сквозь фенотипы. PLoS Biol. 13: e1002033. DOI: 10.1371 / journal.pbio.1002033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Данбар, Х. Э., Уилсон, А. К. С., Фергюсон, Н. Р., Моран, Н. А. (2007). Термостойкость тли определяется точечной мутацией у бактериальных симбионтов. PLoS Biol. 5: e96. DOI: 10.1371 / journal.pbio.0050096
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энгельман, К.D., Baurley, J. W., Chiu, Y.-F., Joubert, B. R., Lewinger, J. P., Maenner, M. J., et al. (2009). Обнаружение взаимодействий между геном и окружающей средой в данных ассоциации по всему геному. Genet. Эпидемиол. 33 (Дополнение 1), S68 – S73. DOI: 10.1002 / gepi.20475
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фельдман, К. Р., Броди, Э. Д. мл., Броди, Э. Д. III, и Пфрендер, М. Э. (2012). Ограничение формирует схождение тетродотоксин-устойчивых натриевых каналов змей. Proc.Natl. Акад. Sci. США 109, 4556–4561. DOI: 10.1073 / pnas.1113468109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Франкель, Н., Эрезилмаз, Д. Ф., МакГрегор, А. П., Ван, С., Пайр, Ф., и Стерн, Д. Л. (2011). Морфологическая эволюция, вызванная множеством малозаметных замен в регуляторной ДНК. Природа 474, 598–603. DOI: 10.1038 / природа10200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Геффени, С.Л., Фудзимото, Э., Броди, Э. Д. III, Броди, Э. Д. мл., И Рубен, П. К. (2005). Эволюционная диверсификация ТТХ-резистентных натриевых каналов во взаимодействии хищник-жертва. Природа 434, 759–763. DOI: 10.1038 / nature03444
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Герштейн, М. Б., Брюс, К., Розовски, Дж. С., Чжэн, Д., Ду, Дж., Корбель, Дж. О. и др. (2007). Что такое ген пост-ENCODE? История и обновленное определение. Genome Res. 17, 669–681.DOI: 10.1101 / gr.6339607
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гилберт, С. Ф. (2000). «Классические гены и гены развития: различные применения генов в эволюционном синтезе», в «Концепция гена в развитии и эволюции» , ред. П. Бертон, Р. Фальк и Х.-Дж. Райнбергер (Кембридж, Нью-Йорк: издательство Кембриджского университета).
Гилберт, С. Ф., и Эпель, Д. (2009). Экологическая биология развития: интеграция эпигенетики, медицины и эволюции .Сандерленд, Массачусетс: Sinauer Associates.
Google Scholar
Гьювсланд, А. Б., Вик, Дж. О., Борода, Д. А., Хантер, П. Дж., И Омхольт, С. В. (2013). Преодоление разрыва между генотипом и фенотипом: что для этого нужно? J. Physiol. 591, 2055–2066. DOI: 10.1113 / jphysiol.2012.248864
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гриффитс П. и Стотц К. (2013). Генетика и философия: введение . Кембридж: Издательство Кембриджского университета.DOI: 10.1017 / CBO9780511744082
CrossRef Полный текст | Google Scholar
Гуджас Б., Алонсо-Бланко К. и Хардтке К. С. (2012). Природные аллели потери функции Arabidopsis brx придают корням адаптацию к кислой почве. Curr. Биол. 22, 1962–1968. DOI: 10.1016 / j.cub.2012.08.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холлохер, Х., Хэтчер, Дж. Л. и Дайресон, Э. Г. (2000). Генетический анализ и анализ развития различий пигментации брюшной полости у разных видов в подгруппе Drosophila dunni . Evol. Int. J. Org. Evol. 54, 2057–2071. DOI: 10.1111 / j.0014-3820.2000.tb01249.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хонг Р. Л., Витте Х. и Соммер Р. Дж. (2008). Природные вариации в привлечении феромонов насекомых Pristionchus pacificus связаны с протеинкиназой EGL-4. Proc. Natl. Акад. Sci. США 105, 7779–7784. DOI: 10.1073 / pnas.0708406105
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джонс, Ф.C., Grabherr, M.G., Chan, Y.F, Russell, P., Mauceli, E., Johnson, J., et al. (2012). Геномные основы адаптивной эволюции трехиглой колюшки. Природа 484, 55–61. DOI: 10.1038 / природа10944
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Камберов Ю.Г., Ван С., Тан Дж., Жербо П., Варк А., Тан Л. и др. (2013). Моделирование недавней эволюции человека на мышах путем экспрессии выбранного варианта EDAR. Ячейка 152, 691–702. DOI: 10.1016 / j.cell.2013.01.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Камменга, Дж. Э., Дорощук, А., Риксен, Дж. А. Г., Хазендонк, Э., Спиридон, Л., Петреску, А.-Дж., и др. (2007). Caenorhabditis elegans дикого типа не подчиняется правилу температуры и размера из-за однонуклеотидного полиморфизма в tra-3. PLoS Genet 3: e34. DOI: 10.1371 / journal.pgen.0030034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Келлер, Э.Ф. (2010). Мираж пространства между природой и питанием . Дарем, Северная Каролина: издательство Duke University Press. DOI: 10.1215 / 9780822392811
CrossRef Полный текст | Google Scholar
Киршнер М. и Герхарт Дж. (1998). Эволюционируемость. Proc. Natl. Акад. Sci. США 95, 8420–8427. DOI: 10.1073 / pnas.95.15.8420
CrossRef Полный текст | Google Scholar
Кислухин, Г., Кинг, Э. Г., Уолтерс, К. Н., Макдональд, С. Дж., И Лонг, А. Д. (2013). Генетическая архитектура токсичности метотрексата сходна у Drosophila melanogaster и человека.3, 1301–1310. DOI: 10.1534 / g3.113.006619
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ландри, К. Р., Рифкин, С. А. (2012). Карты генотипа-фенотипа системной биологии и количественной генетики: отдельные и дополняющие друг друга. Adv. Exp. Med. Биол. 751, 371–398. DOI: 10.1007 / 978-1-4614-3567-9_17
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ланглейд, Н. Б., Фенг, X., Дрансфилд, Т., Копси, Л., Ханна, А. И., Thébaud, C., et al. (2005). Эволюция через генетически контролируемое пространство аллометрии. Proc. Natl. Акад. Sci. США 102, 10221–10226. DOI: 10.1073 / pnas.0504210102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Левонтин Р. (1974a). Генетическая основа эволюционных изменений (Колумбийская биологическая серия) . Колумбия: Издательство Колумбийского университета.
Google Scholar
Левонтин Р. К. (1974b). Дисперсионный анализ и анализ причин. Am. J. Hum. Genet. 26, 400–411.
Google Scholar
Линнен, К. Р., По, Й.-П., Петерсон, Б. К., Барретт, Р. Д. Х., Ларсон, Дж. Г., Дженсен, Дж. Д. и др. (2013). Адаптивная эволюция множества признаков посредством множественных мутаций одного гена. Наука 339, 1312–1316. DOI: 10.1126 / science.1233213
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Луонг, Х. Т. Т., Чаплин, Дж., МакРэй, А. Ф., Медланд, С. Э., Виллемсен, Г., Nyholt, D. R., et al. (2011). Вариация BMPR1B, TGFRB1 и BMPR2 и контроль дизиготного двойникования. Twin Res. Гм. Genet. Выключенный. J. Int. Soc. Twin Stud. 14, 408–416. DOI: 10.1375 / twin.14.5.408
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Линч М. и Уолш Б. (1998). Генетика и анализ количественных признаков . Сандерленд, Массачусетс: Sinauer Associates.
Google Scholar
MacCallum, D. M., Coste, A., Ischer, F., Якобсен, М. Д., Оддс, Ф. К., и Санглард, Д. (2010). Генетическое вскрытие механизмов устойчивости к азолам у Candida albicans и их проверка на мышиной модели диссеминированной инфекции. Антимикробный. Агенты Chemother. 54, 1476–1483. DOI: 10.1128 / AAC.01645–1649
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Манске М., Миотто О., Кампино С., Оберн С., Альмагро-Гарсия Дж., Маслен Г. и др. (2012). Анализ разнообразия Plasmodium falciparum при естественных инфекциях путем глубокого секвенирования. Природа 487, 375–379. DOI: 10.1038 / природа11174
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартин А., Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических горячих точек фенотипической изменчивости. Evol. Int. J. Orgn. Evol. 67, 1235–1250. DOI: 10.1111 / evo.12081
PubMed Аннотация | CrossRef Полный текст | Google Scholar
МакГэри, К. Л., Парк, Т. Дж., Вудс, Дж. О., Ча, Х. Дж., Уоллингфорд, Дж.Б., и Маркотт, Э. М. (2010). Систематическое открытие неочевидных моделей болезней человека через ортологичные фенотипы. Proc. Natl. Акад. Sci. США 107, 6544–6549. DOI: 10.1073 / pnas.00107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мери, Ф., Белей, А. Т., Со, А. К.-К., Соколовски, М. Б., и Кавецки, Т. Дж. (2007). Естественный полиморфизм, влияющий на обучение и память у Drosophila . Proc. Natl. Акад. Sci. США 104, 13051–13055.DOI: 10.1073 / pnas.0702923104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Натараджан К., Иногучи Н., Вебер Р. Э., Фаго А., Морияма Х. и Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340, 1324–1327. DOI: 10.1126 / science.1236862
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ноубл, Д. (2006). Музыка жизни: биология за пределами генома . Оксфорд: Издательство Оксфордского университета.
Google Scholar
Осборн К. А., Робишон А., Берджесс Э., Батланд С., Шоу Р. А., Култхард А. и др. (1997). Полиморфизм естественного поведения за счет цГМФ-зависимой протеинкиназы Drosophila . Наука 277, 834–836. DOI: 10.1126 / science.277.5327.834
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ояма, С. (2000). Онтогенез информации: системы развития и эволюция , 2-е изд.Кембридж, Нью-Йорк: издательство Duke University Press. DOI: 10.1215 / 9780822380665
CrossRef Полный текст | Google Scholar
Патерсон, А. Х., Лин, Ю. Р., Ли, З., Шерц, К. Ф., Добли, Дж. Ф., Пинсон, С. Р. и др. (1995). Конвергентное одомашнивание зерновых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269, 1714–1718. DOI: 10.1126 / science.269.5231.1714
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перуц, М. Ф. (1983).Адаптация видов в белковой молекуле. Мол. Биол. Evol. 1, 1–28.
Google Scholar
Pokorná, M., and Kratochvíl, L. (2009). Филогения механизмов определения пола у плоских рептилий: являются ли половые хромосомы эволюционной ловушкой? Zool. J. Linn. Soc. 156, 168–183. DOI: 10.1111 / j.1096-3642.2008.00481.x
CrossRef Полный текст | Google Scholar
Рокман, М. В. (2012). Программа QTN и аллели, имеющие значение для эволюции: все это золото не блестит. Evol. Int. J. Orgn. Evol. 66, 1–17. DOI: 10.1111 / j.1558-5646.2011.01486.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роджерс, В. А., Саломон, Дж. Р., Тейси, Д. Дж., Камино, Э. М., Дэвис, К. А., Ребеиз, М., и др. (2013). В основе разнообразия пигментации плодовой мушки лежит рекуррентная модификация консервативного цис-регуляторного элемента. PLoS Genet 9: e1003740. DOI: 10.1371 / journal.pgen.1003740
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ромеро-Эррера, А.Э., Леманн, Х., Джойси, К. А., и Фрайдей, А. Э. (1978). Об эволюции миоглобина. Philos. Пер. R. Soc. Лондон. B. Biol. Sci. 283, 61–163. DOI: 10.1098 / rstb.1978.0018
CrossRef Полный текст | Google Scholar
Саркар, С. (1999). От нормы реакции к адаптивной норме: норма реакции, 1909–1960 гг. Biol. Филос. 14, 235–252. DOI: 10.1023 / A: 10066648
CrossRef Полный текст | Google Scholar
Шварц, С.(2000). «Дифференциальная концепция гена: прошлое и настоящее», в The Concept of the Gene in Development and Evolution, eds PJ Beurton, R. Falk, and HJ Rheinberger (Cambridge, NY: Cambridge University Press), 26–39. . DOI: 10.1017 / CBO9780511527296.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ширанги, Т. Р., Дюфур, Х. Д., Уильямс, Т. М., и Кэрролл, С. Б. (2009). Быстрая эволюция экспрессии фермента, продуцирующего половые феромоны, у Drosophila . PLoS Biol. 7: e1000168. DOI: 10.1371 / journal.pbio.1000168
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Симондон, Г. (1968). L’individuation à la Lumière des Notions de Forme et d’Information , 2005th Edn. Гренобль: Миллон.
Google Scholar
Штайнер, К. К., Вебер, Дж. Н. и Хекстра, Х. Э. (2007). Адаптивная изменчивость пляжных мышей вызвана двумя взаимодействующими генами пигментации. PLoS Biol. 5: e219.DOI: 10.1371 / journal.pbio.0050219
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стерн, Д. Л. (2000). Эволюционная биология развития и проблема вариации. Evol. Int. J. Orgn. Evol. 54, 1079–1091. DOI: 10.1111 / j.0014-3820.2000.tb00544.x
CrossRef Полный текст | Google Scholar
Стотц, К. (2012). Убийство в экспрессе развития: кто убил природу / воспитание? Биол . Philos. 27, 919–929.DOI: 10.1007 / s10539-012-9343-1
CrossRef Полный текст | Google Scholar
Стертевант, А. Х. (1932). Использование мозаики в изучении влияния генов на развитие. Proc. Шестой Int. Congr. Genet. Итака Н. Ю. 1, 304–307.
Суппл М., Папа Р., Контрман Б. и Макмиллан У. О. (2014). Геномика адаптивного излучения: понимание континуума видообразования геликониусов. Adv. Exp. Med. Биол . 781, 249–271. DOI: 10.1007 / 978-94-007-7347-9_13
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тауц, Д., и Шмид, К. Дж. (1998). От генов к людям: гены развития и формирование фенотипа. Philos. Пер. R. Soc. Лондон. B. Biol. Sci. 353, 231–240. DOI: 10.1098 / rstb.1998.0205
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Венкатеш, Б., Лу, С.К., Дандона, Н., Си, С.Л., Бреннер, С., и Сунг, Т.В. (2005). Генетические основы устойчивости к тетродотоксинам иглобрюхов. Curr. Биол. 15, 2069–2072. DOI: 10.1016 / j.cub.2005.10.068
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уоддингтон, К. Х. (1957). Стратегия генов, 2014 Edn Reprint. Лондон: Рутледж.
Google Scholar
Вагнер, Г. П. (1996). Гомологи, естественные виды и эволюция модульности. Am. Zool. 36, 36–43. DOI: 10.1093 / icb / 36.1.36
CrossRef Полный текст | Google Scholar
Вагнер, Г. П. (2000). Концепция персонажа в эволюционной биологии .Сан-Диего, Калифорния: Academic Press.
Google Scholar
Вагнер Г. П. (2014). Гомология, гены и эволюционные инновации . Принстон, Нью-Джерси: Издательство Принстонского университета. DOI: 10.1515 / 9781400851461
CrossRef Полный текст | Google Scholar
Wang, J., Wurm, Y., Nipitwattanaphon, M., Riba-Grognuz, O., Huang, Y.-C., Shoemaker, D., et al. (2013). Y-образная социальная хромосома вызывает альтернативную организацию колоний у огненных муравьев. Природа 493, 664–668.DOI: 10.1038 / природа11832
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уотерс, К. К. (2007). Причины, которые имеют значение. J. Philos. 104, 551–579.
Google Scholar
Велтер Д., Макартур Дж., Моралес Дж., Бердетт Т., Холл П., Джанкинс Х. и др. (2014). Каталог NHGRI GWAS, кураторский ресурс ассоциаций SNP-признаков. Nucleic Acids Res. 42, D1001 – D1006. DOI: 10.1093 / nar / gkt1229
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вест-Эберхард, М.Дж. (2003). Пластичность и эволюция развития . Оксфорд: Издательство Оксфордского университета.
Google Scholar
Уилкинс, А. (2014). «Набор генетических инструментов»: история жизни важной метафоры », в Advances in Evolutionary Developmental Biology , ed. Дж.