Физиологические основы восприятия
Процессы, происходящие в органах чувств, нервных волокнах, центральной нервной системе, являются физиологической основой восприятия. В окончаниях нервов, имеющихся в органах чувств под действием раздражителей, возникает нервное возбуждение, передающееся в нервные центры по проводящим путям и в кору головного мозга. Возбуждение поступает в сенсорные зоны коры, представляющие собой центральную проекцию нервных окончаний, имеющихся в органах чувств. Определенная сенсорная информация формируется в зависимости от того, с каким органом связана проекционная зона. Это, в принципе, механизм возникновения ощущений, которые являются отражением окружающей действительности. На уровне предложенной схемы, действительно, формируются ощущения, а это значит, что ощущения могут быть рассмотрены как структурный элемент процесса восприятия.
Процесс восприятия начинается с ощущений. На последующих этапах в процесс формирования целостного образа объекта включаются собственные физиологические механизмы.
Обнаруживается различие в работе этих зон, когда у человека деятельность той или иной зоны нарушена. Нарушение зрительной проекционной зоны приводит к полному лишению зрительных ощущений, и он совсем ничего не видит. Это так называемая центральная слепота.
При сохранности проекционной, но поражении интегративной зоны, человек видит отдельные световые пятна, какие-то контуры, но не понимает что это такое. Аналогичная картина наблюдается и в других модальностях, например, при нарушении слуховых интегративных зон люди перестают понимать речь человека. Это агностические расстройства или агнозии, приводящие к невозможности познания.
Усложняется физиологическая основа восприятия тем, что связана и очень тесно, с двигательной деятельностью, эмоциональными переживаниями, мыслительными процессами.
Можно предположить, что восприятие, как и ощущение, является рефлекторным процессом, поскольку тесно с ним связано.
Рефлекторную основу восприятия раскрыл И.П. Павлов, показав, что условные рефлексы лежат в основе восприятия.
Большое значение в процессе восприятия имеют временные нервные связи. Они обеспечивают возможностью делать гипотезы, необходимые для анализа воспринимаемой ситуации.
Временные нервные связи могут быть двух видов:
- Связи в пределах одного анализатора. Он имеет место при воздействии на организм комплексного раздражителя одной модальности.
- Межанализаторные связи. Эти связи образуются при воздействии комплексного раздражителя, в пределах разных анализаторов.


Избирательность восприятия: апперцепция
В процессе восприятия человек одни аспекты среды выделяет больше, чем другие. Это происходит в соответствии с его целями и намерениями, особенностями деятельности, личностной характеристикой. Совокупность этих факторов определяется как опыт человека. Зависимость процесса восприятия от прошлого опыта получила название апперцепция.
Человек как бы прикладывает свой опыт к восприятию, что дает возможность наполнить чувственный материал образа определенным смыслом.
Сегодня есть большой фактический материал, который иллюстрирует явления, связанные с избирательностью восприятия и с апперцепцией. Еще в прошлом веке многие из этих фактов были хорошо известны. Американский психолог У. Джемс этот феномен поясняет примером: впервые увидев лошадей, жители Полинезии стали называть их свиньями, потому что в их опыте это была наиболее подходящая рубрика. Мальчик, играя с апельсином целую неделю и впервые его увидевшим, стал называть его мячиком.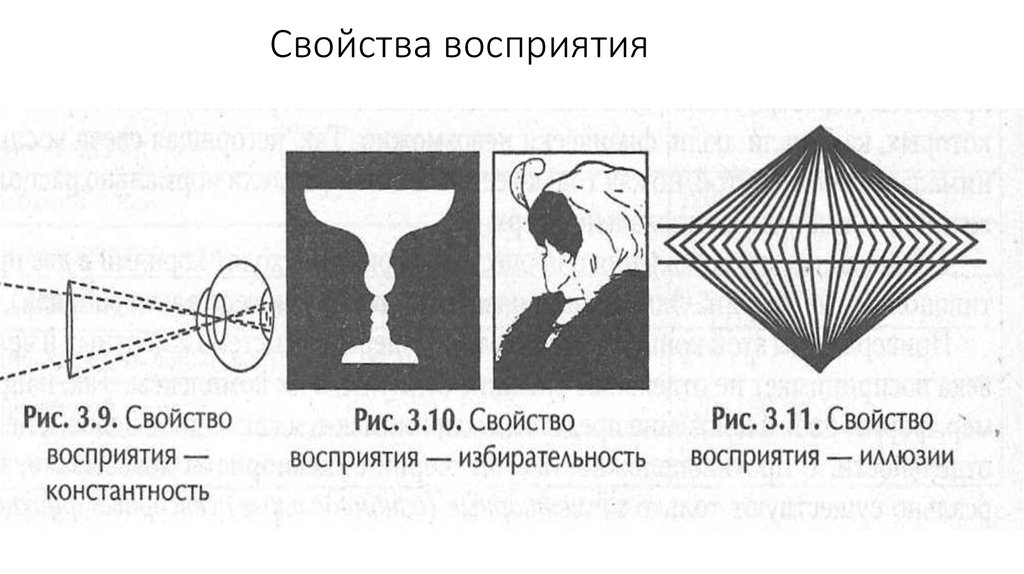 Впервые увиденное цельное яйцо всмятку и вылитое в стакан, ребенок назвал картошкой, потому что видел и ел картофель без кожуры и знал его название.
Впервые увиденное цельное яйцо всмятку и вылитое в стакан, ребенок назвал картошкой, потому что видел и ел картофель без кожуры и знал его название.
В современной психологии феномены, вязанные с избирательностью восприятия, тоже широко изучались. Открытый К. Черри «эффект вечеринки» стал одним из интересных явлений, связанных с этим феноменом и известным многим людям. Суть в следующем – гости разбиваются на группы и каждая обсуждает ту или иную тему. Человеку, зашедшему в комнату случайно, кажется, что разговаривающие должны мешать друг другу. Но стоит только присоединиться к разговору, как все остальное просто не замечается.
Этот феномен изучался с помощью методики дихотического прослушивания. Смысл методики состоит в том, что испытуемому через наушники подавались две различные магнитные записи. Внимательно слушая одну из них, испытуемый легко повторял услышанные слова и фразы, но из другой записи он ничего не мог уловить, даже если слово повторялось много раз.
Такое слежение за сообщениями осуществляется на основе смысловых характеристик – если испытуемый следит, что ему подается на правое ухо и это сообщение начинает вдруг передаваться на левое ухо, то он продолжает следить за этим с легкостью. Эксперименты с дихотическим слежением показывают, что человек обращает внимание при выборе информации в зависимости от своего прошлого опыта.
Эксперименты с дихотическим слежением показывают, что человек обращает внимание при выборе информации в зависимости от своего прошлого опыта.
Кроме устойчивой апперцепции место может иметь и временная апперцепция в которой сказывается влияние ситуативно возникающих психических состояний – установки, эмоции, ожидания и др.
Интересный пример приводит Л.С. Выготский, который говорит, что в процессе преподавания важно учитывать особенности прошлого опыта. Суть заключается в том, что курс советской валюты после революции резко упал, за продукты платили баснословные деньги. Когда на уроке дети услышали, что длина экватора 40 000 верст, то их реакция была далеко не та, которую ожидал учитель. Длину экватора ученики приравняли к стакану семечек, который стоил столько же. Получается, что в опыте детей значение больших чисел было скомпрометировано. Дети не понимали, почему при изображении громадных расстояний, используются такие цифры, о которых у них сложилось впечатление, как о незначительных величинах.
Процесс восприятия и психическое состояние
Человек даже в течение суток может находиться в разном психическом состоянии. Он может быть бодрым или утомленным, активным или пассивным, печальным и радостным и др. Это состояние сказывается на его восприятии и из окружающей социальной среды он выделит те воздействия, которые будут соответствовать его состоянию.
На восприятие окружающего мира большое влияние оказывают богатство и направленность знаний, умений, привычек, возрастной, житейский, профессиональный опыт. Чем больше человек знаком с объектом восприятия, тем больше признаков, сторон, изменений можно в нем заметить.
Особенно важно это при обучении детей. Их восприятие надо организовать так, чтобы они могли легко выделять существенные признаки предметов и явлений, независимо привлекают они к себе внимание или нет.
К благоприятным условиям при решении этой задачи относятся:
- Отчетливость восприятия предмета – расстояние, освещенность, контрастность фона, наличие или отсутствие отвлекающих объектов в поле восприятия;
- Обязательная умственная и практическая активность в процессе восприятия, определение существенных признаков, сравнение с другими объектами, зарисовки, возможность ощупывания и др. ;
- Указания учителя для направления процесса восприятия в нужную сторону;
- Использование специальных приемов выделения замаскированных существенных признаков.
 ;
;Интересную закономерность в своем экспериментальном исследовании обнаружила М.В. Зверева. Если свойство предмета познается при помощи одного анализатора, то и выделять его лучше тоже с помощью одного анализатора, если при помощи нескольких анализаторов, то полезнее комплексное восприятие.
При помощи зрения и осязания человек познает форму, величину, структуру поверхности тела, а это значит, что выделение каждого из этих свойств объекта легче осуществить через зрительно-осязательное восприятие.
19. Восприятие. Физиологические основы восприятия
Восприятие – это целостное отражение предметов, ситуаций, явлений, возникающих при непосредственном воздействии физических раздражителей на рецепторные поверхности органов чувств.
Понятия «ощущение» и «восприятие»
взаимосвязаны, однако между ними
существуют и коренные различия. Суть
процессов ощущения заключается в
отражении лишь отдельных свойств
объектов и явлений окружающего мира.
Однако человек живет не в мире изолированных
световых или цветовых пятен, звуков или
прикосновений, он живет в мире вещей,
предметов и форм, в мире сложных ситуаций.
Все, что бы человек ни воспринимал,
неизменно предстает перед ним в виде
целостных образов. Поэтому основным
отличием восприятия от ощущения является
предметность осознания всего, что
воздействует на человека, т. е.
отображение объекта реального мира в
совокупности всех его свойств, или,
иными словами, целостное отображение
предмета.
Суть
процессов ощущения заключается в
отражении лишь отдельных свойств
объектов и явлений окружающего мира.
Однако человек живет не в мире изолированных
световых или цветовых пятен, звуков или
прикосновений, он живет в мире вещей,
предметов и форм, в мире сложных ситуаций.
Все, что бы человек ни воспринимал,
неизменно предстает перед ним в виде
целостных образов. Поэтому основным
отличием восприятия от ощущения является
предметность осознания всего, что
воздействует на человека, т. е.
отображение объекта реального мира в
совокупности всех его свойств, или,
иными словами, целостное отображение
предмета.
Восприятие включает ощущение и
основывается на нем. При этом всякий
перцептивный образ включает целый ряд
ощущений, так как любой предмет или
явление обладают многими и различными
свойствами, каждое из которых способно
независимо от других свойств вызвать
ощущение. Однако было бы ошибочным
полагать, что такой процесс (от относительно
простых ощущений – к сложному образу
восприятия) является простым суммированием
отдельных ощущений.
В настоящее время существуют различные
теории процесса распознавания образов.
В этих теориях основное внимание уделено
следующему вопросу: как внешние сигналы,
воздействующие на органы чувств,
преобразуются в осмысленные перцептивные
образы? Как правило, человек опознает
окружающие предметы и события легко и
быстро; поэтому может создаться
впечатление, что связанные с распознаванием
операции просты и непосредственны.
Однако это совсем не так. Попытки
инженеров создать машины, которые были
бы способны распознавать символы и
звуки, обычные для окружающей человека
среды, в большинстве случаев заканчиваются
неудачей.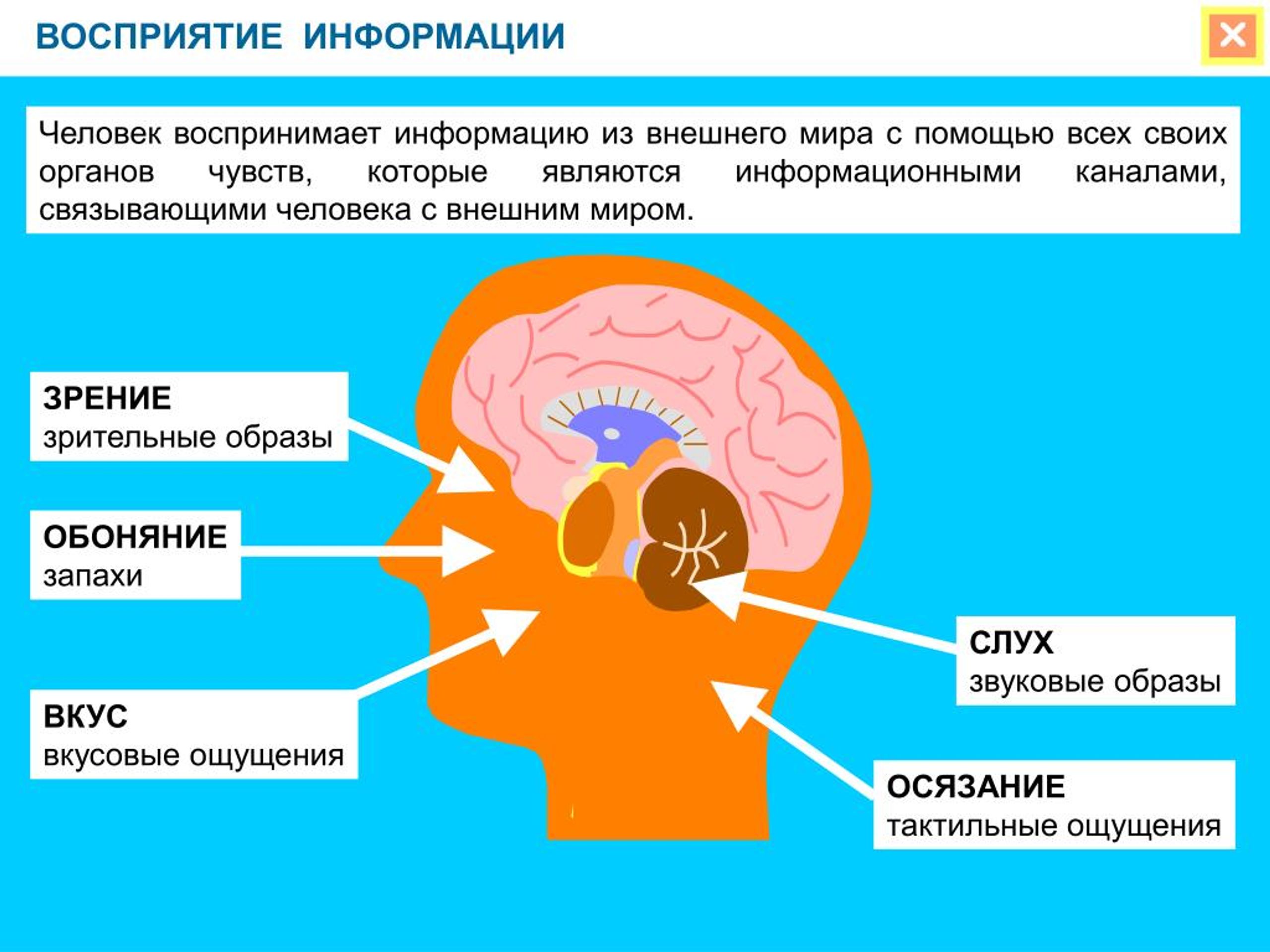
Физиологической основой восприятия являются процессы, проходящие в органах чувств, нервных волокнах и центральной нервной системе. Так, под действием раздражителей в окончаниях нервов, имеющихся в органах чувств, возникает нервное возбуждение, которое по проводящим путям передается в нервные центры и в конечном итоге в кору головного мозга. Здесь оно поступает в проекционные (сенсорные) зоны коры, которые представляют собой как бы центральную проекцию нервных окончаний, имеющихся в органах чувств. В зависимости от того, с каким органом связана проекционная зона, формируется определенная сенсорная информация.
Следует отметить, что описанный выше
механизм является механизмом возникновения
ощущений. И действительно, на уровне
предложенной схемы формируются ощущения.
Следовательно, ощущения могут быть
рассмотрены как структурный элемент
процесса восприятия. Собственные
физиологические механизмы восприятия
включаются в процессе формирования
целостного образа на последующих этапах,
когда возбуждение от проекционных зон
передается в интегративные зоны коры
головного мозга, где и происходит
завершение формирования образов явлений
реального мира.
Это различие отчетливо обнаруживается
при нарушении деятельности той или иной
зоны. Например, при нарушении работы
зрительной проекционной зоны наступает
так называемая центральная слепота,
т. е. при полной исправности периферии
– органов чувств – человек полностью
лишается зрительных ощущений, он ничего
не видит. Совсем иначе обстоит дело при
поражениях или нарушении работы
интегративной зоны. Человек видит
отдельные световые пятна, какие-то
контуры, но не понимает, что он видит.
Он перестает осмысливать то, что
воздействует на него, и не узнает даже
хорошо знакомые предметы. Аналогичная
картина наблюдается при нарушении
деятельности интегративных зон других
модальностей. Так, при нарушении слуховых
интегративных зон люди перестают
понимать человеческую речь. Подобные
заболевания получили название
агностических расстройств (расстройства,
приводящие к невозможности познания),
или агнозий.
Поскольку восприятие тесно связано с
ощущением, можно предположить, что оно,
как и ощущение, является рефлекторным
процессом. Рефлекторную основу восприятия
доказывал И. П. Павлов. Он показал, что
в основе восприятия лежат условные
рефлексы, т. е. временные нервные
связи, образующиеся в коре больших
полушарий головного мозга при воздействии
на рецепторы предметов или явлений
окружающего мира. При этом последние
выступают в качестве комплексных
раздражителей, так как при обработке
вызванного ими возбуждения в ядрах
корковых отделов анализаторов протекают
сложные процессы анализа и синтеза. И.
П. Павлов писал: «В гармонии с беспрерывно
и многообразно колеблющейся природой
агенты в качестве условных раздражителей
то выделялись полушариями для организма
в виде крайне мелких элементов
(анализировались), то сливались в
многообразные комплексы (синтезировались)».
Анализ и синтез обеспечивает выделение
объекта восприятия из окружающей среды,
и на этой основе все его свойства
объединяются в целостный образ.
Временные нервные связи, обеспечивающие процесс восприятия, могут быть двух видов: образуемые в пределах одного анализатора и межанализаторные. Первый вид имеет место при воздействии на организм комплексного раздражителя одной модальности. Например, таким раздражителем является мелодия, представляющая собой своеобразное сочетание отдельных звуков, воздействующих на слуховой анализатор. Весь этот комплекс действует как один сложный раздражитель. При этом нервные связи образуются не только в ответ на сами раздражители, но и на их отношение – временное, пространственное и пр. (так называемый рефлекс на отношение). В результате в коре больших полушарий происходит процесс интегрирования, или сложного синтеза.
Второй вид нервных связей, образуемых
при воздействии комплексного
раздражителя, – это связи в пределах
разных анализаторов, возникновение
которых И. М. Сеченов объяснял существованием
ассоциаций (зрительных, кинестетических,
осязательных и т. д.). Эти ассоциации
у человека обязательно сопровождаются
слуховым образом слова, благодаря
которому восприятие приобретает
целостный характер. Например, если вам
завязать глаза и дать в руки шарообразный
предмет, предварительно сказав, что это
съедобный предмет, и при этом вы можете
ощутить его своеобразный запах,
попробовать его вкус, то вы без труда
поймете, с чем имеете дело. В процессе
работы с этим знакомым, но невидимым
для вас в данный момент предметом, вы
обязательно мысленно назовете его,
т. е. произойдет воссоздание слухового
образа, который по своей сути является
своеобразным обобщением свойств
предмета. В результате человек может
описать даже то, что в данный момент не
наблюдает. Следовательно, благодаря
связям, образуемым между анализаторами,
человек отражает в восприятии такие
свойства предметов или явлений, для
восприятия которых нет специально
приспособленных анализаторов (например,
величина предмета, удельный вес и др.).
Например, если вам
завязать глаза и дать в руки шарообразный
предмет, предварительно сказав, что это
съедобный предмет, и при этом вы можете
ощутить его своеобразный запах,
попробовать его вкус, то вы без труда
поймете, с чем имеете дело. В процессе
работы с этим знакомым, но невидимым
для вас в данный момент предметом, вы
обязательно мысленно назовете его,
т. е. произойдет воссоздание слухового
образа, который по своей сути является
своеобразным обобщением свойств
предмета. В результате человек может
описать даже то, что в данный момент не
наблюдает. Следовательно, благодаря
связям, образуемым между анализаторами,
человек отражает в восприятии такие
свойства предметов или явлений, для
восприятия которых нет специально
приспособленных анализаторов (например,
величина предмета, удельный вес и др.).
Таким образом, в основе сложного процесса
построения образа восприятия лежат
системы внутрианализаторных и
межанализаторных связей, обеспечивающих
наилучшие условия видения раздражителей
и учет взаимодействия свойств предмета
как сложного целого.
Физиологические основы восприятия
2.01.2012
Восприятие, как и ощущение, является рефлекторным процессом. Павлов показал, что в основе восприятия лежат условные рефлексы, временные нервные связи, образующиеся в коре больших полушарий головного мозга при воздействии на рецепторы предметов или явлений окружающего мира. Последние при этом выступают в качестве комплексных раздражителей. В ядрах корковых отделов анализаторов протекают сложный анализ и синтез этих комплексных раздражений. И.П.Павлов пишет об этом: «В гармонии с беспрерывно и многообразно колеблющейся природой, агенты в качестве условных раздражителей то выделялись полушариями для организма в виде крайне мелких элементов (анализировались), то сливались в многообразные комплексы (синтезировались)». Анализ обеспечивает выделение объекта восприятия из фона, на его основе все свойства объекта восприятия объединяются в целостный образ. разрозненные звуки, а слова и фразы. В основе синтеза лежит установление временных нервных связей. В основе восприятия лежат два вида нервных связей: образуемые в пределах одного анализатора, и межанализаторные связи. Первый вид имеет место при воздействии на организм комплексного раздражителя одной модальности. Например, таким раздражителем является мелодия, представляющая собой своеобразное сочетание отдельных звуков, воздействующих на слуховой анализатор. Весь этот комплекс действует как один сложный раздражитель. При этом нервные связи образуются не только в ответ на сами раздражители, но и на их отношение — временное, пространственное и пр. (так называемый рефлекс на отношение). В результате в коре больших полушарий происходит процесс интегрирования, сложный синтез.
разрозненные звуки, а слова и фразы. В основе синтеза лежит установление временных нервных связей. В основе восприятия лежат два вида нервных связей: образуемые в пределах одного анализатора, и межанализаторные связи. Первый вид имеет место при воздействии на организм комплексного раздражителя одной модальности. Например, таким раздражителем является мелодия, представляющая собой своеобразное сочетание отдельных звуков, воздействующих на слуховой анализатор. Весь этот комплекс действует как один сложный раздражитель. При этом нервные связи образуются не только в ответ на сами раздражители, но и на их отношение — временное, пространственное и пр. (так называемый рефлекс на отношение). В результате в коре больших полушарий происходит процесс интегрирования, сложный синтез.
Второй вид нервных связей, образуемых при воздействии комплексного раздражителя, — это связи в пределах разных анализаторов. Сеченов объяснял восприятие предметов или пространства ассоциациями зрительных, кинестезических, осязательных и других ощущений. К этим ассоциациям у человека обязательно присоединяется и слуховой образ слова, которым обозначается данный предмет или пространственное отношение. В акте зрения при восприятии величины предметов, их удаленности и прочего зрительные ощущения всегда ассоциируются с мышечными. Дейетвием определенных лекарственных веществ можно вызвать некоторое нарушение этих связей путем усиления или ослабления глазных мышц. При этом наблюдается макрония (кажущееся увеличение предметов) или микрония (кажущееся уменьшение величины предметов).
К этим ассоциациям у человека обязательно присоединяется и слуховой образ слова, которым обозначается данный предмет или пространственное отношение. В акте зрения при восприятии величины предметов, их удаленности и прочего зрительные ощущения всегда ассоциируются с мышечными. Дейетвием определенных лекарственных веществ можно вызвать некоторое нарушение этих связей путем усиления или ослабления глазных мышц. При этом наблюдается макрония (кажущееся увеличение предметов) или микрония (кажущееся уменьшение величины предметов).
Временные нервные связи, лежащие в основе восприятия, складываются на основе объективных связей свойств предметов или явлений внешнего мира. Благодаря связям, образуемым между анализаторами, мы отражаем в восприятии и такие свойства предметов или явлений, для которых нет специально приспособленных анализаторов (например, величина предмета, удельный вес и пр.). Поэтому в восприятии мы глубже познаем мир, чем в ощущениях.
Таким образом, в основе сложного процесса построения образа восприятия лежат системы внутрианализаторных и межанализаторных связей, обеспечивающих наилучшие условия видения раздражителей и учет взаимодействия свойств предмета как сложного целого.
Ключевые слова: Восприятие
Источник: Рогов Е.И., Общая психология. Курс лекций
| Материалы по теме |
|---|
Диагностика нарушений восприятия в нейропсихологии Нейропсихология: учебник для вузов / М.Е. Баулина. — М.: Издательство ВЛАДОС, 2018. — 391 с… |
Индикаторы осознаваемого и неосознаваемого восприятия … |
Индивидуальные особенности восприятия Кондрашихина О.А., Дифференциальная психология |
Виды восприятия Дубровина И.В., Младший школьник |
Расстройства ощущений, восприятий, представлений Коркина М.В., Практикум по психиатрии |
Сущность взаимопонимания и этапы его установления Ильин Е. |
Основные характеристики приема информации оператором … |
Восприятие младенца Мухамедрахимов Р.Ж., Мать и младенец |
 П., Психология общения и межличностных отношений. — СПб.: Питер, 2009. — 576 с….
П., Психология общения и межличностных отношений. — СПб.: Питер, 2009. — 576 с….Физиологические основы восприятия
Оглавление:
Наше представление о мире стабильно. Наш мир состоит из определенных материальных элементов: камень — это камень, дерево — это дерево, кошка — это кошка. Нам кажется, что иначе быть не может.
Однако мы приспосабливаем мир к нашим собственным стандартам, определяемым нашими человеческими чувствами. Это относительное изображение, совершенно отличное от того, каким может быть ботинок инфузории, летучая мышь или слон. Для одних животных реальность в основном состоит из запахов, которые в значительной степени неизвестны нам, а для других — из звуков, которые в значительной степени не поддаются обнаружению. Каждый вид имеет рецепторы, которые позволяют организму получать информацию, наиболее полезную для его адаптации к окружающей среде, а это значит, что у каждого вида есть свое восприятие реальности.
Каждый вид имеет рецепторы, которые позволяют организму получать информацию, наиболее полезную для его адаптации к окружающей среде, а это значит, что у каждого вида есть свое восприятие реальности.
Окружающая среда посылает нам тысячи сигналов в каждый момент времени, из которых мы можем обнаружить только очень малую часть. Человеческое ухо не способно слышать слишком высокие для него звуки, в то время как собака, дельфин или летучая мышь могут слышать эти ультразвуковые звуки с легкостью. Пространство пронизано бесконечным числом электромагнитных волн, от самых коротких (гамма-лучи, рентгеновские лучи) до самых длинных (радиоволны). Однако наши глаза чувствительны только к небольшой части спектра, которая занимает промежуточное положение — «видимый свет». Как бы выглядела реальность, если бы мы могли различать другие формы энергии? Как бы выглядел наш взгляд на мир, если бы наши глаза могли улавливать рентгеновские снимки и видеть сквозь то, что сейчас кажется непроницаемым? Но какой бы мозг нам понадобился, чтобы воспринимать наше окружение, откуда мы получаем столько сигналов?
Физиологическая основа восприятия.
 Отличие восприятия от ощущения
Отличие восприятия от ощущенияВнешние условия, действующие на наши органы чувств, вызывают субъективный эффект в форме восприятия, без какой-либо противодействующей деятельности со стороны субъекта по отношению к воспринимаемым условиям.
Способность чувствовать дается нам и всем живым существам, у которых с рождения есть нервная система. Только человек и высшие животные наделены способностью воспринимать мир с точки зрения образов; эта способность развивается и совершенствуется через жизненный опыт. В отличие от восприятия, ощущение всегда субъективно связано с реальностью, которая существует вне нас в виде объектов. Чувства внутри нас, воспринимаемые свойства объектов, их изображения локализованы в пространстве. Этот процесс, характерный для восприятия в его отличии от ощущений, называется объективизацией. Другое отличие восприятия, в его развитых формах, от ощущений заключается в том, что результатом восприятия является некое чувство (например, ощущение яркости, громкости, равновесия, сладости и т. д.), в то время как в результате восприятия формируется образ, который содержит комплекс взаимосвязанных различных ощущений, приписываемых человеческому сознанию объекту, явлению, процессу. Для того чтобы объект был воспринят, необходимо провести в отношении него противодействие, направленное на его исследование, построение и уточнение изображения. Индивидуальные ощущения как бы «привязаны» к определенным анализаторам, и достаточно стимулировать их периферийные органы-рецепторы, чтобы ощущения возникли. Образ, возникающий в результате процесса восприятия, предполагает взаимодействие, согласованную работу сразу нескольких анализаторов.
д.), в то время как в результате восприятия формируется образ, который содержит комплекс взаимосвязанных различных ощущений, приписываемых человеческому сознанию объекту, явлению, процессу. Для того чтобы объект был воспринят, необходимо провести в отношении него противодействие, направленное на его исследование, построение и уточнение изображения. Индивидуальные ощущения как бы «привязаны» к определенным анализаторам, и достаточно стимулировать их периферийные органы-рецепторы, чтобы ощущения возникли. Образ, возникающий в результате процесса восприятия, предполагает взаимодействие, согласованную работу сразу нескольких анализаторов.
Таким образом, восприятие функционирует как осмысленный (включающий в себя принятие решений) и значительный (относящийся к языку) синтез множества ощущений, получаемых от целостных объектов или сложных явлений, воспринимаемых как единое целое. Этот синтез действует как изображение заданного объекта или явления, сформированного в процессе его активного отражения.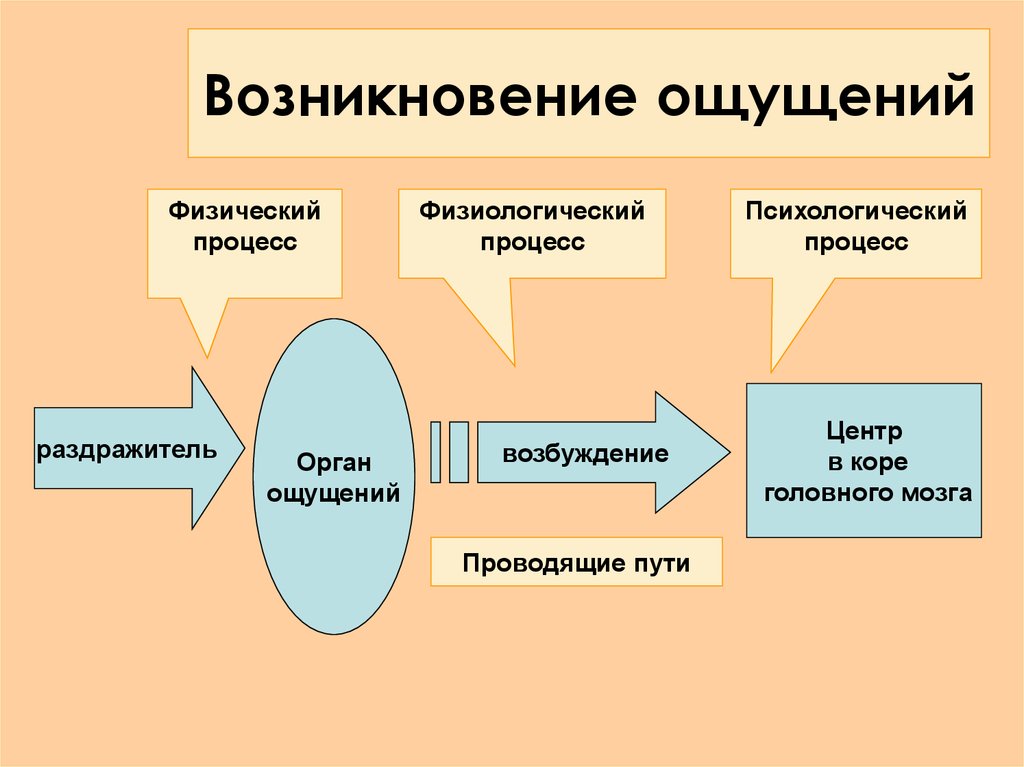
«Все, что воздействует на наши органы чувств, по сравнению с обычным ощущением, вызывает в нас нечто иное: оно возбуждает в мозговых полушариях процессы, которые отчасти обусловлены изменениями в структуре нашего мозга, произведенными в нем предыдущими впечатлениями; в нашем сознании эти процессы порождают идеи, которые тем или иным образом связаны с данным ощущением. Первая такая идея — это представление объекта, которому принадлежит определенное свойство ощущения. Сознание известных материальных объектов до наших органов чувств — это то, что сейчас называется в психологии восприятием».
«Результат сложной аналитико-синтетической работы, которая выделяет некоторые существенные и другие несущественные особенности и собирает воспринимаемые части в значимое целое. Этот сложный процесс размышления над целыми вещами или ситуациями в психологии называется восприятием» (N8 стр. 46).
«Восприятие — это сенсорная репрезентация объекта или явления объективной реальности, которая действует на наши чувства». Человеческое восприятие — это не только сенсорный образ, но и осознание предмета, который выделяется из окружающей среды и сталкивается с объектом. Восприятие чувственно заданного объекта является фундаментальной, наиболее существенной отличительной чертой восприятия» (M9 p.242).
Человеческое восприятие — это не только сенсорный образ, но и осознание предмета, который выделяется из окружающей среды и сталкивается с объектом. Восприятие чувственно заданного объекта является фундаментальной, наиболее существенной отличительной чертой восприятия» (M9 p.242).
Для того чтобы мы могли воспринимать элемент реальности, который нас окружает, энергия (тепловая, химическая, механическая, электрическая или электромагнитная), которую она излучает, должна быть достаточной, чтобы стать стимулом, то есть возбудить один из наших рецепторов. Только тогда, когда в нервных окончаниях одного из наших органов чувств генерируются электрические импульсы, может начаться процесс восприятия. Первичный анализ стимула и кодирование сигнала осуществляется рецепторными клетками, и этот кодированный сигнал затем передается через сенсорные нервы в нервный центр спинного мозга или головного мозга.
Когда сигнал запускается стимулом, который угрожает нанесением вреда организму или направлен на вегетативную нервную систему, очень вероятно, что он сразу же вызовет рефлекторную реакцию, которая возникает в спинном мозге или другом нижнем центре, и это происходит до того, как мы узнаем об эффекте (при ожоге рука дергается, зрачки сжимаются от яркого света). Сигнал продолжает свое путешествие по спинному мозгу, а затем идет двумя разными путями: Один из них ведет в кору головного мозга через таламус (набор ядер серого вещества в мозге, расположенный между срединным мозгом и корой головного мозга, центр, где собираются импульсы от всех органов чувств, кроме обоняния, и происходит их первичный анализ и синтез), а другой проходит через фильтр ретикулярной формации (эта формация простирается по всей оси ствола головного мозга). Он действует как фильтр, который позволяет важным сенсорным сигналам активировать кору, но не пропускает привычные или повторяющиеся сигналы), который держит кору в сознании и решает, является ли сигнал, передаваемый напрямую, достаточно важным, чтобы быть декодированным корой. Если сигнал считается важным, начинается сложный процесс, который приводит к восприятию в истинном смысле этого слова. Этот процесс подразумевает изменения в деятельности многих, тысяч корковых нейронов, которые должны структурировать и организовать сенсорный сигнал, чтобы придать ему смысл.
Сигнал продолжает свое путешествие по спинному мозгу, а затем идет двумя разными путями: Один из них ведет в кору головного мозга через таламус (набор ядер серого вещества в мозге, расположенный между срединным мозгом и корой головного мозга, центр, где собираются импульсы от всех органов чувств, кроме обоняния, и происходит их первичный анализ и синтез), а другой проходит через фильтр ретикулярной формации (эта формация простирается по всей оси ствола головного мозга). Он действует как фильтр, который позволяет важным сенсорным сигналам активировать кору, но не пропускает привычные или повторяющиеся сигналы), который держит кору в сознании и решает, является ли сигнал, передаваемый напрямую, достаточно важным, чтобы быть декодированным корой. Если сигнал считается важным, начинается сложный процесс, который приводит к восприятию в истинном смысле этого слова. Этот процесс подразумевает изменения в деятельности многих, тысяч корковых нейронов, которые должны структурировать и организовать сенсорный сигнал, чтобы придать ему смысл. Первоначально, внимание коры головного мозга к стимулу повлечет за собой серию движений глаз, головы или туловища. Это позволит считывать информацию, поступающую от органа чувств, более глубоко и детально, и, возможно, вовлекать в процесс другие органы чувств. По мере поступления новой информации она связывается со следами аналогичных событий в памяти. Если сигнал оказывается похожим на что-то уже известное, то восприятие приводит к распознаванию. В противном случае она проявляется в восприятии нового аспекта реальности, фиксации в памяти и создании новых следов, которые, в свою очередь, подкрепляются другими актами узнавания. Таким образом, от начала и до конца жизни мозг создает картину реальности, из которой исключаются элементы, не имеющие ничего общего с интересами и потребностями человека.
Первоначально, внимание коры головного мозга к стимулу повлечет за собой серию движений глаз, головы или туловища. Это позволит считывать информацию, поступающую от органа чувств, более глубоко и детально, и, возможно, вовлекать в процесс другие органы чувств. По мере поступления новой информации она связывается со следами аналогичных событий в памяти. Если сигнал оказывается похожим на что-то уже известное, то восприятие приводит к распознаванию. В противном случае она проявляется в восприятии нового аспекта реальности, фиксации в памяти и создании новых следов, которые, в свою очередь, подкрепляются другими актами узнавания. Таким образом, от начала и до конца жизни мозг создает картину реальности, из которой исключаются элементы, не имеющие ничего общего с интересами и потребностями человека.
И.П. Павлов показал, что восприятие основано на условных рефлексах, то есть временных нейронных связях, образующихся в коре больших полушарий под воздействием рецепторов к объектам или явлениям окружающего мира. По сравнению с ощущениями, восприятие является более высокой формой аналитико-синтетической деятельности мозга. Без анализа невозможно осмысленное восприятие. Например, незнакомая иностранная речь воспринимается как непрерывный поток звука. В то же время восприятие речи включает в себя не только анализ, но и синтез, благодаря чему мы воспринимаем не отдельные звуки, а слова. В основе синтеза лежит процесс построения временных нейронных связей. Временные нейронные связи, лежащие в основе восприятия, формируются на основе объективных отношений свойств объектов и явлений внешнего мира. Таким образом, в основе сложного процесса построения перцептивного образа лежит система внутрианализаторных и межанализаторных связей, которая обеспечивает наилучшие условия для идентификации стимулов и учета взаимодействия свойств объекта как сложного целого.
По сравнению с ощущениями, восприятие является более высокой формой аналитико-синтетической деятельности мозга. Без анализа невозможно осмысленное восприятие. Например, незнакомая иностранная речь воспринимается как непрерывный поток звука. В то же время восприятие речи включает в себя не только анализ, но и синтез, благодаря чему мы воспринимаем не отдельные звуки, а слова. В основе синтеза лежит процесс построения временных нейронных связей. Временные нейронные связи, лежащие в основе восприятия, формируются на основе объективных отношений свойств объектов и явлений внешнего мира. Таким образом, в основе сложного процесса построения перцептивного образа лежит система внутрианализаторных и межанализаторных связей, которая обеспечивает наилучшие условия для идентификации стимулов и учета взаимодействия свойств объекта как сложного целого.
Теории восприятия
Наибольшего влияния ассоциативная теория восприятия достигла во второй половине 19 века. Его наиболее выдающимися представителями были немецкие ученые И. Моллер, Э. Моллер, Х. Гельмгольц, Э. Геринг, В. Бундт, Г. Э. Мюллер и американский психолог Э. Б. Титченер. Несмотря на значительные различия в объяснении частичных проблем, эти психологи разделяли общие взгляды на природу восприятия. По их мнению, воспринимаемый нами образ действительно представляет собой сложное объединение первичных элементов сознания — ощущений. В то же время само ощущение понималось как сознательное состояние органа чувств, подверженного воздействию внешнего раздражителя.
Моллер, Э. Моллер, Х. Гельмгольц, Э. Геринг, В. Бундт, Г. Э. Мюллер и американский психолог Э. Б. Титченер. Несмотря на значительные различия в объяснении частичных проблем, эти психологи разделяли общие взгляды на природу восприятия. По их мнению, воспринимаемый нами образ действительно представляет собой сложное объединение первичных элементов сознания — ощущений. В то же время само ощущение понималось как сознательное состояние органа чувств, подверженного воздействию внешнего раздражителя.
Объединение ощущений в восприятие происходит через ассоциации, основанные на близости или сходстве. Этот ассоциативный механизм определяет ведущую роль прошлого опыта в формировании восприятия.
Некоторые из этих авторов, такие как В. Вундт и Г. Гельмгольц, также использовали внутреннюю активность субъекта в форме волевой апперцепции для объяснения процессов синтеза ощущений в восприятие (апперцепция в В. Вундте означает внутреннюю силу, направляющую мысли и ход ментальных процессов) или интеллектуальную бессознательную дедукцию. Фундаментальная ошибка ассоциативной теории заключалась в том, что она опиралась на другие данные для объяснения некоторых субъективных данных, пытаясь таким образом вывести разум из себя. Эта ошибка особенно ярко проявилась в работах структуралистской школы (В. Вундт, Е. Б. Титченер).
Фундаментальная ошибка ассоциативной теории заключалась в том, что она опиралась на другие данные для объяснения некоторых субъективных данных, пытаясь таким образом вывести разум из себя. Эта ошибка особенно ярко проявилась в работах структуралистской школы (В. Вундт, Е. Б. Титченер).
Структуралисты полагали, что поскольку восприятие — это комплекс ощущений, то задача состоит в том, чтобы найти элементарные ощущения в собственном субъективном опыте через интроспекцию, а затем выделить и описать их. Этот метод получил название метода аналитической интроспекции.
Группа немецких психологов: М. Вертхаймер, В. Келлер, К. Коффка и другие выступили с резкой критикой ассоциативности. Они исходили из того, что все процессы в природе изначально целостны. Таким образом, процесс восприятия определяется не отдельными элементарными ощущениями, а целым «полем» стимулов, действующих на организм, структурной ситуацией восприятия в целом. Это направление стало известно как гештальт-психология. Метод аналитической интроспекции был противопоставлен гештальтовской психологией феноменологическому методу, суть которого заключается в непосредственном описании наблюдателем перцептивного содержания.
Метод аналитической интроспекции был противопоставлен гештальтовской психологией феноменологическому методу, суть которого заключается в непосредственном описании наблюдателем перцептивного содержания.
По их мнению, психология восприятия должна дать ответ на вопрос, почему мы видим мир таким, каким он есть. Гесталтисты, как и структуралисты, отказались рассматривать восприятие с точки зрения функции, которой оно служит. Еще одним недостатком является их отрицание историчности восприятия. Прошлые переживания, по их мнению, не могут изменить восприятие объектов, потому что они образуют «хорошую» структуру. Гештальт-психологи собрали большое количество экспериментальных данных, что позволило установить основные закономерности формирования структур в восприятии. Элементы поля объединяются в структуру, которая зависит от таких отношений, как близость, сходство, близость, симметрия и так далее. Обнаружены схемы разделения поля зрения на рисунок и фон. В работах ряда зарубежных психологов были намечены новые способы теоретического анализа восприятия, согласно которым восприятие является результатом активной деятельности субъекта, предоставляющего информацию о внешнем мире.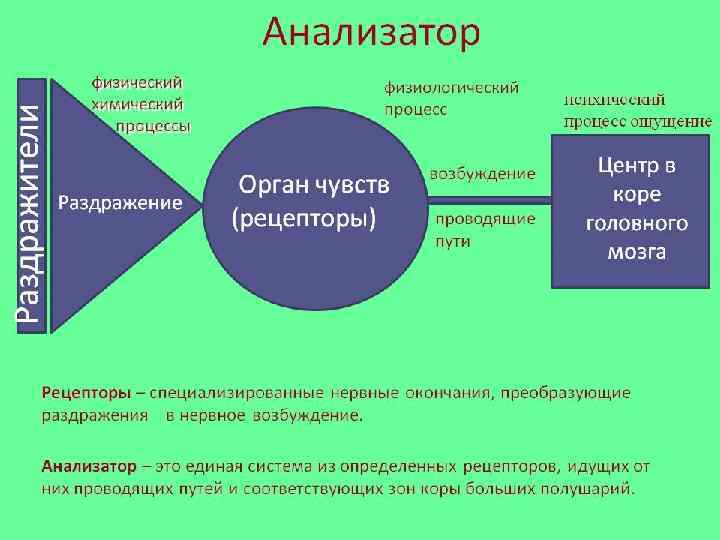 По словам Дж. Гибсона, восприятие интерпретируется им как процесс получения информации об окружающей среде, уменьшающий неопределенность положения тела в ней. Только организованные системы стимулов содержат информацию о внешнем мире. Таким образом, несколько точек, расположенных в порядке уменьшения размера и расстояния друг от друга, образуют так называемый градиент размера и плотности, который несет информацию о протяженности наблюдаемой поверхности в глубину. Поэтому мы воспринимаем поверхность, уходящую вдаль.
По словам Дж. Гибсона, восприятие интерпретируется им как процесс получения информации об окружающей среде, уменьшающий неопределенность положения тела в ней. Только организованные системы стимулов содержат информацию о внешнем мире. Таким образом, несколько точек, расположенных в порядке уменьшения размера и расстояния друг от друга, образуют так называемый градиент размера и плотности, который несет информацию о протяженности наблюдаемой поверхности в глубину. Поэтому мы воспринимаем поверхность, уходящую вдаль.
Деятельность
Активность восприятия заключается, прежде всего, в участии эффекторных компонентов в процессе восприятия, которое происходит в виде движений аппаратов рецепторов и движений тела или его частей в пространстве. Анализ движений рук и глаз делится на два класса. Первый класс включает в себя поиск и позиционирование движений, с помощью которых осуществляется поиск определенного объекта, установка глаза и руки в наиболее благоприятное для восприятия положение, а также изменение этой позиции. Этот класс также включает в себя движения головы для внезапного услышанного звука, отслеживание движений глаз и т.д. Второй класс включает в себя фактические когнитивные движения. При их непосредственном участии происходит оценка размеров, распознаются уже известные объекты, осуществляется фактический процесс построения изображения. Существует непрерывное сравнение изображения с оригиналом. Любое расхождение между ними сразу же приводит к коррекции изображения. Следовательно, роль двигательной активности в восприятии не ограничивается созданием наилучших условий для работы аффективных систем, а сами движения участвуют в формировании субъективного образа объективного объекта.
Этот класс также включает в себя движения головы для внезапного услышанного звука, отслеживание движений глаз и т.д. Второй класс включает в себя фактические когнитивные движения. При их непосредственном участии происходит оценка размеров, распознаются уже известные объекты, осуществляется фактический процесс построения изображения. Существует непрерывное сравнение изображения с оригиналом. Любое расхождение между ними сразу же приводит к коррекции изображения. Следовательно, роль двигательной активности в восприятии не ограничивается созданием наилучших условий для работы аффективных систем, а сами движения участвуют в формировании субъективного образа объективного объекта.
Важность активных движений для развития восприятия подтверждается экспериментами Р. Гехалтена. Для их поведения он использовал специальные очки, которые вызывают оптические искажения. Объект в таких очках впервые видит мир перевернутым или с левой и правой стороны вверх дном. Р. надел очки на свои объекты, которые сдвинули изображение на 20 градусов влево, так что когда они пытались попасть в светящуюся точку с помощью указателя в темной комнате, объекты промахнулись именно на это расстояние. Затем было включено освещение, и испытуемым было разрешено ознакомиться с расположением комнат в лаборатории и их оборудованием, одни испытуемые катались в кресле, а другие передвигались по комнате самостоятельно. Через некоторое время тест на точность был повторен. Было обнаружено, что «активные» испытуемые научились правильно определять положение цели, в то время как «пассивные» испытуемые промахнулись, как и в начале эксперимента.
Затем было включено освещение, и испытуемым было разрешено ознакомиться с расположением комнат в лаборатории и их оборудованием, одни испытуемые катались в кресле, а другие передвигались по комнате самостоятельно. Через некоторое время тест на точность был повторен. Было обнаружено, что «активные» испытуемые научились правильно определять положение цели, в то время как «пассивные» испытуемые промахнулись, как и в начале эксперимента.
Зрительное восприятие включает в себя множество источников информации, выходящих за рамки того, что глаз воспринимает, когда мы смотрим на объект. Процесс восприятия, как правило, включает в себя знания об объекте из предыдущего опыта, и этот опыт не ограничивается зрением. Это еще больше подчеркивает активный процесс восприятия.
Историчность
Восприятие — это система актов восприятия (восприятие — это прямое отражение органами чувств объективной реальности), освоение которой требует специальной подготовки и достаточно длительной практики. Перцептивные акты и критерии адекватности образа не остаются неизменными, но вместе с развитием самой деятельности проходят значительный путь развития. Это означает, что важнейшим свойством восприятия является его историчность — обусловленная конкретными условиями хода действий и прошлым опытом субъекта. Наблюдение за слепым человеком в возрасте десяти месяцев, восстановившим зрение в 52 года, было сделано английским психологом Р. Григорием. Зрительное восприятие этого человека было ограничено прикосновением. Он никогда не учился читать со зрением, но зрительно распознавал заглавные буквы и цифры, которым его учили в школе для слепых. На рисунках этого человека также видна неспособность воспроизвести то, что он ранее не воспринимал прикосновением. Например, он не мог нарисовать переднюю часть автобуса, потому что не мог исследовать его руками.
Перцептивные акты и критерии адекватности образа не остаются неизменными, но вместе с развитием самой деятельности проходят значительный путь развития. Это означает, что важнейшим свойством восприятия является его историчность — обусловленная конкретными условиями хода действий и прошлым опытом субъекта. Наблюдение за слепым человеком в возрасте десяти месяцев, восстановившим зрение в 52 года, было сделано английским психологом Р. Григорием. Зрительное восприятие этого человека было ограничено прикосновением. Он никогда не учился читать со зрением, но зрительно распознавал заглавные буквы и цифры, которым его учили в школе для слепых. На рисунках этого человека также видна неспособность воспроизвести то, что он ранее не воспринимал прикосновением. Например, он не мог нарисовать переднюю часть автобуса, потому что не мог исследовать его руками.
В процессе социального контакта ребенок постепенно усваивает социально развитые системы сенсорных качеств — сенсорные шкалы (А.В. Запорожец). К ним относятся: шкала музыкальных звуков, система геометрических форм, «сетка фонем» родного языка. Если сенсорные стандарты являются результатом социальной и исторической человеческой деятельности, то результат индивидуальной человеческой деятельности по овладению сенсорными стандартами называется оперативными единицами восприятия. Оперативные единицы восприятия функционируют как содержание, которое субъект подчеркивает при выполнении той или иной перцептивной задачи. Развитие восприятия связано с изменением оперативных единиц восприятия. Это изменение выражается в трансформации групп случайных, частных знаков в структурные, интегральные знаки. В результате того, что оперативными единицами восприятия становятся образы объектов и даже модели целых ситуаций, появляется возможность одноуровневого восприятия, независимо от количества знаков, содержащихся в объекте или ситуации. Конечно, задача формирования образа может возникнуть и вне детства. Всякий раз, когда появляются новые вещи или изображение оказывается недостаточным, процесс восприятия снова переходит из одной стадии в другую и осуществляется посредством расширенных перцептивных действий.
К ним относятся: шкала музыкальных звуков, система геометрических форм, «сетка фонем» родного языка. Если сенсорные стандарты являются результатом социальной и исторической человеческой деятельности, то результат индивидуальной человеческой деятельности по овладению сенсорными стандартами называется оперативными единицами восприятия. Оперативные единицы восприятия функционируют как содержание, которое субъект подчеркивает при выполнении той или иной перцептивной задачи. Развитие восприятия связано с изменением оперативных единиц восприятия. Это изменение выражается в трансформации групп случайных, частных знаков в структурные, интегральные знаки. В результате того, что оперативными единицами восприятия становятся образы объектов и даже модели целых ситуаций, появляется возможность одноуровневого восприятия, независимо от количества знаков, содержащихся в объекте или ситуации. Конечно, задача формирования образа может возникнуть и вне детства. Всякий раз, когда появляются новые вещи или изображение оказывается недостаточным, процесс восприятия снова переходит из одной стадии в другую и осуществляется посредством расширенных перцептивных действий.
Таким образом, развитие восприятия приводит к созданию определенного набора образов или моделей восприятия окружающей среды. Если на этапе построения образа объекта системы восприятия становятся похожими на свойства воздействия, то на этапе распознавания или действия, основанного на разработанных оперативных единицах восприятия, свойства и направление процесса существенно изменяются. По мнению А.В. Запорожца, эти изменения заключаются в том, что субъект не только воссоздает образ объекта, но и переводит полученную информацию на язык оперативных единиц восприятия или уже изученных моделей восприятия. И это преобразование приводит к формированию полного адекватного образа.
Субъективность
Третьим важным свойством восприятия является его субъективность. Под объективностью восприятия понимается соотнесение всей информации о внешнем мире с самими объектами. Это способность субъекта воспринимать мир не как набор несвязанных ощущений, а как отдельные объекты, обладающие свойствами, вызывающими эти ощущения.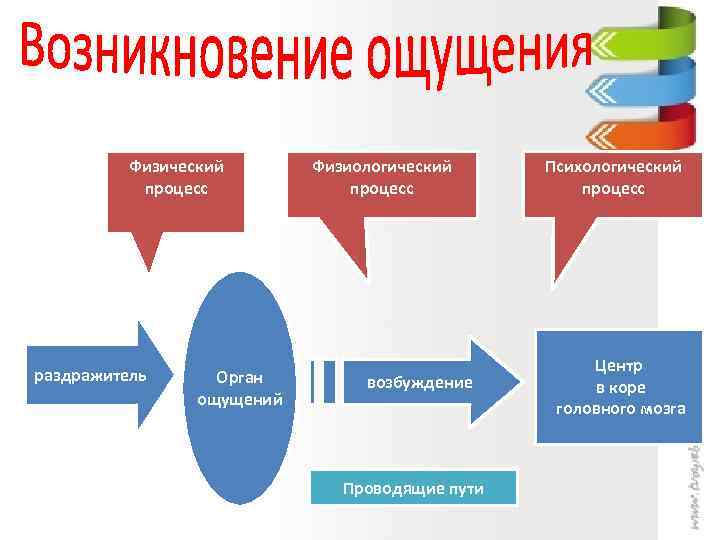 Поскольку перцептивные действия направлены на объективное отражение ситуации, важность объектной среды оказывается решающей для нормального функционирования восприятия. Объект погружался в ванну с физиологическим раствором при комфортной температуре.
Поскольку перцептивные действия направлены на объективное отражение ситуации, важность объектной среды оказывается решающей для нормального функционирования восприятия. Объект погружался в ванну с физиологическим раствором при комфортной температуре.
Во время этого процесса субъект слышал только однообразные ритмические звуки и видел рассеянный белый свет, в то время как покрытие на руках препятствовало приобретению тактильных ощущений. Через несколько часов испытуемые почувствовали беспокойство и попросили прекратить эксперимент. Они отметили возникновение галлюцинаций, а также ухудшение восприятия времени. После опыта у испытуемых наблюдалась дезориентация в пространстве, нарушение восприятия движения, формы, цвета и т.п. Субъективность восприятия проявляется в виде целостности, постоянства и осмысленности воспринимаемого образа.
На странице курсовые работы по психологии вы найдете много готовых тем для курсовых по предмету «Психология».
- Здесь темы рефератов по психологии
Читайте дополнительные лекции:
- Актуальные проблемы психологии мышления — Специфика мыслительного процесса
- Развитие внимания
- Психологические основы бизнес-тренинга как метода профессионального обучения
- Педагогическая диагностика как вид деятельности воспитателя
- Особенности конфликтного поведения у подростков — Психологические особенности подросткового возраста
- Девиантное поведение подростков, его особенности
- Задатки и способности в психологии
- Деятельность психологической службы
- Что такое дизонтогенез в клинической психологии
- Психологическая коррекция ДЦП
Физиологические основы восприятия
Деятельность восприятия как психического процесса обеспечивают процессы, проходящие в органах чувств, нервных волокнах и центральной нервной системе.
Под действием раздражителей в окончаниях нервов, имеющихся в органах чувств, возникает нервное возбуждение, которое по проводящим путям передается в нервные центры и, в конечном итоге, в кору головного мозга. Здесь нервное возбуждение поступает в проекционные (сенсорные) зоны коры, которые представляют собой таким образом центральную проекцию нервных окончаний, имеющихся в органах чувств. Разные проекционные зоны связаны с разными органами чувств, и в зависимости от того, с каким органом связана проекционная зона, формируется определенная сенсорная информация.
Описанный, до этого момента, механизм является механизмом возникновения ощущений. Эти ощущения — практически в буквальном смысле — являются отражением окружающей действительности. Как в зеркале или на фотографии отражены окружающие объекты, так и в проекционных зонах отражаются эти же самые объекты, только в виде нервных возбуждений, от точки к точке.
С ощущений процесс восприятия лишь начинается. Собственные физиологические механизмы восприятия включаются в процессе формирования целостного образа объекта на последующих этапах, когда возбуждение от проекционных зон передается в интегративные зоны коры головного мозга, где и происходит завершение формирования образов явлений реального мира. Поэтому интегративные зоны коры головного мозга, завершающие процесс восприятия, часто называют перцептивными зонами. Их функция существенно отличается от функций проекционных зон.
Собственные физиологические механизмы восприятия включаются в процессе формирования целостного образа объекта на последующих этапах, когда возбуждение от проекционных зон передается в интегративные зоны коры головного мозга, где и происходит завершение формирования образов явлений реального мира. Поэтому интегративные зоны коры головного мозга, завершающие процесс восприятия, часто называют перцептивными зонами. Их функция существенно отличается от функций проекционных зон.
Различие в работе проекционных и интегративных зон обнаруживается, когда у человека нарушена деятельность той или иной зоны. При нарушении работы зрительной проекционной зоны наступает так называемая центральная слепота, т. е. при полной исправности периферии — органов чувств — человек полностью лишается зрительных ощущений, он совсем ничего не видит. Если поражена интегративная зона (при сохранности проекционной), человек видит отдельные световые пятна, какие-то контуры, но не понимает, что он видит. Он перестает осмысливать то, что воздействует на него, не узнает даже хорошо знакомые предметы и людей.
В других модальностях наблюдается аналогичная картина. При нарушении слуховых интегративных зон люди перестают понимать человеческую речь. Подобные заболевания получили название агностических расстройств (расстройства, приводящие к невозможности познания), или агнозий,
Восприятие тесно связано с двигательной деятельностью, эмоциональными переживаниями, мыслительными процессами, и это еще более усложняет понимание физиологических основ восприятия. Начавшись в органах чувств, нервные возбуждения, вызванные внешними раздражителями, переходят в нервные центры, где охватывают собой различные зоны коры, вступают во взаимодействия с другими нервными возбуждениями. Вся эта сложная сеть возбуждений разрастается. Взаимодействующие между собой возбуждения широко охватывают разные зоны коры.
В процессе восприятия большое значение имеют временные нервные связи. Как ручка и листок бумаги помогают считать столбиком, так и временные нервные связи обеспечивают восприятие возможностью делать гипотезы, которые необходимы для глубокого анализа воспринимаемой ситуации. Временные нервные связи, обеспечивающие процесс восприятия, могут быть двух видов:
Временные нервные связи, обеспечивающие процесс восприятия, могут быть двух видов:
— связи, образуемые в пределах одного анализатора,
— межанализаторные связи.
Первый вид связей имеет место при воздействии на организм комплексного раздражителя одной модальности. Например, таким раздражителем является мелодия, представляющая собой своеобразное сочетание отдельных звуков, воздействующих на слуховой анализатор. Весь этот комплекс действует как один сложный раздражитель. При этом нервные связи образуются не только в ответ на сами раздражители, но и на их отношение — временное, пространственное и пр. (так называемый рефлекс на отношение). В результате в коре больших полушарий происходит процесс интегрирования, или сложного синтеза.
Межанализаторные нервные связи образуются при воздействии комплексного раздражителя. Это связи в пределах разных анализаторов, возникновение которых И. М. Сеченов объяснял существованием ассоциаций (зрительных, кинестетических, осязательных и т. д.). Эти ассоциации у человека обязательно сопровождаются слуховым образом слова, благодаря которому восприятие приобретает целостный характер.
д.). Эти ассоциации у человека обязательно сопровождаются слуховым образом слова, благодаря которому восприятие приобретает целостный характер.
Благодаря связям, образуемым между анализаторами, мы отражаем в восприятии такие свойства предметов или явлений, для восприятия которых нет специально приспособленных анализаторов (например величина предмета, удельный вес).
Таким образом, в основе сложного процесса построения образа восприятия лежат системы внутрианализаторных и межанализаторных связей, обеспечивающих наилучшие условия видения раздражителей и учет взаимодействия свойств предмета как сложного целого. Но кроме этого прямо и косвенно на процесс восприятия оказывают влияние разные участки головного мозга. Даже, например, лобные доли оказывают некоторое участие в процессах восприятия, обеспечивая целенаправленность этого процесса.
Литература
Маклаков А. Г. Общая психология. СПб: Питер, 2001.
См.
 также
такжеВосприятие
- Автономная сенсорная меридиональная реакция
- Адекватность ощущения и восприятия
- Антиципация
- Апперцепция
- Взаимоотношение целого и части в восприятии
- Восприятие времени
- Восприятие движения и времени
- Восприятие и его основные свойства
- Восприятие пространства
- Восприятие устной речи
- … и другое
§16. Физиологические основы восприятия. Психология. Учебник для средней школы.
§16. Физиологические основы восприятия
Так как восприятие всегда включает в себя ощущения, получаемые от разных органов чувств, то и физиологические процессы, лежащие в основе восприятия, включают в себя процессы возбуждения, начинающиеся в органах чувств под воздействием предметов внешнего мира и передающиеся по центростремительным нервам в кору мозга. В зависимости от того, каким анализатором осуществляется отражение предметов и явлений внешнего мира или какой анализатор является при этом ведущим, восприятия разделяются на зрительные, слуховые, двигательные, кожные, вкусовые и обонятельные.
Опыты по изучению условных рефлексов показали, что когда на анализатор систематически действует не изолированный раздражитель, а совокупность раздражителей (например, ряд раздражителей, предъявляемых в определённом темпе), то ответная реакция начинает зависеть не от индивидуальности каждого раздражителя, а от своеобразия их связи, их соотношения. Так, в некоторых опытах применялся звуковой раздражитель, прерывающийся определённое количество раз в минуту; оказалось, что рефлекс, выработавшийся на такое прерывистое звучание, не зависит от того, какие именно звуки при этом применяются. То же самое наблюдалось и при световых раздражителях, когда рефлекс вырабатывался на соотношение между раздражителями: одни световые сигналы могли заменяться другими, предъявляемыми с определённой частотой. Это явление было названо И. П. Павловым рефлексом на отношение.
Связи между отдельными раздражениями, которые были вскрыты при изучении рефлексов на отношения, играют существенную роль в процессах восприятия. Образование таких связей в слуховом анализаторе даёт возможность узнавать мелодию по соотношению входящих в неё звуков, несмотря на различие в их абсолютной высоте и силе. Образование таких связей в зрительном анализаторе обеспечивает узнавание контуров фигуры, независимо от размеров, цвета и т. п. Таким образом, восприятие целостных предметов включает в себя восприятие соотношения раздражителей.
Образование таких связей в слуховом анализаторе даёт возможность узнавать мелодию по соотношению входящих в неё звуков, несмотря на различие в их абсолютной высоте и силе. Образование таких связей в зрительном анализаторе обеспечивает узнавание контуров фигуры, независимо от размеров, цвета и т. п. Таким образом, восприятие целостных предметов включает в себя восприятие соотношения раздражителей.
Кроме процессов, протекающих в пределах одного анализатора, огромное значение имеют взаимосвязи анализаторов между собой. Роль временных связей в процессе восприятия можно показать на примере зрительного восприятия величины.
Зрительное восприятие величины предмета зависит прежде всего от величины изображения предмета на сетчатке глаза. Предмет, вдвое более длинный, даст и на сетчатке вдвое более длинное изображение. Однако так обстоит дело лишь при восприятии предметов, одинаково удалённых от глаз. По законам оптики величина изображения предмета на сетчатке изменяется в зависимости от расстояния до предмета. Например, предмет вдвое более длинный, но находящийся на вдвое большем расстоянии, даст изображение такой же длины, как предмет, вдвое более короткий, но находящийся вдвое ближе. Каким же образом нам удаётся точно воспринимать величину предметов, находящихся на разных расстояниях?
Например, предмет вдвое более длинный, но находящийся на вдвое большем расстоянии, даст изображение такой же длины, как предмет, вдвое более короткий, но находящийся вдвое ближе. Каким же образом нам удаётся точно воспринимать величину предметов, находящихся на разных расстояниях?
Существенное значение для процесса зрительного восприятия имеют повороты глаз. При переводе взора с далёкого предмета на близкий зрительные оси глаз сводятся, и происходит поворот глаз во внутрь. При обратном переводе взора с близкого предмета на далёкий имеет место разведение зрительных осей — поворот глаз наружу. Повороты глаз вызываются сокращениями и расслаблениями глазных мышц; возникающие при этом двигательные ощущения, которых мы обычно не замечаем, играют важную роль в восприятии величины предмета.
Важным является и то, что форма хрусталика при приближении и удалении предметов меняется Это достигается опять-таки сокращением и расслаблением особой мышцы, что связано с некоторыми ощущениями, обычно нами не замечаемыми.
Зрительное восприятие величины осуществляется благодаря образованию с самого раннего детства прочной связи между раздражением, поступающим от сетчатки, а также раздражением от мышц, устанавливающих направление глаза, и от мышц, изменяющих кривизну хрусталика применительно к расстоянию. Нервные связи в коре, лежащие в основе зрительного восприятия, вырабатываются постепенно, путём длительного опыта, начинающегося с первых месяцев нашей жизни.
Нервные связи, лежащие в основе зрительного восприятия величины, являются по своему происхождению условным рефлексом. Этот сложный условный рефлекс вырабатывается в раннем детстве, когда знакомство с действительной величиной предметов происходит и путём ощупывания; осязательные раздражения играют таким образом роль подкрепления.
На примере зрительного восприятия величины ясно видно, как благодаря образованию устойчивых нервных связей, лежаших в основе восприятия, достигается возможность познавать во внешнем мире такие свойства, которые не доступны отдельным ощущениям.
Значение восприятия (Смысл восприятия)
Значение восприятия (Смысл восприятия) У каждого человека свои восприятия. Разные люди воспринимают разные вещи в одной и той же ситуации. Больше того, каждый приписывает разный смысл тому, что он воспринимаем. И у одного человека значения могут меняться. Он может
Физиологические факторы
Физиологические факторы Вопреки традиционному представлению о трансе одержимости как о патологии, психологические аспекты ритуального транса обусловлены участием естественных функций правого полушария головного мозга (ГМ). В проведенном Р. Орнштейном обзоре
Физиологические основы памяти
Физиологические основы памяти
Конечно, многие из читателей этой книжки слышали о мнемонике, или «искусстве приобрести хорошую память», но, наверное, очень немногие знают, что такое мнемоника на самом деле. Если бы такое искусство в действительности было возможно, то оно
Если бы такое искусство в действительности было возможно, то оно
§22. Физиологические основы и внешнее выражение внимания
§22. Физиологические основы и внешнее выражение внимания В основе внимания лежит один из важнейших законов работы больших полушарий мозга, подробно изученный И. П. Павловым,— закон индукции нервных процессов (стр. 8). Согласно этому закону, всякий возникающий в коре
53. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ВНИМАНИЯ
53. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ВНИМАНИЯ Говоря о физиологических основах внимания, нельзя не сказать еще о двух очень важных явлениях: об иррадиации нервных процессов и доминанте. Закон индукции нервных процессов, установленный Ч. Шерринг-тоном и широко использованный И.П.
Физиологические и психологические основы внимания
Физиологические и психологические основы внимания
Говоря о внимании, мы в первую очередь имеем в виду активное внимание, физиологические механизмы которого связаны с активной мозговой деятельностью. Следовательно, их изучение возможно только при общем бодрствовании
Следовательно, их изучение возможно только при общем бодрствовании
Физиологические основы внимания
Физиологические основы внимания Первичный факт, в котором выражается внимание, заключается в том, что некоторые моменты, как бы выступающие на передний план, приобретают господствующее, доминирующее значение для течения психических процессов. Физиологической основой
Физиологические основы внимания
Физиологические основы внимания В течение длительного времени психологи и физиологи пытались описать механизмы, которые определяют избирательное протекание процессов возбуждения и лежат в основе внимания. Однако эти попытки долго ограничивались лишь указанием на тот
Физиологические основы памяти
Физиологические основы памяти
Сохранение следов в нервной системеЯвления длительного сохранения следов раздражителя отмечались исследователями на всем протяжении развития животного мира. Многократно наблюдался факт, что однократное раздражение электрическим ударом
Многократно наблюдался факт, что однократное раздражение электрическим ударом
Физиологические реакции
Физиологические реакции Мы упомянули чуть выше, что эмоциональная реакция помимо психического компонента (переживание, отношение) включает и физиологический. Любая эмоция активирует нервную систему, а это запускает определенные изменения в работе эндокринной
Физиологические потребности
Физиологические потребности На самом нижнем уровне пирамиды располагаются потребности, обеспечивающие физическое выживание индивида. Соответственно, они являются наиболее неотложными и обладают самой мощной побудительной силой. Они должны быть удовлетворены хотя бы
§ 2. Физиологические основы эмоций
§ 2. Физиологические основы эмоций
Эмоции и чувства связаны с различным функциональным состоянием головного мозга, возбуждением его определенных подкорковых областей и с изменениями в деятельности вегетативной нервной системы. И. П. Павлов отмечал, что эмоции связаны с
И. П. Павлов отмечал, что эмоции связаны с
Физиологические симптомы
Физиологические симптомы Застенчивые люди описывают следующие физиологические симптомы своего состояния: пульс учащается, сердце колотится, выступает пот, начинается нервная дрожь. Однако мы все испытываем подобные реакции в связи с любой сильной эмоцией – будь то
Потребности физиологические
Потребности физиологические Здесь «все, как у всех»! Еда, вода, жилье, работа, отдых, здоровье, желание избежать боли, трудностей, проблем… и секс, естественно! Мы цивилизованные, современные люди, однако, как и много веков назад, физиологические потребности – сильны. Они
III. Физиологические основы восприятия
Я утверждал, что восприятие достигается путем обработки градиентов поглощенной энергии, свойства которых причинно связаны с свойствами объектов в силу физических законов, определяющих взаимодействие объектов и различных форм энергии.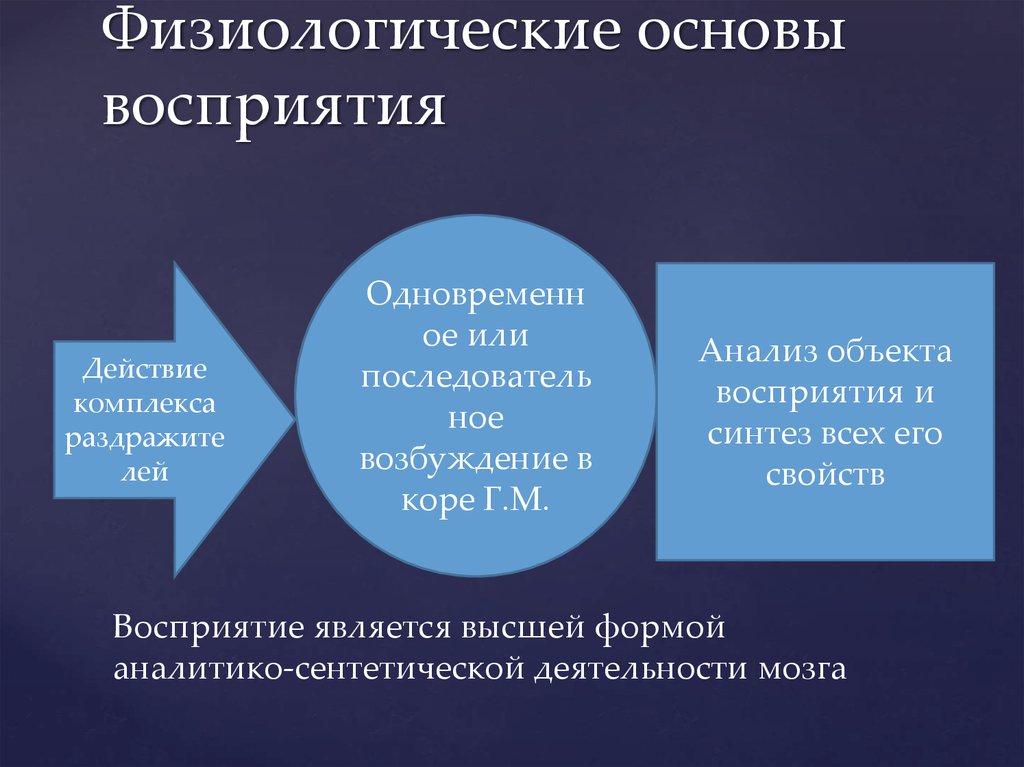 Понимание физиологических основ восприятия требует, чтобы мы определили:
Понимание физиологических основ восприятия требует, чтобы мы определили:
(1) как энергетические градиенты получаются активным организмом и применяются к энергочувствительным рецепторным тканям, и
(2) Как объектные атрибуты градиентов энергии выделяются и сегрегируются сенсорной системой на физиологическом уровне. {17}
A. Выборка и отображение энергетической структуры.
В целом существует два способа получения градиентов энергии, необходимых для восприятия объекта: Рецептор может путем прямого контакта улавливать градиенты энергии, существующие на поверхности объектов. Я назову это сбором контактных градиентов . Вторая основная категория улавливания градиента энергии состоит из перехвата рецептором паттернов окружающей энергии, проходящей через среду между организмом и объектами, составляющими его среду. Этот последний метод называется улавливанием градиентов энергии окружающей среды.
Датчик контактного градиента
Способ получения контактного градиента, который является почти универсальным для воспринимающих организмов, заключается в механическом зондировании и подметании. Как правило, к объекту прикладывают какой-либо придаток организма, содержащий рецепторы, чувствительные к механическим деформациям, либо серией механических контактов и отдергиваний, т. контакт. Исследуемый объект не обязательно должен находиться в прямом контакте с телом организма. Человек, например, может ощупывать предметы с помощью палки или трости. Во всех случаях механического ощупывания и подметания источником механической энергии, ответственной за изменение паттернов давления, являются собственные мышечные усилия организма. Таким образом, эти градиенты равны организм заряжен энергией , и его можно противопоставить градиентов, заряженных внешней энергией.
Как правило, к объекту прикладывают какой-либо придаток организма, содержащий рецепторы, чувствительные к механическим деформациям, либо серией механических контактов и отдергиваний, т. контакт. Исследуемый объект не обязательно должен находиться в прямом контакте с телом организма. Человек, например, может ощупывать предметы с помощью палки или трости. Во всех случаях механического ощупывания и подметания источником механической энергии, ответственной за изменение паттернов давления, являются собственные мышечные усилия организма. Таким образом, эти градиенты равны организм заряжен энергией , и его можно противопоставить градиентов, заряженных внешней энергией.
Модели давления могут отображаться во времени, как в случае зондирования антеннами, палкой или тростью; или модели давления могут отображаться в виде распределенных в пространстве градиентов, как в случае зондирования {18} рукой или приведения объекта в кожный контакт с телом.
В любом случае контактные градиенты имеют тот же последовательный порядок, что и границы и поля объектов, которые их вызывают.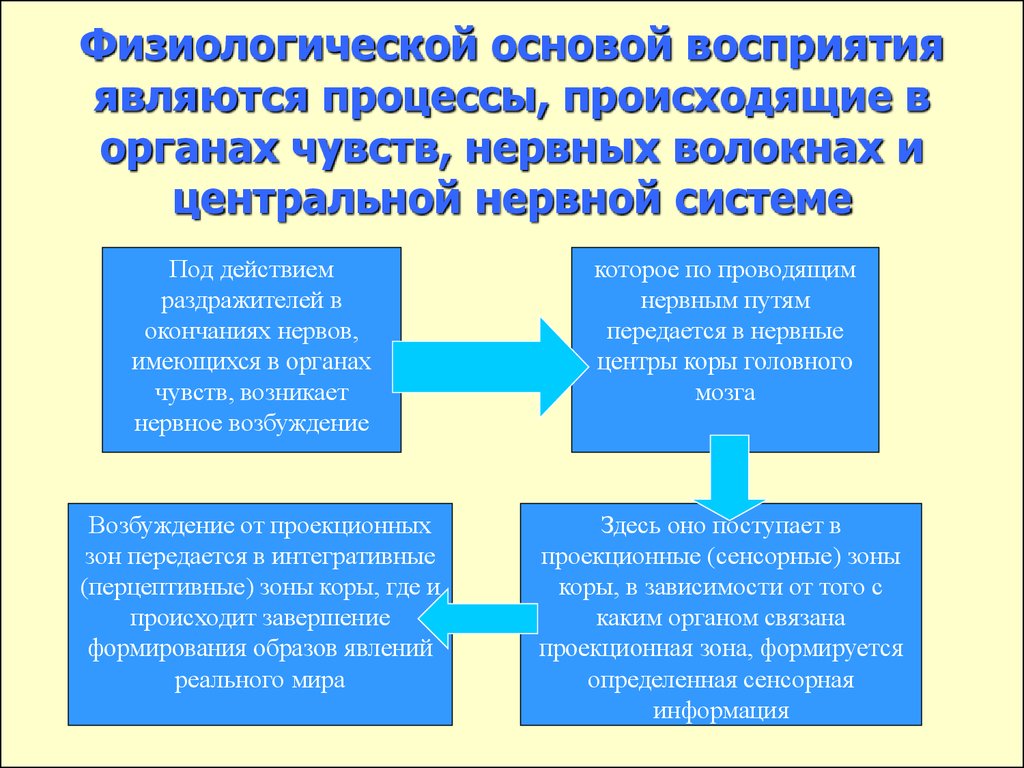 Например, на рисунке 1 обратите внимание, что поле M2 зафиксировано между полем M1 и полем M3. При непрерывном проведении зондом по поверхности этого объекта неправильной формы временной порядок «прогибов», регистрируемых на конце зонда организма, всегда будет соответствовать пространственному порядку краев (или впадин) независимо от длины зонда или угла, под которым применяется зонд. Если направление движения зонда меняется на противоположное, то порядок встречающихся границ соответственно меняется на обратный и, следовательно, изменяется временной порядок регистрируемых отклонений.
Например, на рисунке 1 обратите внимание, что поле M2 зафиксировано между полем M1 и полем M3. При непрерывном проведении зондом по поверхности этого объекта неправильной формы временной порядок «прогибов», регистрируемых на конце зонда организма, всегда будет соответствовать пространственному порядку краев (или впадин) независимо от длины зонда или угла, под которым применяется зонд. Если направление движения зонда меняется на противоположное, то порядок встречающихся границ соответственно меняется на обратный и, следовательно, изменяется временной порядок регистрируемых отклонений.
Те же факты соответствующего заказа сохраняются, конечно, независимо от того, является ли зонд механическим датчиком, термодатчиком, химическим датчиком или фотодатчиком.
Кроме того, когда объект проводится несколькими датчиками, например, при проведении кончиками пальцев по объекту или при проведении объекта по поверхности тела, все «края» стимула, вызванные краями объекта, перемещаются по коже вместе и по тому же курсу.
Улавливание градиентов контакта с окружающей средой
Окружающая энергия относится к энергетическим волнам, сходящимся в каждой точке окружающей среды из нескольких источников в окружающей среде. Источники окружающей энергии бывают либо первичными (излучатели энергии), либо вторичными (отражатели и передатчики энергии). Кроме того, окружающие градиенты могут быть либо возбуждены организмом, например, в случае высокочастотного звукового излучения летучих мышей и морских свиней, либо возбуждены извне, например, светом, излучаемым солнцем, или производимым окружающей средой звуком, которые являются наиболее широко доступны для организмов, живущих над землей или в воде. {19}
Для целей нашего анализа формы волновой энергии могут быть расположены вдоль континуума в соответствии со степенью, в которой каждая форма дифрагирует или не дифрагирует на краях объектов. В общем, свойство волновой энергии зависит от длины волны (или частоты). Коротковолновая (высокочастотная) волновая энергия, такая как свет, пренебрежимо мало дифрагирует на краях объектов, и поэтому говорят, что они распространяются по «прямым линиям». Длинноволновая (низкочастотная) волновая энергия, такая как звук, сильно преломляется и поэтому легко огибает края объектов.
Длинноволновая (низкочастотная) волновая энергия, такая как звук, сильно преломляется и поэтому легко огибает края объектов.
Рис. 1.
Пикап контактных градиентов. Пространственный порядок на поверхности объекта регистрируется как соответствующий временной порядок отклонения на конце анатомического зонда, принадлежащем организму. (См. текст на стр. 18.)
Угловое положение анатомического датчика регистрирует относительное направление к каждой границе или границе удаленного объекта. (См. текст на стр. 26.) {20}
Улавливание градиентов окружающего света
Вследствие того, что свет распространяется по прямым линиям, границы объектов, через которые происходит поглощение или отражение лучистой энергии, резко изменяются, производит запасы энергии , которые уносятся от объекта по прямым линиям. Эти поля могут быть уловлены и отображены на поверхности рецептора или нейронной сети организмом, находящимся в точке окружающей среды, подвергающейся воздействию окружающей лучистой энергии. Гибсон (1966) называет такую точку окружающей среды, доступную организму для потенциального приема окружающей энергии: станционная точка .
Гибсон (1966) называет такую точку окружающей среды, доступную организму для потенциального приема окружающей энергии: станционная точка .
Необходимым условием для отображения запасов энергии, определяющих оптическую структуру в точке станции, является наличие анатомической структуры, способной регистрировать изменение интенсивности или частоты в зависимости от разностей направлений или изменений ориентации рецепторов. Глаз с камерами у позвоночных и сложный глаз насекомого представляют собой две анатомические структуры, выполняющие функцию отображения оптической структуры окружающей среды, полученной при 9-кратном увеличении.0011 точка станции . {21}
В камерном глазу энергетические запасы, составляющие «изображение» сетчатки, представляют собой специфичные для направленности регистрации пространственных отношений границ проецируемых объектов, которые являются их отдаленной причиной. В отличие от контактных градиентов окружающие градиенты создают поля, порядок которых сам по себе не соответствует пространственному порядку границ объектов, которые являются их отдаленной причиной.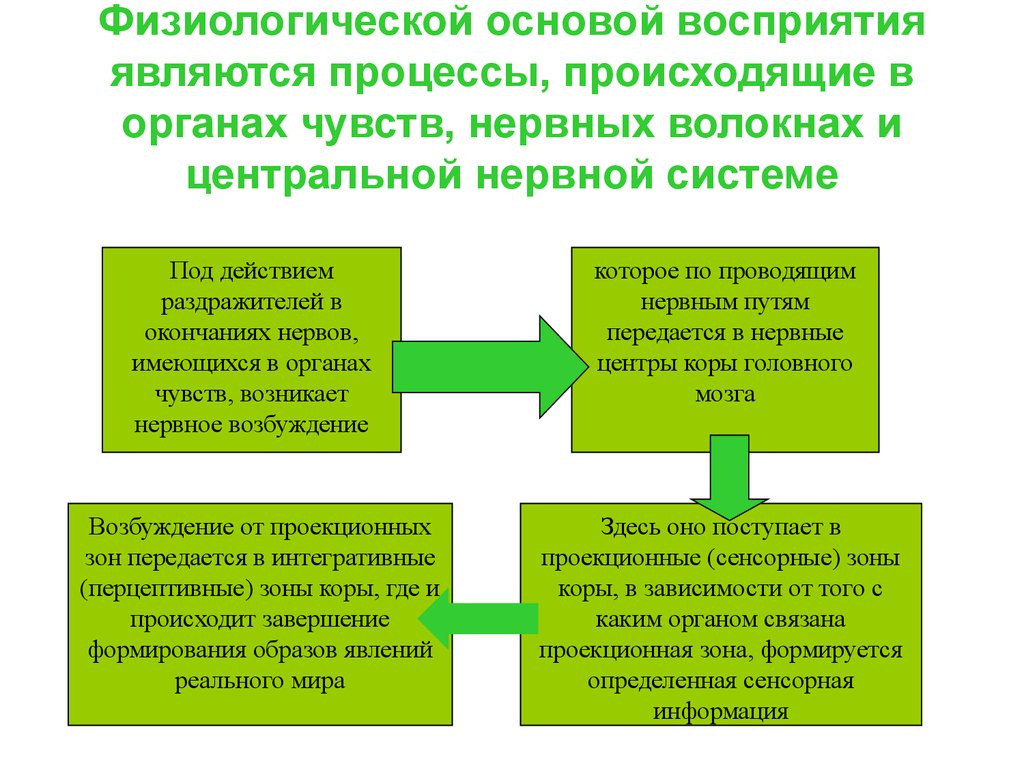 Кроме того, движение глаза не приводит к равномерному движению оптических границ. Оптические поля перемещаются с разной скоростью (относительно друг друга). Различия в скоростях перемещения границ обусловлены и определяют относительное изменение направления от каждой границы объекта к каждой последовательно выбранной точке станции. Совпадение полей указывает на совпадение направления; .к каждой соответствующей границе объекта. Кроме того, скорость изменения направления, которая вызывает относительную скорость движения границ, определяет различия в расстояниях до границ объектов. Эти последние различия определяют пространственное расположение объекта к объекту и поверхностей и частей объекта друг к другу.
Кроме того, движение глаза не приводит к равномерному движению оптических границ. Оптические поля перемещаются с разной скоростью (относительно друг друга). Различия в скоростях перемещения границ обусловлены и определяют относительное изменение направления от каждой границы объекта к каждой последовательно выбранной точке станции. Совпадение полей указывает на совпадение направления; .к каждой соответствующей границе объекта. Кроме того, скорость изменения направления, которая вызывает относительную скорость движения границ, определяет различия в расстояниях до границ объектов. Эти последние различия определяют пространственное расположение объекта к объекту и поверхностей и частей объекта друг к другу.
Рисунок 2.
Улавливание градиентов окружающего света. По мере того, как глаз с камерами в кружке перемещается из положения P1 в P2, отображаемые поля A», B», C», D» смещаются по сетчатке с разной скоростью. Эти поля являются продуктом границ объектов, структурирующих окружающий свет.
 (Прямоугольные формы O 1 и O 2 обозначают удаленные объекты.) (См. текст на стр. 22.){22}
(Прямоугольные формы O 1 и O 2 обозначают удаленные объекты.) (См. текст на стр. 22.){22} Я сказал, что функция глаза состоит в том, чтобы отображать структуру оптической энергии, определяемую различиями значений интенсивности и частоты в разных направлениях. Я также сказал, что так называемое «изображение» сетчатки представляет собой пространственное отображение этой оптической структуры. Но оптическую структуру можно уловить и зарегистрировать, не создавая «изображения»; сложный глаз насекомого тому пример. Эта версия глаза улавливает разницу энергий в разных направлениях, но не дает изображения. Это достигается с помощью множества оптических «воронок», называемых оматидиями, каждая из которых направлена в разные стороны. Массив оматидиев расположен так, что их проксимальные концы, если их вытянуть, сходятся в общей точке в центре глаза насекомого, а {23} так, что их дистальные концы вместе составляют сферическую внешнюю поверхность глаза насекомого. Сложный глаз — яркая иллюстрация того факта, что изображение на сетчатке как таковое, как впервые определил Гибсон, совершенно не связано с сущностью зрительного восприятия. Изображение на сетчатке представляет собой лишь один из способов , отображающий изменяющиеся или текущие оптические градиенты, замеренные в точке станции, и не представляет отображение жесткой структуры «представленных объектов».
Сложный глаз — яркая иллюстрация того факта, что изображение на сетчатке как таковое, как впервые определил Гибсон, совершенно не связано с сущностью зрительного восприятия. Изображение на сетчатке представляет собой лишь один из способов , отображающий изменяющиеся или текущие оптические градиенты, замеренные в точке станции, и не представляет отображение жесткой структуры «представленных объектов».
Улавливание градиентов окружающего звука
Улавливание и отображение звуковых образов отличается от улавливания и отображения оптических изображений в силу того факта, что звук легко огибает края объектов вследствие что звук распространяется с почти одинаковой энергией во всех направлениях от края или границы объекта. Границы объектов не создают резких акустических границ. Это исключает определение границ объекта с точки зрения акустических градиентов (по крайней мере, для частотного диапазона звука, слышимого человеком, а именно от 2 OHZ до 20 000 Гц). Еще одним следствием отсутствия резких акустических границ является то, что направленность удаленного акустического события не может быть определена в одной точке станции.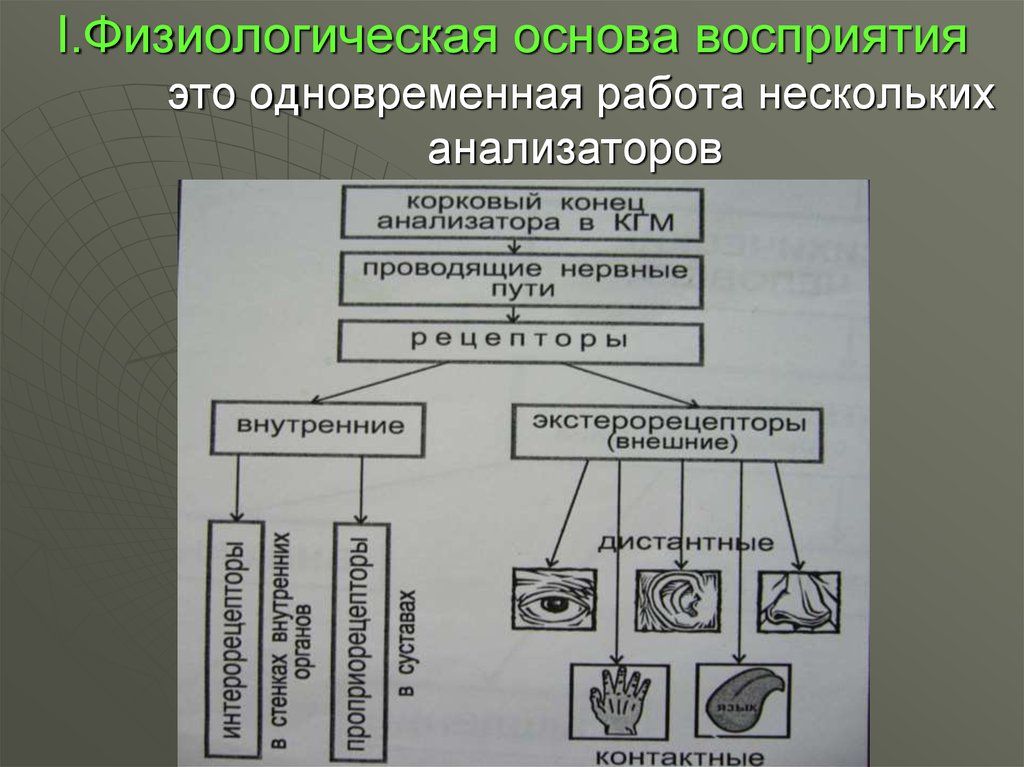 С другой стороны, в отличие от света, звук распространяется с относительно низкой скоростью (около 750 миль в час в воздухе). Эта скорость достаточно мала, чтобы позволить нервной ткани по-разному реагировать на разницу во времени прихода одного и того же звукового события, зарегистрированного в двух (или более) одновременно занятых точках станции. Конечно, разность интенсивности звукового события может быть зарегистрирована и в одновременно занятых точках станции. Два уха, работающие как единый сенсорный орган, регистрируют частотный спектр звука, а также различия в интенсивности и времени ожидания удаленного звукового события в точках станции, занятых каждым из {24} двух ушей. Внутреннее ухо преобразует распределенный во времени энергетический паттерн (в овальном окне) в пространственно распределенный паттерн или градиент (поперек барабанной перепонки). Временные, частотные и энергетические параметры акустического события мы отображали в виде градиента движения вдоль базилярной мембраны.
С другой стороны, в отличие от света, звук распространяется с относительно низкой скоростью (около 750 миль в час в воздухе). Эта скорость достаточно мала, чтобы позволить нервной ткани по-разному реагировать на разницу во времени прихода одного и того же звукового события, зарегистрированного в двух (или более) одновременно занятых точках станции. Конечно, разность интенсивности звукового события может быть зарегистрирована и в одновременно занятых точках станции. Два уха, работающие как единый сенсорный орган, регистрируют частотный спектр звука, а также различия в интенсивности и времени ожидания удаленного звукового события в точках станции, занятых каждым из {24} двух ушей. Внутреннее ухо преобразует распределенный во времени энергетический паттерн (в овальном окне) в пространственно распределенный паттерн или градиент (поперек барабанной перепонки). Временные, частотные и энергетические параметры акустического события мы отображали в виде градиента движения вдоль базилярной мембраны.
Если при вращении головы совместно учитывать разницу между латентностью и интенсивностью между ушами, получаются следующие характеристики:
1. Без изменений разница между интерауральными задержками плюс неизменный интер; различия в интенсивности звука указывают на источник звука внутри или на голове.
2. Изменение межушных различий в задержке или изменение внутриушных различий в интенсивности указывает источник звука вне головы и вдали от нее.
3. скорости , при которых самодвижение коррелирует с изменением межушной латентности и межушной разницы интенсивности , укажите оба относительных направлений и относительных расстояний от головы нескольких источников звука: (а) чем больше скорость изменения при вращении головы, тем ближе источник звука; б) при повороте головы в сторону звука интенсивность звука в ведущем ухе уменьшается, а в ведомом ухе увеличивается, при этом латентность звука в ведущем ухе увеличивается, а в ведомом ухе уменьшается; в) в этих условиях поворота головы, когда интерауральные различия латентности и интенсивности достигают нуля, голова обращена в сторону источника звука.
Ультразвуковой датчик и дисплей . Летучие мыши, морские свиньи, киты и различные виды птиц и грызунов используют издаваемые ими самими высокочастотные звуки, помогающие им ориентироваться в окружающей среде. Важнейшими среди таких {25} существ являются летучие мыши, некоторые виды которых могут излучать ультразвуковые импульсы в диапазоне частот 450 000 Гц (Milne &’Milne, 1962). В таких диапазонах частот звук ведет себя как свет, двигаясь почти по прямой линии, создавая достаточно резкие акустические границы по краям и углам объектов. Таким образом, ультразвуковая энергия способна передавать характеристики границ объекта. Только с помощью ультразвука летучие мыши и морские свиньи способны различать объекты, имеющие разную форму или разную текстуру (Van Bergeijk, et al., 19).60; Стивенс и Варшофски, 1965).
Органом ультразвукового улавливания является ухо, которое регистрирует спецификацию границы объекта с точки зрения изменения акустических паттернов, которые регистрируются с течением времени при вращении головы или перемещении тела в воздухе или воде.
Действия летучих мышей и морских свиней доказывают, что нет ничего в природе звука или слухового представления как такового, что препятствует восприятию объекта с помощью звуковой энергии, улавливаемой ушами.
Порядок как сущность отображения.
Рецепторный аппарат в целом представляет собой устройство для «улавливания» и отображения паттерна или порядка, содержащегося в изменяющихся градиентах энергии. Поскольку именно порядок является фундаментальным, а не форма, в которой передается порядок, сенсорная система может улавливать паттерны, переносимые как временной порядок в энергетическом событии, и отображать эти паттерны в терминах пространственных смежностей. (как это происходит на базилярной мембране). Или пространственный порядок в энергетическом паттерне может быть отображен с точки зрения временные смежности (как в оптическом, тактильном и акустическом сканировании ). Способность «обменять пространство на время» (Gibson, 1966), по-видимому, является неотъемлемым свойством сенсорных систем.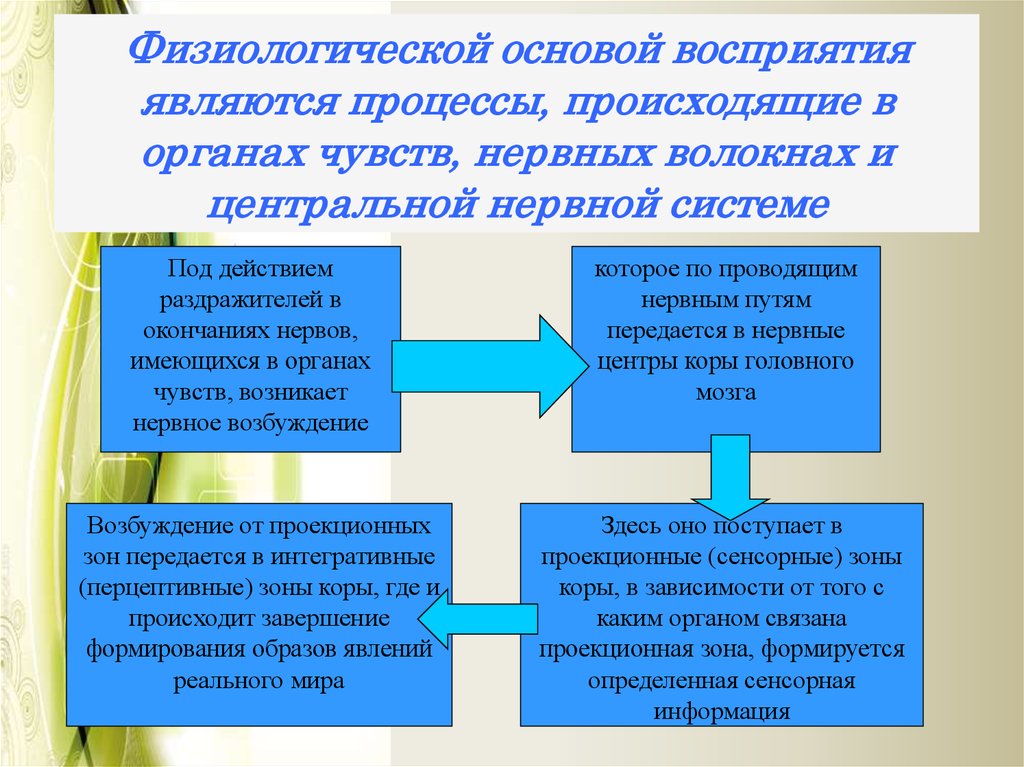 {26}
{26}
Спецификация пространства
Все пространственные атрибуты объектов, в том числе: (а) расстояние от объекта до организма, (б) расстояния между объектами, (в) пространственная форма объектов, (г) расположение окружающей среды, и (e) расположение собственных частей тела организма, указанное у рецептора с точки зрения направленность различия связанные с разными компонентами зарегистрированных энергетических паттернов. Этот принцип пространственной спецификации с помощью зарегистрированных различий в направленности пересекает все сенсорные модальности. Мы уже отмечали, как различия в направленности определяют пространственные атрибуты для поглощения окружающей энергии. Тот же принцип применим к улавливанию контактных градиентов. В случае контактного захвата направленность регистрируется по изменению и скорости изменения углового положения рецепторных придатков организма друг относительно друга и относительно тела организма по мере того, как организм прощупывает или подметает объект (см.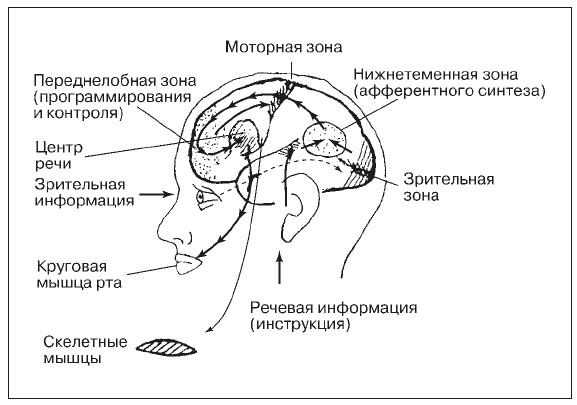 1).
1).
Спецификация пространства/объекта и самостоятельное движение.
Спецификация пространственных атрибутов и, следовательно, спецификация объектов зависит от улавливания изменений в направленности энергии, когда организм перемещает свое тело или части тела в пространстве. Спецификация объекта требует активного, самоинициируемого движения, а также активного самоинициируемого сопротивления движению. Это требует, чтобы организм был по большей части свободен от пассивного движения, навязываемого ему окружающей средой, а это означает, что организм должен обладать телом достаточной массы и конструкции, чтобы сопротивляться силам, создаваемым нормальной турбулентностью и потоком воздуха. его естественная среда, будь то воздух или вода. Как Флорес (1973) правильно идентифицированы:
Движение с внешним давлением исключает их осознание. Осознанность требует сопротивления, способности не уступать, несоответствия между собственными движениями тела и движениями в пространстве вокруг него.
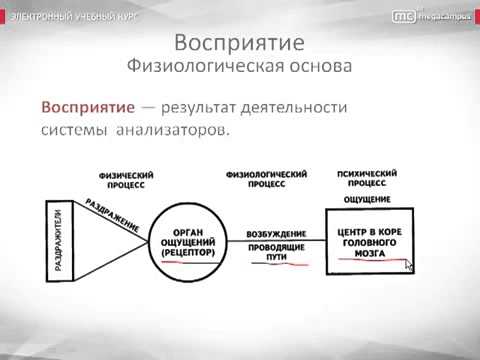 (стр. 124) {27}
(стр. 124) {27}Поскольку для сбора информации об объекте требуется, чтобы организмы были способны двигать частями своего тела, органами или придатками, сохраняя при этом свое место в окружающей среде, а также способные перемещать свои тела в окружающей среде, организму требуется сократительная или мышечная система. А поскольку двигательные действия организма должны быть функционально интегрированы и регулироваться энергочувствительными сенсорными системами, двигательная система и сенсорная система должны совместно действовать через нервную систему. Нервная система, следовательно, должна быть способна, с одной стороны, реагировать на градиенты энергии, а с другой стороны, быть способна осуществлять двигательные действия и контроль. Короче говоря, нервная система должна быть 9-й опорой организма.0011 сенсорно-моторный интегратор .
В этом разделе я обсудил проблему того, как, в принципе, определяющие объект энергетические градиенты улавливаются или перехватываются рецепторным аппаратом и как характеристики объектов отображаются в энергочувствительной ткани рецептора. Наличие объектоспецифических градиентов энергии, улавливание и отображение этих градиентов являются необходимыми условиями восприятия объектов. Но тот факт, что характеристики объектов отображаются на рецепторе, не объясняет, как эти характеристики физиологически изолированный . Это проблема, которая будет обсуждаться в следующем разделе.
Наличие объектоспецифических градиентов энергии, улавливание и отображение этих градиентов являются необходимыми условиями восприятия объектов. Но тот факт, что характеристики объектов отображаются на рецепторе, не объясняет, как эти характеристики физиологически изолированный . Это проблема, которая будет обсуждаться в следующем разделе.
B. Физиологическая изоляция и разделение энергетических атрибутов объекта.
Физиологическое объяснение в психологии
Большая часть наших современных знаний о физиологии восприятия состоит из выводов из психологических данных. Это необходимый и правильный подход к изучению {28} физиологии восприятия, поскольку нет другого способа определить, что представляет собой физиологический механизм, кроме как сначала определить, что именно представляет собой физиологический механизм.0011 функция должна быть достигнута. Физиологическая единица – это функциональная единица. Физиолог должен искать механизм, объясняющий функциональные достижения. Нельзя построить психологию из физиологических «первичных», ибо без предварительного знания принципов, управляющих функциональной целью, не может быть настоящей физиологии. По этой причине физиология не является самостоятельной дисциплиной внутри биологии. Физиолог не может работать один. Принципов физиологии как таковой нет. Существуют только принципы [физиологии обмена веществ, дыхания или размножения, роста, иммунологии, старения и т. д.] Это в равной степени верно и для физиологии восприятия. Как заключает Роберт Эфрон в своей статье «Что такое восприятие»:
Нельзя построить психологию из физиологических «первичных», ибо без предварительного знания принципов, управляющих функциональной целью, не может быть настоящей физиологии. По этой причине физиология не является самостоятельной дисциплиной внутри биологии. Физиолог не может работать один. Принципов физиологии как таковой нет. Существуют только принципы [физиологии обмена веществ, дыхания или размножения, роста, иммунологии, старения и т. д.] Это в равной степени верно и для физиологии восприятия. Как заключает Роберт Эфрон в своей статье «Что такое восприятие»:
Феномены сознания должны быть поняты, прежде чем можно будет надеяться «объяснить» их с точки зрения нервной деятельности. Попытка многих нейрофизиологов изменить этот порядок — изучить нейронные механизмы, лежащие в основе восприятия до , до любого адекватного определения или концептуализации восприятия — обречена на неудачу. (Efron 1969, p. 171)
Восприятие и психофизика
В предыдущем разделе указывалось, что психологические переживания яркости, громкости, цвета, высоты тона, твердости и т.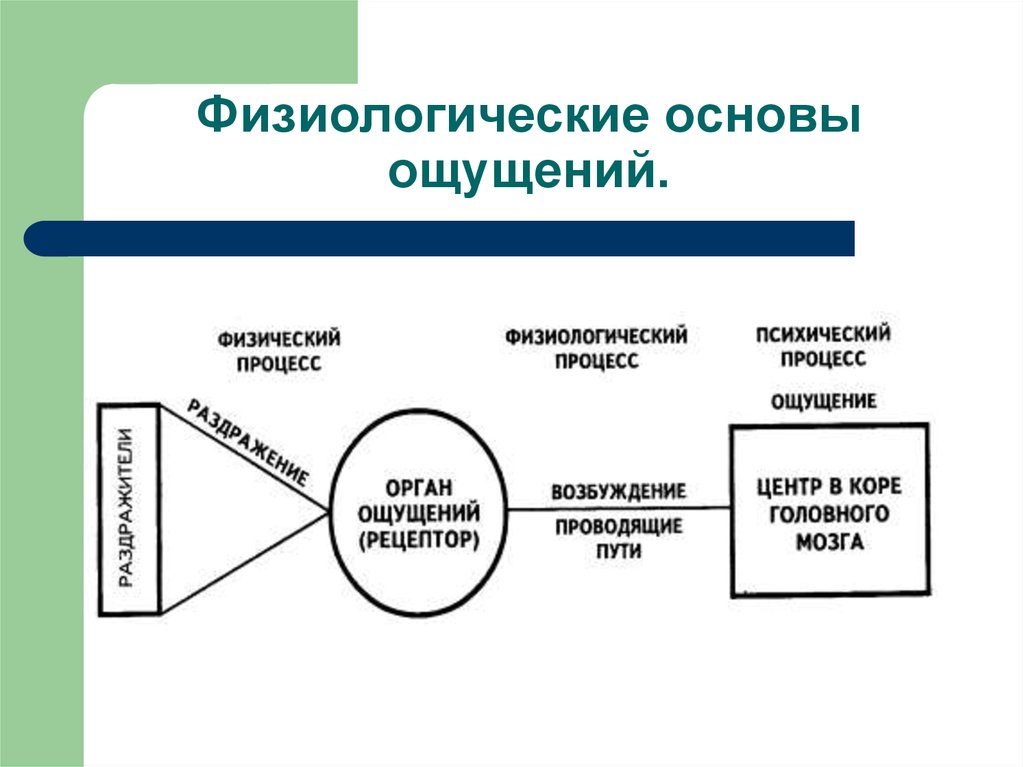 д. являются категорией сознательных форм в что мы ценим что-то об энергозависимых атрибутах объектов. Эта категория сенсорных качеств была центральной темой субдисциплины психологии, называемой психофизикой. Цель психофизика состоит в том, чтобы количественно оценить способность сенсорной системы по-разному реагировать на энергетические переменные, поглощаемые рецепторами, и открыть законы, по которым энергетические различия лежат в основе различий в различных измерениях {29}.} наш сенсорный опыт.
д. являются категорией сознательных форм в что мы ценим что-то об энергозависимых атрибутах объектов. Эта категория сенсорных качеств была центральной темой субдисциплины психологии, называемой психофизикой. Цель психофизика состоит в том, чтобы количественно оценить способность сенсорной системы по-разному реагировать на энергетические переменные, поглощаемые рецепторами, и открыть законы, по которым энергетические различия лежат в основе различий в различных измерениях {29}.} наш сенсорный опыт.
Психофизика установила, например, что при достаточно длительном воздействии стимулов при неизменности всех остальных факторов и изменении только интенсивности источника видимого света изменения воспринимаемой «яркости» происходят в том же направлении, что и изменения интенсивности. Увеличение интенсивности приводит к увеличению яркости; уменьшение интенсивности приводит к уменьшению яркости. Было обнаружено, что аналогичные отношения сохраняются и для других сенсорных качеств, таких как, например, между увеличением звукового давления и слышимой громкостью; повышение температуры и ощущение тепла; повышение концентрации сахара в растворе и вкусовой сладости; и так далее. Кроме того, психофизика установила, что непрерывные качественные изменения приложенной энергии вызывают непрерывные качественные изменения в соответствующем сенсорном опыте. Например, последовательность дискретных световых импульсов достаточной продолжительности и внутриимпульсного разделения, расположенных в последовательном порядке увеличения длины волны, дает последовательность сенсорных переживаний дискретных цветовых импульсов, которые можно расположить в том же порядке с точки зрения длины волны. эмпирическое измерение «подобия оттенка». Таким образом, если бы испытуемого попросили расположить цветные огни в порядке их воспринимаемого сходства, порядок был бы таким же, как у физиков, упорядочивающих огни в соответствии с увеличением длины волны. Было обнаружено, что аналогичные отношения существуют между акустической частотой и воспринимаемой высотой звука. Из этих наблюдений ясно, что сенсорная система дает отдельные переживания для отдельных форм энергии и что эти переживания обладают размерами, параллельными измерениям зарегистрированной формы энергии. Различия в качестве энергии, например в частоте, приводят к {30} различиям в сенсорном качестве, например в цвете. Таким образом, мы можем заключить, что наши сенсорные качества представляют собой формы, в которых выделяются и сознательно сохраняются определенные атрибуты энергетических паттернов. Чтобы понять значение этих фактов для восприятия, необходимо определить: (1) Каковы отдельные черты энергетических паттернов, которые определяют атрибуты наших сенсорных качеств? и (2) Как выделение этих конкретных энергетических характеристик делает возможным наше восприятие объектов? Другими словами, какова функциональная ценность для восприятия объекта физиологических принципов, лежащих в основе выделения энергетических атрибутов для получения сенсорных качеств?
Сенсорная регистрация энергетических характеристик
Чтобы ответить на наш первый вопрос, недостаточно знать, например, что яркость — это сознательная форма, в которой мы оценили интенсивность источника света. Ибо этот факт не говорит нам, какова интенсивность источника света, которую мы оцениваем, когда осознаем его яркость. Чтобы ответить на этот вопрос, мы должны знать, как интенсивность регистрируется как яркость. {31}
Рисунок 3.
Сенсор Регистрация величины энергии : Схема трех основных способов регистрации величины энергии: регистрация в терминах абсолютных местных величин; Регистрация по разнице местных величин; и регистрация в терминах соотношений местных величин. (См. текст на стр. 32.)
В общем, существует три основных способа регистрации атрибутов энергии: «Абсолютная» регистрация, «Разностная» регистрация и «Относительная» регистрация. (См. рис. 3.) Абсолютный регистр — это регистр, который всегда дает одно и то же указание или выходной сигнал для одной и той же абсолютной энергии и выходной сигнал которого прямо пропорционален абсолютной величине падающей энергии.
Разностный регистр — это регистр, который дает одинаковые показания или выходные данные для одной и той же разности энергий между любыми двумя точками и чьи выходные данные прямо пропорциональны разнице в величине между падающими энергиями в двух точках поглощения. {32}
Регистр отношения — это регистр, который дает одинаковый результат для одного и того же соотношения энергий между любыми двумя точками и выход которого пропорционален отношению величин энергии в двух точках поглощения. Основным ограничивающим фактором для всех регистров энергии является количество энергии, которое они могут выдержать без повреждения или перегрузки. Элементы, поглощающие энергию, могут быть защищены от энергетической перегрузки с помощью механизма, который автоматически ослабляет интенсивную энергию до того, как она достигнет поглощающих элементов или до того, как ее воздействие достигнет регистрирующего механизма. Однако любой ослабляющий механизм должен изменять показания индикатора, полученные от респондентов абсолютной энергии и от респондентов разности.
На рис. 3 этот факт схематично проиллюстрирован. Обратите внимание, что когда два источника энергии, a и b, ослабляются, чтобы достичь поглощающих элементов со значениями энергии, уменьшенными до a’ и b’, реагирующие устройства с абсолютной энергией и реагирующие устройства с разностной энергией будут давать разные реакции на условия без ослабления по сравнению с условиями ослабления. Но реагирующий на отношение не изменит свою реакцию. Для респондента отношения оба условия дают одинаковый результат. Таким образом, датчики отношения являются единственной категорией регистров энергии, которые позволяют затухать общую энергию, не влияя на их выход. Конечно, реагирующие на соотношение расплачиваются за это преимущество тем, что не могут предоставить информацию, касающуюся абсолютной энергии или разницы энергий в их рецепторных элементах.
Обратимся теперь к психологическим данным, чтобы определить, какую категорию энергетического отклика представляют наши сенсорные переживания. Наблюдения за сенсорным опытом, вызванным одиночными стимулами, недостаточно, чтобы определить, как наши чувства регистрируют величины энергии. Мы должны использовать несколько стимулов и определить эмпирические эффекты каждого из этих {33} стимулов, когда они предъявляются наблюдателю одновременно.
Психофизические данные с использованием множественных стимулов убедительно свидетельствуют о том, что наши сенсорные системы в значительной степени реагируют на отношение. Рассмотрим, например, классические демонстрации Гелба (сообщенные в Ellis, 19).39), на котором в дверном проеме, соединяющем две затемненные комнаты, подвешен плоский черный диск. Диск освещается источником белого света, невидимым для наблюдателя, находящегося в одной из комнат лицом к черному диску. Наблюдатель сообщает, что видит ярко-белый диск на темном фоне соседней комнаты. Затем на свет выносится небольшой белый лист бумаги и прикладывается к черному диску. Мгновенно субъект сообщает, что диск теперь стал черным, а лист бумаги кажется белым. Этот эксперимент демонстрирует, что наше восприятие объектов как светлых или темных не представляет собой сенсорную регистрацию локальной абсолютной энергии, поскольку наше восприятие яркости одной и той же локальной области может измениться, когда в этой области не происходит никаких изменений энергии. Наше восприятие яркости объектов зависит некоторым образом от энергии, поступающей к нам от объекта, а также от энергии, поступающей к нам от окружающих объектов или фона. Ставшие уже классическими эксперименты Уоллаха (1948) являются результатом попытки выявить точное соотношение между стимулом и окружением, определяющее воспринимаемую яркость каждого из них. В этих экспериментах использовались два диска и два подвеса. Интенсивность света на любом из дисков или на окружении регулировалась независимо. Когда первый диск/объемный дисплей, который служил эталоном, был настроен на любое значение освещенности диска и другое значение объемного освещения, а объемное освещение тестового {34} дисплея было установлено на некоторое значение, оно отличается от ISS, испытуемые, когда их просили сопоставить тестовый диск с эталонным диском, регулируя освещенность первого, последовательно регулировали освещенность тестового диска так, чтобы I сд /И сс = л тд /И тс (приблизительно). Уоллах заключает из этих экспериментов с тем, что он называет «постоянством яркости и ахроматическими цветами», что цвета, возникающие в этих условиях, в близком приближении зависят от соотношений вовлеченных интенсивностей и кажутся независимыми от абсолютной интенсивности локальной стимуляции. Область с более высокой интенсивностью будет иметь белый цвет, а область с более низкой интенсивностью — серый (или черный), что зависит от соотношения интенсивностей двух областей. (стр. 241)
На аналогичную зависимость между световым составом объекта или зрительной области и составом окружающих областей указывает известное явление «одновременного цветового контраста». Это явление показывает, что восприятие цвета, по крайней мере частично, определяется составом длин волн окружения или фона. В качестве примера этого явления, если красное и белое круглые световые пятна спроецировать на экран так, что эти два пятна частично перекрываются, область перекрытия будет восприниматься как белая, физически белый I полумесяц, т. е. полумесяц. получающий свет только от белого проектора, будет восприниматься как сине-зеленый, а красный полумесяц будет восприниматься как красный. Если синее пятно заменяет красное, восприятие трех областей будет: а) белым, где точки пересекаются, б) желтым, когда на экран попадает только физически белый свет, и в) синим, где достигает только синий свет. экран. Как правило, серповидная область белого света воспринимается как дополнительный цвет к цвету другого проектора, а область перекрытия воспринимается как белая. Кроме того, насыщенность воспринимаемого цвета в области, получающей только белый свет, пропорциональна интенсивности света, попадающего на экран от цветного проектора, и не зависит от изменений общей интенсивности.
Эксперименты Эдвина Лэнда (1959, 1965) по восприятию цвета позволяют предположить, что воспринимаемый цвет зависит, по крайней мере частично, от совместных эффектов масштабирования освещенности с точки зрения отношения локальной энергии к фоновой энергии и к общему освещению, переносимому двумя или более рецепторными системами, каждая из которых избирательно реагирует на разные (но перекрывающиеся) области видимого спектра. Лэнд продемонстрировал, например, что смешивание белого света с монохроматическим светом таким образом, что на отображаемой проекции создается узор из различных соотношений или комбинаций смесей белого и монохроматического света, приводит к восприятию широкого спектра цветов, большинство из которых не может быть получен стандартными процедурами изолированных цветовых смесей белого и монохроматического света. Кроме того, цвета, полученные Лэндом, оставались стабильными, когда общая энергия проекторов увеличивалась или уменьшалась. Изменение общего освещения не влияет на локальные соотношения частот.
Реакция отношения и восприятие объекта
Отвечая на второй из вопросов, поднятых в начале этого раздела, ценность реакции отношения заключается в том, что отношения энергий говорят нам об объектах больше, чем энергетические различия. Эти последние, с другой стороны, говорят нам больше об энергии, чем об объектах. Чтобы понять, что это значит, мы должны вспомнить, что объект — это пространственно ограниченная, когерентная, обособленная «вещь». Способ, которым мы воспринимаем объект, заключается в регистрации того факта, что реакция на энергию в области пространства, занимаемой объектом, отличается от реакции на энергию области, окружающей объект. Границы объекта определяют области, в которых реакции на энергию {36} различны. Эти границы объекта создают запас энергии в окружающем свете (Gibson, 19). 66), и эти запасы энергии отображаются в виде демаркационных линий, определяющих паттерны градиентов энергии, распределенных по сетчатке.
Именно структурные различия между объектами объясняют их соответствующие реакции на энергию. Эти структурные различия не вызваны энергией; они энергонезависимы. Как я уже говорил ранее, структура, форма и пространственные отношения объекта с другими объектами являются энергетически независимыми атрибутами объекта.
Имея в виду эти факты, мы теперь можем понять, почему сенсорная система, которая регистрирует разницу в степени, в которой объект отражает или поглощает энергию, по сравнению со степенью, в которой фон отражает или поглощает энергию, независимо от фактической интенсивности общая падающая энергия превосходит сенсорную систему, которая регистрирует различия в степени энергии, отраженной или поглощенной объектом, по сравнению с отражением или поглощением фоном. Для первой (относительно отзывчивой) системы границы объектов будут регистрироваться одинаково хорошо, т. е. с одинаковой резкостью, как в условиях интенсивного, так и слабого освещения. Для последней (реагирующей на разницу) системы границы объектов регистрируются с уменьшением резкости по мере уменьшения освещенности. Пороги отношений, действующие в первой системе, позволяют регистрировать границы объектов в гораздо более широком диапазоне преобладающих энергетических условий, чем это допускается в системах, управляемых разностными порогами, действующими во второй системе.
Подпороговые и надпороговые энергетические отношения , будучи независимыми от падающей энергоемкости, говорят нам кое-что о самих объектах. {37} Подпороговые и надпороговые энергетические различия , будучи энергозависимыми, могут больше сказать о преобладающем энергетическом уровне, чем об объектах. Для сенсорной системы, работающей за счет разностной реакции, границы объектов будут попеременно появляться и исчезать по мере увеличения и уменьшения общего освещения. Но для сенсорных систем, работающих в терминах реакции отношения, границы объектов остаются очень стабильными независимо от изменений освещения. По этой причине относительный ответ предпочтительнее разностного отклика для оценки границ объекта . И наоборот, разностный отклик предпочтительнее, чем относительный отклик, для оценки границ энергии .
Поскольку оптимальное определение границ объекта необходимо для оптимального восприятия объекта, для зрения наилучшим способом оптимизации определения границ объекта является масштабирование яркости с точки зрения отношения энергии.
Ценность и значение постоянства цвета.
«Хроматическое и ахроматическое масштабирование яркости стало возможным благодаря энергозависимым свойствам объектов, а именно, их отражательной способности поверхности и пигментации. Но ценность масштабирования легкости заключается в том, что оно позволяет регистрировать энергонезависимые атрибуты объекта, а именно границы объекта. Тот факт, что оцениваются также энергозависимые свойства объектов, не связан с восприятием объектов. Важным в объектах для восприятия является не их локальный цвет, а их цвет, контрастирующий с их окружением; не их легкость, а их отражательная способность контрастирует с их окружением. Тот факт, что воспринимаемые цвета и яркость объектов остаются относительно постоянными независимо от изменений общего освещения, является отдельным и второстепенным вопросом. Что первично, так это регистрация границ объектов в терминах 9; различия яркости. В своей статье о постоянстве цвета Хелсон (1943) определяет {38} фундаментальную важность различий яркости для восприятия контуров и границ:
Контрастные градиенты, особенно различия яркостей, устанавливают границы между объектами и их окружением. Чтобы видеть объекты в третьем измерении, требуется разделение плоскостей и формирование границ. Градиенты помогают отделить объекты от окружения
. . . (стр. 262)
. . .
Представление легкости служит для того, чтобы отделить объект от его окружения и сделать его вещью в той же мере, в какой оно стремится к постоянству цвета. (стр. 26b)
На самом деле, весь вопрос о постоянстве цвета — сплошная путаница. Сами термины «постоянство цвета» и «постоянство светлоты» вводят в заблуждение. Эти термины обычно подразумевают, что наши чувства склонны сохранять «истинные» или постоянные локальные цвета или яркость объектов. Но это неверно. Во-первых, явление легкости и цвета контраст показывают, что воспринимаемая яркость и цвет объекта определенно изменяются в зависимости от окружающего контекста, в котором представлены объекты. Во-вторых, фотореактивные свойства объектов, такие как их абсолютная локальная ахроматическая или хроматическая отражательная способность, не поддаются непосредственному зрительному восприятию и не являются соответствующими светлоте и цвету соответственно. С другой стороны, верно, что если состав длин волн источника освещения поддерживается постоянным, а источник изменяется только по интенсивности, то относительная яркость и цвет останутся постоянными. Но это не отражает действие сенсорного механизма, который имеет тенденцию удерживать постоянными локальных значений цвета и яркости. Цвета и светлоты воспринимаются как постоянные, потому что эти чувственные качества определяются энергетическими соотношениями, которые при данных условиях изменения интенсивности источника освещения являются постоянными. {39}
Существует очень тонкое заблуждение в понятиях легкости и постоянства цвета. Эти термины являются продуктами теории зрения, которая утверждает, что восприятие света и цвета — это формы, в которых мы непосредственно оцениваем локальные факты о зависящих от света свойствах объектов. На самом деле, однако, цвет и светлота — это формы, в которых мы воспринимаем отношения между зависящими от света атрибутами объекта.0011 объект по сравнению со светозависимыми атрибутами фона , в котором этот объект оказывается обнаруженным. Следовательно, эта взаимосвязь меняется по мере изменения фона объектов, что приводит к восприятию разных цветов объектов в разных фоновых условиях. В любом случае, наше восприятие цвета объекта в равной степени достоверно, поскольку то, что мы воспринимаем в виде изменения локальных цветов, есть изменения светореактивного отношения объект/фон, которым, в конце концов, соответствует воспринимаемый цвет.
Проблема разрешения кажущегося противоречия между фактами постоянства цвета и фактами цветового контраста возникает только в том случае, если мы предположим, что яркость соответствует локальной силе света, а цвет соответствует локальному составу длин волн, и если мы предположим, что «яркость ” соответствует местной отражательной способности. Эти предположения вынуждают нас интерпретировать данные о восприятии цвета и яркости в том смысле, что наша зрительная система способна преодолевать изменения в освещении, чтобы предоставить нам точную информацию о локальных яркостях объектов, достижение, называемое «постоянством яркости», и что наша зрительная система неспособность преодолеть влияние на воспринимаемую яркость, вызванное различиями в фоновом или окружающем освещении, искажение , называемое «одновременным яркостным контрастом». Но объективность восприятия может быть вновь подтверждена {40}, если мы откажемся от этой интерпретации данных, отказавшись от необоснованных предположений о корреспондентах зрительного восприятия, на которых эта интерпретация основана, и признаем тот факт, что и яркость, и цвет соответствуют сложным отношения между фотореактивными свойствами объектов и фотореактивными свойствами окружающих объектов.
Если мы хотим понять восприятие, мы не должны начинать с каких-либо предубеждений о том, какими должны быть соответствия сенсорных качеств . Мы должны эмпирически выяснить, что представляют собой эти корреспонденты . Если мы совершим ошибку, приняв первый подход, т. е. предвосхищая то, чему должны соответствовать наши чувственные качества, когда мы обнаружим, что наши чувства не могут надежно регистрировать то, что они «должны» регистрировать, мы можем совершить необоснованный и эпистемологически самостоятельный шаг. противоречивый вывод о том, что наши чувства ненадежны и подвержены всевозможным «иллюзиям». По этому губительному пути пошло большинство писателей о чувственном восприятии.
Четкие энергетические границы как предпосылка для восприятия объектов отображаемые градиенты энергии и определяющие внешние границы объекта. Объекты не просто пространственно ограничены; они, по большей части, резко очерчены своими внешними поверхностями. Именно на поверхности предметов организмы взаимодействуют с предметами. Кроме того, объекты с эфирными или туманными краями экологически редки (например, облака или туман) и пространственно нестабильны.
Именно по этим причинам система восприятия {41} превратилась в весьма избирательный пограничный регистр. Поскольку границы объектов задаются у рецептора в терминах соотношений между энергетическими границами, восприятие объектов требует регистрации энергетических отношений, складывающихся на четко очерченных энергетических границах. Расплывчатые или постепенные границы порождают нестабильное восприятие формы или, если они достаточно постепенные, вообще не воспринимают формы. Ниже приведены два примера применения этого принципа в визуальной области: Первый пример состоит из эксперимента, который читатель может легко провести. Поместите карандаш между большим рассеянным источником света и плоской поверхностью так, чтобы на поверхность отбрасывалась расплывчатая тень карандаша. Теперь зафиксируйте центр этой расплывчатой тени и обратите внимание, как быстро она исчезает. Это исчезновение сопровождается небольшим уменьшением кажущейся яркости поверхности, что, возможно, связано с каким-то усреднением энергии зрительной системой. Тень можно восстановить, увеличив резкость градиента яркости на сетчатке. Этого можно добиться, не изменяя крутизны градиента энергии на затененной поверхности, быстрым перемещением карандаша вбок, т.е. под прямым углом к линии, соединяющей источник света и поверхность, или перемещением глаза так, чтобы затененная область сместилась по сетчатке. Любое из этих движений вызывает увеличение различий в скорости поглощения энергии в соседних областях сетчатки, тем самым восстанавливая градиент адекватной резкости для границы и, следовательно, для восприятия формы.
Мой второй пример относится к работе Лэнда по восприятию цвета. Вы помните, что Лэнд (1959) смог воспроизвести восприятие большого количества цветов, используя только красный и белый свет, смешанные в разных соотношениях на освещенном экране. Это было достигнуто с помощью двух черно-белых прозрачных пленок сцены, сфотографированных один раз через красный фильтр {42} и один раз через зеленый фильтр. Два получившихся негатива были преобразованы в две позитивные диапозитивы, названные соответственно 9. 0011 длинная и короткая запись. Красный свет проецировался через длинную пластинку, а белый свет проецировался через короткую пластинку. Когда два полученных изображения были наложены на проекционный экран, получился многоцветный массив, содержащий те же цвета, что и исходная сцена, но со значительной потерей насыщенности.
После длительных экспериментов с различными типами черно-белой пленки мне удалось подготовить набор из длинных и — короткие записи , которые довольно хорошо воспроизводят результаты Лэнда.* Я смог воспринимать, используя только красный и белый свет; красный, бледно-голубой, синий, зеленый, бледно-желтый, оранжевый, коричневый, черный, серый и белый. Они соответствовали цветам исходной сцены, но со значительным обесцвечиванием, особенно для синего и желтого.
Суть нашего последнего примера, однако, касается второго эксперимента Лэнда, описанного в той же статье. В этом эксперименте Лэнд попытался воспроизвести все возможные соотношения красного и белого света, заменив две прозрачные пленки двумя клиньями нейтральной плотности, ориентированными под прямым углом друг к другу. Таким образом, интенсивность белого света непрерывно уменьшалась от верхней части экрана к нижней, тогда как интенсивность красного света непрерывно уменьшалась слева направо на экране. Лэнд описывает результаты следующим образом:
Когда оба проектора включены, мы теперь имеем бесконечное разнообразие соотношений красного и белого на экране, дублируя все те, которые могут иметь место в цветном изображении. Однако они расположены в строго упорядоченной последовательности. Случайности нет. А на экране нет цвета только градуированная розовая заливка. (p93)
* Я хотел бы поблагодарить г-на Джея Конна за его бесценные предложения и за его фотографическую работу при подготовке слайдов. {43}
Вывод Лэнда о том, что цвета могут быть восприняты только тогда, когда хроматические соотношения изменяются случайным образом от точки к точке, не совсем достаточен для объяснения отсутствия цвета в этих условиях постоянно меняющихся соотношений. Лэнд все еще пытался разобраться с восприятием цвета в терминах точечного или локального анализа частотного состава. Тот факт, что для восприятия цвета требуется то, что Лэнд называет случайным распределением локальных соотношений, гораздо более понятен, если его описать в терминах гипотезы, которую я здесь отстаиваю, а именно (с чем может согласиться Гибсон), что для восприятия цвета требуются четкие границы хроматической энергии, т. е. , границы, за которыми резко меняются хроматические соотношения.
Еще один драматический пример важности четких энергетических границ для восприятия цветовых различий можно найти в экспериментах Ганцфельда, проведенных Хохбергом, Трибелем и Симаном (1951). Ганцфельд состоит из однородного бестекстурного поля зрения, обычно получаемого экспериментально путем равномерного освещения белой бесфактурной поверхности, занимающей все поле зрения. Хохберг и его коллеги использовали половинки мячей для настольного тенниса, удобно, но прочно приклеенные к глазам и освещенные снаружи. Два общих вывода экспериментов Ганцфельда имеют отношение к нашему настоящему обсуждению.
Первое важное открытие заключается в том, что при полном отсутствии визуальной текстуры, т. е. визуальных контуров или граничных паттернов, исчезает всякое восприятие цвета. Если, например, поле освещается красным или зеленым светом, испытуемые сообщают, что восприятие красного или зеленого происходит только в момент включения освещения, после чего цвет постепенно исчезает. Когда цвет исчез, введение визуального градиента, созданного путем отбрасывания тени от пальца экспериментатора {44} на центральную область глазного яблока, дало следующие результаты:
В группе A1, в которой падающий свет был красным, все пять S сообщили о черной тени, окруженной красным ореолом, и тень, и ореол быстро исчезли после удаления пальца. В группе A2, в которой падающий свет был зеленым, четыре из пяти S сообщили о черной тени на зеленом фоне или ореоле. Остальные S сообщили о пурпурной тени на фоне зеленого ореола. (Hochberg, Triebel, and Seaman, 1951, стр. 66)
Эти результаты показывают, что восприятие цвета может поддерживаться только при наличии градиентов энергии в поле зрения. Кроме того, два факта, полученные в этих экспериментах, по-видимому, противоречат указанию на то, что исчезновение цвета является пассивным результатом усталости сетчатки от монохроматического света. Во-первых, введение тени вызывает немедленное возвращение цвета в область 9.0011 окружает тень, хотя энергия падающего света, падающего на области сетчатки, окружающие изображение тени на сетчатке, не меняется. Во-вторых, девять из десяти испытуемых сообщили, что, когда диффузное освещение прерывалось на интервалы примерно в 2 секунды, они мгновенно ощущали комплементарный цвет, хотя следует предположить, что стимуляция комплементарных рецепторов сетчатки была либо неизмененной , либо, в лучшем случае чуть уменьшил , прервав подсветку. В любом случае эффект прерывания освещения должен был вызвать, согласно простой гипотезе о локальном утомлении или возбуждении сетчатки, либо отсутствие восприятия дополнительного цвета, либо, в лучшем случае, снижение восприятия дополнительного цвета. Мы можем заключить, что в отсутствие энергетических паттернов хроматические свойства световой энергии недостаточны для устойчивого восприятия цвета.
Второе крупное важное открытие, сделанное Хохбергом и Коэном в исследованиях Ганцфельда (1957) заключается в том, что при полном отсутствии визуальной текстуры восприятие радикально нарушается до такой степени, что некоторые испытуемые {45} сообщают о сильном беспокойстве и страхе «ослепнуть» (Hochberg et al.). Об этом феномене Коэн сообщает:
В ходе адаптации пять из шестнадцати наблюдателей сообщили о полном прекращении зрительного опыта. Двое сообщали о потере сознания почти каждый раз, когда они сталкивались с длительной гомогенной стимуляцией. темное, недифференцированное поле зрения. Характерно следующее описание: «туманная белизна, все затемняется, возвращается, уходит. Я чувствую себя слепым. Я даже черноты не вижу. Это отличается от черного цвета при выключенном свете». Можно предположить, что перцептивный механизм развился, чтобы справляться с дифференцированным полем, и при отсутствии дифференцирования происходит временный сбой механизма… (стр.
Знак восприятия Значение сенсорной адаптацииСенсорная адаптация — это общий термин, относящийся к прогрессивному и обратимому изменению реакции сенсорной системы на компоненты доступной энергии. Под сенсорной адаптацией я подразумеваю физиологическую адаптацию к энергетическим компонентам и не включаю перцептивные адаптации, такие как те, которые достигаются при ношении инвертирующих или смещающих призм, которые включают сенсомоторное обучение .
Простым примером того, как адекватный стимул становится неадекватным, является адаптация к однородному цвету в экспериментах Ганцфельда. Здесь присутствие монохроматического света постепенно перестает вызывать восприятие цветной поверхности и вместо этого вызывает либо ощущение неопределенного тумана, либо вообще не вызывает визуального осознания.
Другим примером в этой категории является адаптация к давлению на кожу, когда вес, помещенный на кожу, постепенно становится незаметным. Оба этих примера следует рассматривать как случаи стимул отказ, а не нейронный отказ.
Их результаты показывают, что груз, однажды помещенный на поверхность кожи, не остается на ней, а продолжает двигаться вниз. на удивление долго. Давление ощущается только до тех пор, пока сохраняется запредельная скорость движения. Когда сопротивление ткани уменьшает движение до неопределяемого уровня, ощущение исчезает. Конечная точка «адаптации» достигнута. Вывод очевиден: «полная адаптация» означает отказ от стимула. Тот факт, что ответ не следует искать в «усталости» или истощении ресурсов органа-мишени, легко доказывается увеличением нагрузки на ткань после того, как адаптация, по-видимому, исчерпала себя. В такой ситуации возникает свежая реакция, новое ощущение, сопровождающее дальнейшее сдавливание, пока раздражитель снова не «отказывает».
Эти два примера сенсорной адаптации на самом деле представляют собой последствия отсутствия стимула и предполагают, что минимальные условия для восприятия требуют большего, чем просто отображение энергии на рецепторе, и больше, чем отображение резкого градиента энергии. у рецептора. Что требуется, так это изменяющееся распределение энергии, то есть отображение энергии, которое меняется на рецепторе с течением времени. Это требование восприятия ярко иллюстрируется в области зрения экспериментальными исследованиями фиксированных изображений на сетчатке. Эта совокупность данных, рассмотренная Хеккенмюллером (1965), связан с перцептивным исчезновением объекта, оптическое изображение которого отображается на сетчатке таким образом, что положение изображения относительно сетчатки остается фиксированным независимо от того, как глаза двигаются относительно объекта. В этом случае восприятие объекта нарушается, даже если на рецепторе существует четко определенный паттерн энергии.
Независимо от того, каковы конкретные неврологические механизмы, общий принцип, предложенный экспериментами с неподвижными изображениями сетчатки, Ганцфельдом, цветовым и яркостным контрастом и кожной чувствительностью к давлению, заключается в том, что адекватная сенсорная стимуляция требует постоянно меняющихся проявлений энергетических паттернов.
Философская записка об эпистемологическом значении отказа от стимула
Требования к адекватной сенсорной стимуляции, т. е. стимуляции, которая приведет к устойчивому восприятию, можно рассматривать как условия, заложенные в перцептивных системах для того, чтобы гарантировать, насколько это физиологически возможно, что восприятие вызвано экзогенными паттернами нервного возбуждения и не зависит от эндогенных паттернов возбуждения.
Обратимся теперь к категории сенсорной адаптации, при которой ранее неадекватный раздражитель становится адекватным. Из данных об этой категории приспособления, по-видимому, вытекает другой принцип сенсорного функционирования, а именно. что наши сенсорные системы постоянно приспосабливают свои ответы к компонентам энергетических градиентов, чтобы максимизировать диапазон перцептивных дифференциаций, возможных при данных условиях стимуляции.
Рассмотрим, например, классический феномен цветовой адаптации. Если поле зрения заполнено светом одного цвета или если сегмент видимого спектра отфильтрован, например, при ношении цветных очков, первоначальный перцептивный эффект — резкое уменьшение диапазона различимых цветов.
Рисунок 4.
Схема гипотетического случая цветовой адаптации отношения различий хроматических откликов в областях A и B, а именно, отношение различий между красным и зеленым откликом в A, выраженное как R A -G A , и красным и зеленым откликом в B , выраженный как R B -G B , то уменьшение общего количества зеленого света путем фильтрации уменьшит коэффициент разности и, таким образом, уменьшит хроматический контраст.
Цветовая адаптация, как и адаптации, приводящие к отказу от стимула, является активным сенсорным механизмом для обеспечения оптимальной адекватной сенсорной стимуляции, а не просто пассивным результатом рецепторного «усталости» или «истощения» красных (или зеленых) рецепторов . То, что последнее не является адекватным объяснением цветовой адаптации, ярко иллюстрируется экспериментами Колера (Kohler, 1962) с цветными очками. Испытуемые Келера в течение длительного периода времени носили очки, каждый монокль которых был разделен пополам по вертикали, так что левое поле было окрашено в желтый цвет, а правое — в синий.
Из экспериментальных данных, оцененных в контексте концепции восприятия, предложенной в этой статье, можно сделать несколько общих выводов:
1.
2. Наши сенсорные системы выполняют свою функцию выделения и регистрации энергетических атрибутов объекта путем выделения атрибутов изменяющихся градиентов энергии, которые независимы от энергии . Например, наши сенсорные системы склонны выделять энергетические отношения , поскольку они относительно независимы от изменений общей энергии. Наши сенсорные системы выделяют острые энергетические границы , так как они также не зависят от изменений общей энергии. Наши сенсорные системы выделяют систематические изменения в направленность градиентов энергии, отображаемых на приемнике, поскольку направленность также не зависит от изменений общей энергии.
3. Система восприятия специально использует принцип транспозиции паттернов, чтобы изолировать паттерны возбуждения {53}, вызванные извне. Возбуждающий эффект, производимый паттерном, содержит признаки, инвариантные по отношению к трансляции паттерна на нейронном субстрате. Именно эти реляционные инварианты, а не сами нервные элементы составляют сенсорные единицы. Наше восприятие даже «самых простых» сенсорных фактов, таких как различия в освещенности, должно зависеть: от создания нейронных паттернов, содержащих реляционные признаки, которые инвариантны по отношению к трансляции паттерна или градиента стимуляции.
До сих пор мы обсуждали некоторые чисто физиологические механизмы, ответственные за изоляцию и извлечение объектно-специфических энергетических эффектов. Мы видели, что сенсорная система имеет встроенные процессы, которые автоматически преодолевают последствия неинформативных энергетических событий и эндогенных физиологических нарушений. Эти результаты достигаются благодаря тому, что сенсорная система имеет дело с реляционными свойствами изменяющихся энергетических паттернов. Хотя мы определили, каковы общие физические и физиологические предпосылки восприятия, их самих по себе недостаточно, чтобы объяснить восприятие как сознательное осознание объектов, внешних по отношению к воспринимающему и независимых от него. Восприятие не является пассивным продуктом физических и физиологических событий, но его полное функциональное осуществление зависит от взаимодействия между сознательных процессов и автоматических физиологических процессов.
Последняя редакция страницы: 19 апреля 2014 г. 15:58
Нравится:
Нравится Загрузка…
Психология 159: Физиологические основы восприятия
Психика 159: Физиологические основы восприятияИнструктор: Дон Маклауд
Классные собрания: лекции, вторник и четверг с 12:30 до 1:50. вечера, 1507 Мандлер
Часы работы: Среда, 10-12, комната 5121 McGill; Тел. 534-3975.
Электронная почта: dmacleod@ucsd.edu
Интернет: http://pages.ucsd.edu/~dmacleod/ (и нажмите на ссылку «курсы»)
Заключительные вопросы (с прошлого года): здесь
Цели и область применения: Мы рассматриваем процесс восприятия как причинно-следственная цепь, начинающаяся с сенсорный стимул и продолжается через последовательные стадии нейронной представление, завершающееся, наконец, построением поведенчески полезное представление об окружающей среде.
Формат: Это занятие в стиле семинара с активной участие поощряется/требуется относительно небольшим размером группа. Текстовая глава или другое чтение для каждой встречи должно быть сделано заранее . В классе большую часть времени будет групповое обсуждение по прочитанному. Дискуссия будет оформлена как лекции (в основном мои), так и презентации студентов.
Текст: ‘ Зрение: вычислительный подход к Биологическое зрение», Фрисби и Стоун. Держите Тексты Psych 102 в качестве фона; особенно полезен Sensation & Восприятие Джереми Вулфа и других. (Sinauer, 2006), но и другие книги «Введение в ощущение и восприятие», в частности Секулер и Блейк почти так же полезны.
В дополнение к тексту я планирую включить несколько кратких он-лайн чтения , что следует скачать и ознакомиться перед соответствующим собранием учебный класс.
Я хочу, чтобы каждый из вас написал мне по электронной почте до полуночи накануне каждый класс, по крайней мере один вопрос . предмет обсуждения или интересная веб-ссылка, относящаяся к теме дня или вдохновленный назначенным чтением, которое, по вашему мнению, будет полезно для стимулировать или направлять дискуссию. Мы обсудим выбор из эти в классе. Чтобы быть полезными, эти вопросы должны в идеале отражать некоторое вдумчивое внимание к чтению или, возможно, некоторые соответствующие разведка в интернете.
· Пожалуйста, напишите мне свой вопрос, комментарий или веб-информацию до полуночи за день до каждого занятия ; Я сопоставлю их и принесу класс и / или опубликовать их в Интернете.
· Если возможно, включите в свой текст соответствующую цитату. и/или номер страницы из чтения.
· Пожалуйста, укажите Психология 159 в строке темы вашего сообщения.
· Пожалуйста, ограничьте длину своего предложения парой предложений
· Пожалуйста, будьте готовы кратко представить свою точку зрения в классе.
Требования и пожелания: Психология 102, Введение в Восприятие указано как обязательное. Уверенные студенты не исключаются, однако, особенно если они имеют некоторый опыт в нейробиологии или физиологической психологии. По мере прочтения текста шоу, довольно много тем, которые мы рассматриваем, будут рассмотрены количественно , поэтому энтузиазм мыслить количественно, или некоторый опыт в физике или технике, также является отличным преимущество… и отвращение к количественному мышлению является соответствующий гандикап!
Оценка (предварительный план):
Предварительный план состоит в том, чтобы сдать только выпускной экзамен (без промежуточного экзамена).
Бумага будет подана (ориентировочно) в начале занятие в четверг седьмой недели. Рекомендуемый объем — 10 страниц (двойной с интервалом, пожалуйста). Выбор темы остается за вами, при условии, что она соответствующие тематике курса. Сначала обсудите это со мной, если ты неуверен. Оригинальность приветствуется, но не обязательный. Я могу предложить возможные темы, и возможные варианты будут обсуждали в классе. Вы должны выбрать тему и сказать мне свой выбор минимум за неделю до сдачи бумаги. Конечно, я ожидаю, что вы обращаться к исследовательской литературе за пределами рекомендуемой литературы в подготовка бумаги.
· Формат статьи гибкий, но: используйте цитаты в бумаги для обоснования ваших требований.
· Цитируйте авторов по имени и дате, например (Смит и Джонс, 1922 г.).
· Включите полные ссылок в конце статьи (полные цитаты; формат не важен, APA — один из стандартных форматов).
· Включить аннотацию (краткое изложение ключевых моментов, менее 1 страница).
· Вы можете заменить бумагу более конкретным проектом: для например, вы могли бы провести эксперимент по восприятию (чтобы помочь в моей лаборатории возможно), выполните компьютерное моделирование или создайте информативная визуальная демонстрация.
· Пожалуйста, пришлите мне статью по электронной почте, а также в печатном виде. если ты сделать проект, он должен сопровождаться кратким письменным отчетом.
Ваша презентация должна быть короткой и быстрой, но понятное (для ваших одноклассников) изложение какой-либо темы, относящейся к тема дня. Это хорошая идея, чтобы обсудить ваш выбор темы со мной заранее. Один из возможных вариантов (но не лучший один) заключается в том, чтобы просто подытожить и прояснить часть чтения, которое мы занимаясь в день вашей презентации, надеюсь, развивая ваше собственное понимание этой темы, используя другие источники, такие как рекомендуемые чтения.
Рекомендуется использовать немного графики для презентации. PowerPoint — хороший способ. У нас будет компьютерная графика проектор. Если вы можете использовать собственный ноутбук ( с выходом VGA; если у вас нет разъема VGA, пожалуйста, принесите адаптер или позвольте мне знаю ), это будет хорошо. Иначе у меня вообще будет свое. Если вы планируете, у нас есть
Предварительное распределение баллов: Статья/проект/презентация (25% для каждого из двух), финал (30%) и участие в классе (20%).
Выпускной экзамен будет в основном или полностью типа эссе вопросы , возможно также какой-нибудь краткий ответ/множественный выбор вопросы — возможно, соотношение 80/20 между типом эссе и коротким ответить на вопросы в общей оценке.
ССЫЛКА НА ОБСУЖДЕНИЕ И СООТВЕТСТВУЮЩИЕ МАТЕРИАЛЫ
ССЫЛКИ НА ПРЕЗЕНТАЦИИ:
РАСПИСАНИЕ (ТОЛЬКО ЗАПОЛНИТЕЛЬ, ДАТЫ И ЧТЕНИЯ БУДУТ ИЗМЕНЕНО ПОСЛЕ НЕДЕЛИ 1)
Неделя 1 Вторник: Организационное собрание.
Неделя 1 Четверг: Проблема восприятия
Чтение: Глава 1 Фрисби и Стоуна
Chapter1 Видя: Что это? Страница 1
Ch01LectureDon.
Неделя 2, вторник, 7 апреля: Зрение после продолжительной слепоты
Мощность Пункт о Майке May
Прочитайте о нашей работе с Майком Мэем здесь:
http://www.ncbi.nlm.nih.gov/pubmed/12937420
Неделя 2 Четверг, 9 апреля: Эволюция и видение: общая картина
Интерфейсная бумага Хоффмана (локальная копия)
Эволюция глаз
Хоффман Презентация «Теория интерфейса восприятия»
Нильссон и моделирование Пелгером эволюции глаза
Неделя 3….Вторник 14, Четверг 16: Восприятие как обратная задача; использование природных ограничений
Глава 2 Видение формы из Текстуры 29
Неделя 4, вторник, 21 апреля:
Глава 3 Видение с помощью Рецептивные поля 55
Power Point из моей лекции по оптике (подробнее, чем нужно)
Почему зрение не идеальное?
Глава 4 Зрение Последействие: микроэлектрод психолога 75
Неделя 4 Четверг, 23 апреля:
Глава 5 Видение краев 111
Неделя 5, вторник, 28 апреля:
Глава 6 Видение и Сетчатка 133
Сила Пункт из моей лекции о фоторецепторах и зрительной чувствительности
Неделя 5, четверг, 30 апреля:
Глава 7 Видение рисунка с земли 155
Неделя 6, вторник, 5 мая:
Глава 8 Зрение Объекты 173
Неделя 6, четверг, 7 мая:
Глава 9 Видение с помощью Клетки мозга 205
Глава 10 Зрение мозгом Карты 229
Неделя 7, вторник, 12 мая:
Глава 11 Видение и теория сложности 255
Глава 12 Видение и Психофизика 281
Неделя 7 Четверг, 14 мая: ДОКУМЕНТЫ: краткие презентации на основе ваши документы
Неделя 8, вторник, 19 мая: уточняется или перерыв (Дона нет в городе)
Неделя 8, четверг, 21 мая: уточняется
Глава 13 Восприятие как умозаключение 307
Неделя 9, вторник, 26 мая:
Глава 14 Видение движения, часть I 325
Глава 15 Видение движения, часть II 355
Неделя 9, четверг, 28 мая:
Глава 16 Видение черного, серого и Белый 373
Глава 17 Видение цвета 397
Неделя 10, вторник, 2 июня: Stereo Vision, Cue Combination
Глава 18 Взгляд двумя глазами, часть I 419
Глава 19 Взгляд двумя глазами, Часть II 465
Глава 20 Видеть, комбинируя подсказки 497
Неделя 10, четверг, 4 июня: Восприятие как конструктивное процесс; Символическое представление и его вычисление 90 190
Глава 21 Взгляд в блоки Мир 511
Глава 22 Видение и Сознание 527
Глава 23 Обзор видения 539
ФИНАЛ: как в расписании занятий
Дополнительная рекомендуемая литература: общая
Выдающееся введение в физиологические основы слуха является Schnupp et al.
Что касается зрения, если вы хотите преуспеть (а тем более, если вы стремится углубить свое понимание предмета) вы захотите читать дальше. Для начала всем, кто не прошел Психологический 102 может оказаться полезным просмотреть более вводный (Psych 102 или эквивалентный) текст для ознакомления с фоном. Два хороших примера:
Р. Секулер и Р. Блейк, Зрительное восприятие , Кнопф, 1985
Э.Б.Гольдштейна, Ощущение и восприятие , Уодсворт, 1989 г. ( немного более физиологически ориентированный, и поэтому незначительно предпочтительнее здесь, к Секулеру и Блейку).
T.N.Cornsweet, Зрительное восприятие , Academic Press, 1970 г. та редкость: учебник мыслящего человека.
Ричард Грегори, Глаз и мозг , Принстон, 1990 короче, более избирательный, менее техничный и даже более веселый, чем Cornsweet.
С. Э. Палмер, Наука о зрении: от фотонов к феноменологии, Массачусетский технологический институт 1999 г. — это превосходный и всесторонний обзор наших понимание зрительного восприятия, охватывающее феноменологические и познавательной, так и физиологической точек зрения.
Полезно для более продвинутых и современных математически ориентированных обзор большей части материала курса: Wandell B.A. Основы Видение . Sinauer Associates Inc., 1995.
Еще более продвинутый, но полезный для решения текущих вопросов в подготовке статей, являются материалы последних научных симпозиумы: Л. Спиллманн и Дж.С. Вернер (ред.) Зрительное восприятие: Нейрофизиологические основы .
Рекомендуемая литература: специальные темы
Другие книги, рекомендованные для ясного и всестороннего описания часть материала:
ПК Кайзер и Р.М.Бойнтон, Цветовое зрение человека , оптический Society of America, 1996. Здесь обсуждается не только цветовое зрение, но и также зрительная чувствительность (всего около 1/3 курса материал) в достаточно развитой, но понятной форме. Самый полезны главы 1-7 и 10, особенно главы 5, 6 и 7. Для актуального обсуждения актуальных вопросов в цвете зрение вы также можете попробовать R. Mausfeld и D. Heyer, Color Восприятие: разум и физический мир , Оксфорд, 2003.
Д. Марр, Видение , Фримен, 1982 — самая влиятельная книга о видении, написанном в этом столетии, описывающем то, что стало известно как «вычислительный» подход, объединяющий искусственные интеллектуальные и физиологические точки зрения на зрение.
Глава 4 книги Вычислительный мозг, Пэт Черчленд и Терри Сейновски, это хорошее краткое изложение в духе Марра, снова концентрируясь на глубине и трехмерной форме.
С. Зеки, Видение мозга , Блэквелл, 1993 г. красиво иллюстрировано, но особенно однобоко и противоречиво обсуждение особой роли различных областей головного мозга кора в зрении. Более широкий и менее своеобразный, но менее актуальный отчет о физиологических основах зрения с фокус на коре — Д.Х.Хубель, Глаз, мозг и зрение, Фримен, 1988.
Р. В. Родик, Первые шаги к видению , Синауэр, 1998 г. сложен своей строгостью и вниманием к деталям, но он написано ясно, чтобы заинтересованный и вдумчивый читатель мог понять его без особых предварительных знаний. Особое внимание уделяется раннему этапы зрения, в основном глаз и сетчатка. Дополняет Зеки, где основное внимание уделяется коре.
Д.Б. Dusenbery, Сенсорная экология, Freeman 1992 интересный обзор сенсорных процессов у животных.
Дэвид М. Риган, Человеческое восприятие объектов , Синауэр, 2000 подробно обсуждает раннюю визуальную обработку пространственной формы определяется яркостью, цветом, текстурой, движением и бинокулярным несоответствие.
Несколько полезных веб-сайтов ресурсы :
Веб-сайт: Нейронная организация сетчатки позвоночных. актуальный ресурс на сетчатке.
Больше иллюзий от Майкла Бах: http://michaelbach.de/ot/index.html
ViperLib. Обширная и интересная онлайн-коллекция изображений, иллюстрирующих интересные с научной точки зрения моменты о зрении.
Иллюзии Галерея. Сборник зрительных иллюзий с туториалом Информация.
Цвет Системы и изображения Историческая коллекция цветовых систем (Ханс Иртель).
Интерактивный Иллюстрации цвета Восприятие Джон Ф. Хьюз, Джефф Билл и Адам Доппельт, Университет Брауна.
Ощущение и Восприятие Учебники. от Джона Кранца в Ганновере
А коллекция случайных точек одиночного изображения «волшебный глаз» стереограммы
Вичек. Сайт с программой, которая может имитировать, как изображения и веб-страницы будет смотреть на людей с различными типами цветовых недостатков.
Визуальный Экология Веб-сайт ведущей исследовательской группы по таким вещам как дизайн глаз.
Количественный данные о механизмах цветового зрения от профессора Эндрю Стокман, Лондон. В основном полезно для продвинутых читателей, особенно для моделирования реакции глаза на цвет
«Изменить демонстрации слепоты. Они поразительно показывают, как мало мы принимаем то, на что специально не обращаем внимания.
http://auditoryneuroscience.com/ превосходная коллекция слуховых демонстраций от Яна Шнуппа и коллеги.
Физиологические основы ощущений | Измерение ощущений
Фильтр поиска панели навигации Oxford AcademicThe Measurement of SensationNeuropsychologyBooksJournals Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicThe Measurement of SensationNeuropsychologyBooksJournals Термин поиска на микросайте
Расширенный поиск
Иконка Цитировать Цитировать
Разрешения
- Делиться
- Твиттер
- Еще
Cite
Laming, Donald,
‘The physiological basis of sensation’
,
The Measurement of Sensation
, Oxford Psychology Series
(
Oxford,
1997;
online edn,
Oxford Academic
, 1 января 2008 г.
), https://doi.org/10.1093/acprof:oso/9780198523420.003.0006,
, по состоянию на 2 октября 2022 г.
Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Фильтр поиска панели навигации Oxford AcademicThe Measurement of SensationNeuropsychologyBooksJournals Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicThe Measurement of SensationNeuropsychologyBooksJournals Термин поиска на микросайте
Advanced Search
Abstract
В этой главе исследуется гипотеза о том, что степенное преобразование реализуется в элементарной нейронной функции.
Ключевые слова: дифференциальная связь, нейронная функция, степенной закон, экспонента степенного закона, сенсорные иллюзии, изменчивость, достоверные оценки В настоящее время у вас нет доступа к этой главе.
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Нажмите Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- При посещении сайта учреждения используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Войти с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Вход через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic.
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Вход через личный кабинет
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. Смотри ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр учетных записей, вошедших в систему
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Ведение счетов организаций
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т.
Покупка
Наши книги можно приобрести по подписке или приобрести в библиотеках и учреждениях.
Информация о покупке
Сенсорное восприятие – анатомия и физиология
Соматическая нервная система
OpenStaxCollege
Цели обучения
К концу этого раздела вы сможете:
- Описывать различные типы сенсорных рецепторов
- Опишите структуры, ответственные за особые чувства вкуса, обоняния, слуха, равновесия и зрения
- Различать, как передаются разные вкусы
- Опишите средства механорецепции слуха и равновесия
- Перечислите поддерживающие структуры вокруг глаза и опишите строение глазного яблока
- Описать процессы фототрансдукции
Основная роль сенсорных рецепторов заключается в том, чтобы помочь нам узнать об окружающей нас среде или о состоянии нашей внутренней среды.
Описание сенсорной функции термином «ощущение» или «восприятие» является преднамеренным различием. Ощущение – это активация сенсорных рецепторных клеток на уровне раздражителя. Восприятие — это центральная обработка сенсорных стимулов в осмысленный паттерн. Восприятие зависит от ощущений, но не все ощущения воспринимаются. Рецепторы — это клетки или структуры, которые обнаруживают ощущения. Рецепторная клетка изменяется непосредственно под воздействием раздражителя.
Стимулы внешней среды активируют специализированные рецепторные клетки периферической нервной системы. Разные типы стимулов воспринимаются разными типами рецепторных клеток. Рецепторные клетки можно разделить на типы на основе трех различных критериев: тип клетки, положение и функция. Рецепторы можно структурно классифицировать на основе типа клеток и их положения по отношению к воспринимаемым ими раздражителям.
Типы структурных рецепторов
Клетки, которые интерпретируют информацию об окружающей среде, могут быть либо (1) нейронами, имеющими свободные нервные окончания с дендритами, встроенными в ткани, которые будут воспринимать ощущения; (2) нейрон, имеющий инкапсулированное окончание, в котором окончания чувствительных нервов инкапсулированы в соединительную ткань, повышающую их чувствительность; или (3) специализированная рецепторная клетка, имеющая отдельные структурные компоненты, которые интерпретируют определенный тип стимула ([ссылка]). Болевые и температурные рецепторы в дерме кожи являются примерами нейронов со свободными нервными окончаниями. Также в дерме кожи расположены пластинчатые тельца, нейроны с инкапсулированными нервными окончаниями, которые реагируют на давление и прикосновение.
Классификация рецепторов по типу клеток
Типы рецепторов клеток можно классифицировать на основе их структуры. Сенсорные нейроны могут иметь либо (а) свободные нервные окончания, либо (б) инкапсулированные окончания. Фоторецепторы в глазах, такие как палочки, являются примерами (c) специализированных рецепторных клеток. Эти клетки выделяют нейротрансмиттеры на биполярную клетку, которая затем образует синапсы с нейронами зрительного нерва.
Другой способ классификации рецепторов основан на их расположении относительно раздражителей. Экстерорецептор — это рецептор, расположенный рядом с раздражителем во внешней среде, такой как соматосенсорные рецепторы, расположенные в коже. Интерорецептор интерпретирует стимулы от внутренних органов и тканей, таких как рецепторы, воспринимающие повышение кровяного давления в аорте или каротидном синусе.
Функциональные типы рецепторов
Третья классификация рецепторов основана на том, как рецептор преобразует стимулы в изменения мембранного потенциала. Стимулы бывают трех основных типов. Некоторые стимулы представляют собой ионы и макромолекулы, которые воздействуют на белки трансмембранных рецепторов, когда эти химические вещества диффундируют через клеточную мембрану. Некоторые стимулы представляют собой физические изменения в окружающей среде, которые влияют на потенциал мембраны рецепторных клеток. Другие стимулы включают электромагнитное излучение видимого света. Для человека единственной электромагнитной энергией, воспринимаемой нашими глазами, является видимый свет. У некоторых других организмов есть рецепторы, которых нет у людей, например, датчики тепла змей, датчики ультрафиолетового света пчел или магнитные рецепторы у перелетных птиц.
Рецепторные клетки могут быть дополнительно классифицированы на основе типа стимулов, которые они передают. Химические стимулы могут быть интерпретированы хеморецептором, который интерпретирует химические стимулы, такие как вкус или запах объекта. Осморецепторы реагируют на концентрацию растворенных веществ в жидкостях организма. Кроме того, боль — это прежде всего химическое ощущение, которое интерпретирует присутствие химических веществ в результате повреждения тканей или подобных интенсивных раздражителей через ноцицептор. Физические стимулы, такие как давление и вибрация, а также ощущение звука и положения тела (равновесия) интерпретируются через механорецепторы. Другой физический раздражитель, который имеет собственный тип рецепторов, — это температура, которая воспринимается терморецептором, чувствительным либо к температурам выше (тепло), либо ниже (холод) нормальной температуры тела.
Спросите любого, что такое органы чувств, и он, скорее всего, назовет пять основных чувств: вкус, обоняние, осязание, слух и зрение.
В области физиологии чувства можно разделить на общие и специфические. Общее чувство распространено по всему телу и имеет рецепторные клетки в структурах других органов. Примерами этого типа являются механорецепторы в коже, мышцах или стенках кровеносных сосудов. Общие чувства часто вносят вклад в осязание, как описано выше, или в проприоцепцию (движение тела) и кинестезию (движение тела), или в висцеральное чувство, которое наиболее важно для вегетативных функций. Особое чувство — это то, которому посвящен определенный орган, а именно глаз, внутреннее ухо, язык или нос.
Каждое из чувств называется сенсорной модальностью. Модальность относится к способу кодирования информации, что похоже на идею преобразования. Основные сенсорные модальности можно описать на основе того, как каждая из них передается. Химические чувства – это вкус и обоняние. Общее чувство, которое обычно называют осязанием, включает химическое ощущение в форме ноцицепции или боли. Давление, вибрация, растяжение мышц и движение волос под действием внешнего раздражителя воспринимаются механорецепторами. Слух и равновесие также воспринимаются механорецепторами. Наконец, зрение включает в себя активацию фоторецепторов.
Перечисление всех различных сенсорных модальностей, которых может насчитываться до 17, включает в себя разделение пяти основных чувств на более конкретные категории или субмодальности большего чувства. Индивидуальная сенсорная модальность представляет собой ощущение определенного типа раздражителя. Например, общее осязание, известное как соматоощущение, можно разделить на легкое давление, глубокое давление, вибрацию, зуд, боль, температуру или движение волос.
Вкус (вкус)
Лишь несколько признанных субмодальностей существуют в пределах чувства вкуса или вкуса. До недавнего времени различали только четыре вкуса: сладкий, соленый, кислый и горький. Исследования на рубеже 20-го века привели к признанию пятого вкуса, умами, в середине 1980-х годов. Умами — это японское слово, которое означает «восхитительный вкус» и часто переводится как пикантный. Самые недавние исследования показали, что может существовать шестой вкус к жирам или липидам.
Вкус — особое чувство, связанное с языком. Поверхность языка, как и остальная полость рта, выстлана многослойным плоским эпителием. Приподнятые бугорки, называемые сосочками (единственное число = сосочек), содержат структуры для вкусовой трансдукции. Существует четыре типа сосочков в зависимости от их внешнего вида ([ссылка]): желобчатые, листовидные, нитевидные и грибовидные. В структуре сосочков находятся вкусовые рецепторы, которые содержат специализированные клетки вкусовых рецепторов для передачи вкусовых раздражителей.
Язык
Язык покрыт небольшими бугорками, называемыми сосочками, которые содержат вкусовые рецепторы, чувствительные к химическим веществам в пище или питье. Различные типы сосочков находятся в разных областях языка. Вкусовые сосочки содержат специализированные клетки вкусовых рецепторов, которые реагируют на химические раздражители, растворенные в слюне. Эти рецепторные клетки активируют сенсорные нейроны, входящие в состав лицевого и языкоглоточного нервов. LM × 1600. (Микрофотография предоставлена Регентами Медицинской школы Мичиганского университета © 2012 г.)
Соленый вкус — это просто восприятие ионов натрия (Na + ) в слюне.
Кислый вкус – это восприятие концентрации H + . Так же, как ионы натрия в соленых ароматизаторах, эти ионы водорода проникают в клетку и вызывают деполяризацию. Кислые вкусы — это, по сути, восприятие кислот в нашей пище. Повышение концентрации ионов водорода в слюне (снижение pH слюны) вызывает постепенное усиление градиентных потенциалов во вкусовых клетках. Например, апельсиновый сок, содержащий лимонную кислоту, будет иметь кислый вкус, потому что его значение pH приблизительно равно 3. Конечно, его часто подслащивают, чтобы замаскировать кислый вкус.
Первые два вкуса (соленый и кислый) вызываются катионами Na + и H + . Другие вкусы возникают в результате связывания пищевых молекул с рецептором, связанным с G-белком. Система передачи сигнала G-белка в конечном итоге приводит к деполяризации вкусовой клетки. Сладкий вкус – это чувствительность вкусовых клеток к присутствию глюкозы, растворенной в слюне. Другие моносахариды, такие как фруктоза, или искусственные подсластители, такие как аспартам (NutraSweet™), сахарин или сукралоза (Splenda™), также активируют рецепторы сладкого. Сродство к каждой из этих молекул варьируется, и некоторые из них будут слаще глюкозы, потому что они по-разному связываются с рецептором, связанным с G-белком.
Горький вкус похож на сладкий тем, что молекулы пищи связываются с рецепторами, связанными с G-белком. Однако существует множество различных способов, которыми это может произойти, поскольку существует большое разнообразие молекул с горьким вкусом. Некоторые горькие молекулы деполяризуют вкусовые клетки, тогда как другие гиперполяризуют вкусовые клетки.
Одной из основных групп молекул с горьким вкусом являются алкалоиды. Алкалоиды представляют собой азотсодержащие молекулы, которые обычно встречаются в горьких на вкус растительных продуктах, таких как кофе, хмель (в пиве), дубильные вещества (в вине), чай и аспирин. Благодаря содержанию токсичных алкалоидов растение менее восприимчиво к микробным инфекциям и менее привлекательно для травоядных.
Таким образом, функция горького вкуса может быть в первую очередь связана со стимуляцией рвотного рефлекса, чтобы избежать проглатывания ядов. Из-за этого многие горькие продукты, которые обычно употребляются в пищу, часто комбинируют со сладкими компонентами, чтобы сделать их более вкусными (например, сливки и сахар в кофе). Самая высокая концентрация горьких рецепторов, по-видимому, находится в задней части языка, где рвотный рефлекс все еще может выплевывать ядовитую пищу.
Вкус, известный как умами, часто называют пикантным вкусом. Подобно сладкому и горькому, он основан на активации рецепторов, связанных с G-белком, определенной молекулой. Молекула, которая активирует этот рецептор, представляет собой аминокислоту L-глутамат. Поэтому вкус умами часто ощущается при употреблении продуктов, богатых белком. Неудивительно, что блюда, содержащие мясо, часто называют пикантными.
Когда вкусовые клетки активируются вкусовыми молекулами, они высвобождают нейротрансмиттеры на дендриты сенсорных нейронов. Эти нейроны являются частью лицевого и языкоглоточного черепных нервов, а также компонентом блуждающего нерва, отвечающим за рвотный рефлекс. Лицевой нерв соединяется со вкусовыми рецепторами в передней трети языка. Языкоглоточный нерв соединяется со вкусовыми сосочками в задних двух третях языка. Блуждающий нерв соединяется со вкусовыми рецепторами в крайней задней части языка, граничащими с глоткой, которые более чувствительны к вредным раздражителям, таким как горечь.
Посмотрите это видео, чтобы узнать о докторе Даниэлле Рид из Центра химических ощущений Монелла в Филадельфии, штат Пенсильвания, которая заинтересовалась наукой в раннем возрасте из-за своего сенсорного опыта. Она признала, что ее чувство вкуса было уникальным по сравнению с другими людьми, которых она знала. Теперь она изучает генетические различия между людьми и их чувствительность к вкусовым раздражителям. На видео есть краткое изображение человека, высунувшего язык, покрытый цветной краской. Именно так доктор Рид может визуализировать и подсчитывать сосочки на поверхности языка. Люди делятся на две группы, известные как «дегустаторы» и «не дегустаторы», в зависимости от плотности сосочков на языке, что также указывает на количество вкусовых рецепторов. Недегустаторы могут ощущать вкус пищи, но они не так чувствительны к определенным вкусам, например к горечи. Доктор Рид обнаружила, что она не умеет дегустировать, что объясняет, почему она воспринимала горечь не так, как другие люди, которых она знала.
Обоняние (обоняние)
Как и вкус, обоняние также реагирует на химические раздражители. Нейроны обонятельных рецепторов расположены в небольшой области в верхней носовой полости ([ссылка]). Эта область называется обонятельным эпителием и содержит биполярные чувствительные нейроны. Каждый обонятельный сенсорный нейрон имеет дендриты, отходящие от апикальной поверхности эпителия в слизь, выстилающую полость. Когда переносимые по воздуху молекулы вдыхаются через нос, они проходят над областью обонятельного эпителия и растворяются в слизи. Эти пахучие молекулы связываются с белками, которые растворяют их в слизи и помогают транспортировать их к обонятельным дендритам. Комплекс пахучий белок связывается с белком-рецептором внутри клеточной мембраны обонятельного дендрита. Эти рецепторы связаны с G-белком и будут создавать градуированный мембранный потенциал в обонятельных нейронах.
Аксон обонятельного нейрона идет от базальной поверхности эпителия через обонятельное отверстие в решетчатой пластинке решетчатой кости в головной мозг. Группа аксонов, называемая обонятельным трактом, соединяется с обонятельной луковицей на вентральной поверхности лобной доли. Оттуда аксоны расходятся, чтобы отправиться в несколько областей мозга. Некоторые из них перемещаются в головной мозг, особенно в первичную обонятельную кору, расположенную в нижней и медиальной областях височной доли. Другие проецируются на структуры лимбической системы и гипоталамуса, где запахи ассоциируются с долговременной памятью и эмоциональными реакциями. Именно так определенные запахи вызывают эмоциональные воспоминания, например, запах еды, связанный с местом рождения. Запах — это единственная сенсорная модальность, которая не образует синапсов в таламусе до того, как соединяется с корой головного мозга. Эта тесная связь между обонятельной системой и корой головного мозга является одной из причин, по которой запах может быть мощным триггером воспоминаний и эмоций.
Эпителий носа, включая обонятельные клетки, может быть поврежден переносимыми по воздуху токсичными химическими веществами. Следовательно, обонятельные нейроны регулярно заменяются внутри носового эпителия, после чего аксоны новых нейронов должны найти соответствующие соединения в обонятельной луковице. Эти новые аксоны растут вдоль аксонов, которые уже находятся в черепном нерве.
Обонятельная система
(a) Обонятельная система начинается в периферических структурах полости носа. (б) Нейроны обонятельных рецепторов находятся внутри обонятельного эпителия. (c) Аксоны нейронов обонятельных рецепторов проходят через решетчатую пластинку решетчатой кости и синапсы с нейронами обонятельной луковицы (источник ткани: обезьяна). LM × 812. (Микрофотография предоставлена Регентами Медицинской школы Мичиганского университета © 2012)
Заболевания…
Обонятельная система: аносмия
Травма лица тупым предметом, например, во многих автомобильных авариях, может привести к потере обонятельного нерва и впоследствии к потере чувства запах.Потеря обоняния может сделать вкус пищи безвкусным. Человеку с нарушением обоняния может потребоваться дополнительное количество специй и приправ для вкуса пищи. Аносмия также может быть связана с некоторыми проявлениями легкой депрессии, потому что потеря удовольствия от еды может привести к общему чувству отчаяния.
Способность обонятельных нейронов к самовосстановлению снижается с возрастом, что приводит к возрастной аносмии. Это объясняет, почему некоторые пожилые люди солят пищу больше, чем молодые. Однако это повышенное потребление натрия может увеличить объем крови и кровяное давление, увеличивая риск сердечно-сосудистых заболеваний у пожилых людей.
Прослушивание (Слух)
Слух или прослушивание — это преобразование звуковых волн в нервный сигнал, которое стало возможным благодаря структурам уха ([ссылка]). Большая мясистая структура на боковой стороне головы известна как ушная раковина. В некоторых источниках эта структура также упоминается как ушная раковина, хотя этот термин больше подходит для подвижной структуры, такой как внешнее ухо кошки. С-образные изгибы ушной раковины направляют звуковые волны в сторону слухового прохода. Канал входит в череп через наружный слуховой проход височной кости. В конце слухового прохода находится барабанная перепонка, или барабанная перепонка, которая вибрирует после удара звуковой волны.
Структуры уха
Наружное ухо содержит ушную раковину, слуховой проход и барабанную перепонку. Среднее ухо содержит слуховые косточки и соединяется с глоткой евстахиевой трубой. Внутреннее ухо содержит улитку и преддверие, которые отвечают за слух и равновесие соответственно.
Внутреннее ухо часто называют костным лабиринтом, так как оно состоит из ряда каналов, встроенных в височную кость. Он состоит из двух отдельных областей, улитки и преддверия, которые отвечают за слух и равновесие соответственно. Нервные сигналы от этих двух областей передаются в ствол мозга через отдельные пучки волокон. Однако эти два отдельных пучка идут вместе от внутреннего уха к стволу мозга в виде преддверно-улиткового нерва. Звук преобразуется в нервные сигналы в кохлеарной области внутреннего уха, которая содержит сенсорные нейроны спиральных ганглиев. Эти ганглии расположены внутри спиралевидной улитки внутреннего уха. Улитка прикрепляется к стремени через овальное окно.
Овальное окно расположено в начале заполненной жидкостью трубки внутри улитки, называемой вестибулярной лестницей. Лестница преддверия отходит от овального окна, проходя над улитковым протоком, который является центральной полостью улитки, содержащей нейроны, передающие звук. На самом верхнем конце улитки вестибулярная лестница изгибается над верхушкой улиткового канала.
Передача звуковых волн в улитку
Звуковая волна вызывает вибрацию барабанной перепонки. Эта вибрация усиливается при движении по молоточку, наковальне и стремени. Усиленная вибрация улавливается овальным окном, вызывающим волны давления в жидкости вестибулярной лестницы и барабанной лестницы. Сложность волн давления определяется изменениями амплитуды и частоты звуковых волн, поступающих в ухо.
Поперечное сечение улитки показывает, что вестибулярная лестница и барабанная лестница проходят вдоль обеих сторон улиткового канала ([ссылка]).
Поперечный разрез улитки
Выделены три основных пространства внутри улитки. Барабанная лестница и вестибулярная лестница лежат по обе стороны от улиткового протока. Кортиев орган, содержащий волосковые клетки механорецепторов, прилегает к барабанной лестнице, где он расположен на базилярной мембране.
Кортиевы органы содержат волосковые клетки, названные в честь волосовидных стереоцилий, отходящих от апикальной поверхности клетки ([ссылка]).
Волосковая клетка
Волосковая клетка представляет собой механорецептор с набором стереоцилий, выходящих из его апикальной поверхности. Стереоцилии связаны вместе белками, которые открывают ионные каналы, когда массив изгибается в сторону самого высокого члена их массива, и закрываются, когда массив изгибается в сторону самого короткого члена их массива.
Улитка и Кортиев орган
LM × 412. (Микрофотография предоставлена Регентами Медицинской школы Мичиганского университета © 2012)
Просмотрите WebScope Мичиганского университета, чтобы более подробно изучить образец ткани. Базилярная мембрана представляет собой тонкую мембрану, простирающуюся от центрального ядра улитки до края. Что прикреплено к этой мембране, чтобы их можно было активировать движением жидкости внутри улитки?
Как указано выше, данная область базилярной мембраны будет двигаться только в том случае, если входящий звук имеет определенную частоту.
Частотное кодирование в улитке
Стоячая звуковая волна, генерируемая в улитке движением овального окна, отклоняет базилярную мембрану в зависимости от частоты звука. Следовательно, волосковые клетки в основании улитки активируются только высокими частотами, тогда как клетки в верхушке улитки активируются только низкими частотами.
Посмотрите это видео, чтобы узнать больше о том, как структуры уха преобразуют звуковые волны в нервный сигнал, перемещая «волоски» или стереоцилии улиткового канала. Определенные места по длине воздуховода кодируют определенные частоты или высоты тона. Мозг интерпретирует значение звуков, которые мы слышим, как музыку, речь, шум и т. д. Какие структуры уха отвечают за усиление и передачу звука от наружного уха к внутреннему уху?
Посмотрите эту анимацию, чтобы узнать больше о внутреннем ухе и увидеть, как разворачивается улитка, основание которой находится сзади, а верхушка — впереди. Определенные длины волн звука вызывают вибрацию определенных областей базилярной мембраны, подобно тому, как клавиши пианино производят звук на разных частотах.
Равновесие (Баланс)
Наряду со слухом внутреннее ухо отвечает за кодирование информации о равновесии, чувстве равновесия. Аналогичный механорецептор — волосковая клетка со стереоцилиями — определяет положение головы, движение головы и движение тела. Эти клетки расположены в преддверии внутреннего уха. Положение головы ощущается маточкой и мешочком, тогда как движение головы ощущается полукружными каналами. Нервные сигналы, генерируемые в вестибулярном ганглии, передаются через преддверно-улитковый нерв к стволу головного мозга и мозжечку.
Маточка и мешочек в основном состоят из ткани макулы (множественное число = макулы). Макула состоит из волосковых клеток, окруженных опорными клетками. Стереоцилии волосковых клеток превращаются в вязкий гель, называемый отолитовой мембраной ([ссылка]). Поверх отолитовой мембраны находится слой кристаллов карбоната кальция, называемый отолитами.
Кодирование линейного ускорения с помощью пятен
Пятна специализируются на восприятии линейного ускорения, например, когда гравитация действует на наклоняющуюся голову или голова начинает двигаться по прямой линии. Разница в инерции между стереоцилиями волосковых клеток и отолитовой мембраной, в которую они встроены, приводит к сдвигающей силе, которая заставляет стереоцилии изгибаться в направлении этого линейного ускорения.
Полукружные каналы представляют собой три кольцеобразных продолжения преддверия.
Вращательное кодирование полукружными каналами
Вращательное движение головы кодируется волосковыми клетками в основании полукружных каналов. Когда один из каналов движется по дуге вместе с головкой, внутренняя жидкость движется в противоположном направлении, вызывая искривление купулы и стереоцилий. Движение двух каналов в плоскости дает информацию о направлении, в котором движется голова, а активация всех шести каналов может дать очень точное указание на движение головы в трех измерениях.
Соматоощущение (осязание)
Соматоощущение считается общим чувством, в отличие от особых чувств, обсуждаемых в этом разделе. Соматосенсоры — это группа сенсорных модальностей, связанных с осязанием, проприоцепцией и интероцепцией. Эти модальности включают давление, вибрацию, легкое прикосновение, щекотку, зуд, температуру, боль, проприоцепцию и кинестезию. Это означает, что его рецепторы не связаны со специализированным органом, а вместо этого распределены по всему телу в различных органах.
Двумя типами соматосенсорных сигналов, которые передаются свободными нервными окончаниями, являются боль и температура. Эти два модальности используют терморецепторы и ноцицепторы для преобразования температурных и болевых раздражителей соответственно. Температурные рецепторы стимулируются, когда локальные температуры отличаются от температуры тела. Некоторые терморецепторы чувствительны только к холоду, а другие — только к теплу. Ноцицепция – это ощущение потенциально повреждающих раздражителей. Механические, химические или термические раздражители, превышающие установленный порог, вызывают болезненные ощущения. Напряженные или поврежденные ткани выделяют химические вещества, которые активируют рецепторные белки в ноцицепторах. Например, ощущение тепла, связанное с острой пищей, связано с капсаицином, активной молекулой острого перца.
Если провести пальцем по текстурированной поверхности, кожа пальца начнет вибрировать. Такие низкочастотные колебания воспринимаются механорецепторами, называемыми клетками Меркеля, также известными как кожные механорецепторы I типа. Клетки Меркеля располагаются в базальном слое эпидермиса. Глубокое давление и вибрация передаются пластинчатыми (Пачиниевыми) тельцами, которые представляют собой рецепторы с инкапсулированными окончаниями, расположенными глубоко в дерме или подкожной ткани.
Другие соматосенсорные рецепторы находятся в суставах и мышцах. Рецепторы растяжения контролируют растяжение сухожилий, мышц и компонентов суставов. Например, вы когда-нибудь растягивали мышцы до или после тренировки и замечали, что вы можете растягиваться только до тех пор, пока ваши мышцы не сокращаются и возвращаются в менее растянутое состояние? Этот спазм является рефлексом, который инициируется рецепторами растяжения, чтобы избежать разрыва мышц. Такие рецепторы растяжения могут также предотвратить чрезмерное сокращение мышцы.
*Нет соответствующего одноименного имени. Механорецепторы соматоощущения Имя Историческое (одноименное) название Местоположение(а) Стимулы Свободные нервные окончания * Дерма, роговица, язык, суставные капсулы, внутренние органы Боль, температура, механическая деформация Механорецепторы Диски Меркеля Эпидермо-дермальное соединение, слизистые оболочки Низкочастотная вибрация (5–15 Гц) Луковичное тельце Тельца Руффини Дерма, суставные капсулы Растяжка Тактильное тельце Тельца Мейснера Папиллярная дерма, особенно на кончиках пальцев и губах Легкое прикосновение, вибрация ниже 50 Гц Пластинчатое тельце тельца Пачини Глубокая дерма, подкожная клетчатка Глубокое давление, высокочастотная вибрация (около 250 Гц) Сплетение волосяного фолликула * Оборачивают волосяные фолликулы в дерме Движение волос Мышечное веретено * В соответствии со скелетными мышечными волокнами Сокращение и растяжение мышц Орган растяжения сухожилий Сухожильный орган Гольджи В соответствии с сухожилиями Растяжение сухожилий Зрение
Зрение – это особое чувство зрения, основанное на преобразовании световых раздражителей, поступающих через глаза.
Глаз в орбите
Глаз расположен внутри орбиты и окружен мягкими тканями, защищающими и поддерживающими его функцию. Орбита окружена черепными костями черепа.
Движение глаза внутри орбиты осуществляется за счет сокращения шести экстраокулярных мышц, которые берут начало от костей орбиты и прикрепляются к поверхности глазного яблока ([ссылка]).
Экстраокулярные мышцы иннервируются тремя черепными нервами. Латеральная прямая мышца, обеспечивающая отведение глаза, иннервируется отводящим нервом. Верхняя косая иннервируется блоковым нервом. Все остальные мышцы иннервируются глазодвигательным нервом, как и мышца, поднимающая верхнее веко. Двигательные ядра этих черепных нервов соединяются со стволом мозга, который координирует движения глаз.
Экстраокулярные мышцы
Экстраокулярные мышцы перемещают глаз по орбите.
Сам глаз представляет собой полую сферу, состоящую из трех слоев ткани. Самый внешний слой представляет собой фиброзную оболочку, которая включает белую склеру и прозрачную роговицу.
Глаз также делится на две полости: переднюю полость и заднюю полость. Передняя полость представляет собой пространство между роговицей и хрусталиком, включая радужную оболочку и цилиарное тело. Он заполнен водянистой жидкостью, называемой водянистой влагой. Задняя полость — это пространство позади хрусталика, простирающееся до задней стороны внутренней части глазного яблока, где расположена сетчатка. Задняя полость заполнена более вязкой жидкостью, называемой стекловидным телом.
Сетчатка состоит из нескольких слоев и содержит специализированные клетки для первичной обработки зрительных стимулов. Фоторецепторы (палочки и колбочки) меняют свой мембранный потенциал при стимуляции световой энергией. Изменение мембранного потенциала изменяет количество нейротрансмиттера, которое фоторецепторные клетки выделяют на биполярные клетки во внешнем синаптическом слое.
Строение глаза
Сферу глаза можно разделить на переднюю и заднюю камеры. Стенка глаза состоит из трех слоев: фиброзной оболочки, сосудистой оболочки и нервной оболочки. В нервной оболочке находится сетчатка с тремя слоями клеток и двумя синапическими слоями между ними. В центре сетчатки есть небольшое углубление, известное как ямка.
Обратите внимание, что фоторецепторы в сетчатке (палочки и колбочки) расположены позади аксонов, ГКС, биполярных клеток и кровеносных сосудов сетчатки.
Свет, падающий на сетчатку, вызывает химические изменения молекул пигмента в фоторецепторах, что в конечном итоге приводит к изменению активности ГКС. Фоторецепторные клетки состоят из двух частей: внутреннего сегмента и внешнего сегмента ([ссылка]). Внутренний сегмент содержит ядро и другие общие органеллы клетки, тогда как внешний сегмент представляет собой специализированную область, в которой происходит фоторецепция. Существует два типа фоторецепторов — палочки и колбочки, различающиеся по форме внешнего сегмента.
Фоторецептор
(a) Все фоторецепторы имеют внутренние сегменты, содержащие ядро и другие важные органеллы, и внешние сегменты с мембранными массивами, содержащими светочувствительные молекулы опсина. Наружные сегменты стержней представляют собой длинные столбчатые формы со стопками мембранных дисков, содержащих пигмент родопсин. Наружные сегменты конусов короткие, конической формы со складками мембраны вместо дисков в стержнях.
На молекулярном уровне зрительные раздражители вызывают изменения в молекуле фотопигмента, что приводит к изменению мембранного потенциала фоторецепторной клетки. Одна единица света называется фотоном, который описывается в физике как пакет энергии со свойствами как частицы, так и волны. Энергия фотона представлена его длиной волны, причем каждая длина волны видимого света соответствует определенному цвету. Видимый свет — это электромагнитное излучение с длиной волны от 380 до 720 нм. Длины волн электромагнитного излучения более 720 нм попадают в инфракрасный диапазон, тогда как длины волн короче 380 нм попадают в ультрафиолетовый диапазон. Свет с длиной волны 380 нм — синий, тогда как свет с длиной волны 720 нм — темно-красный. Все остальные цвета находятся между красным и синим в различных точках по шкале длин волн.
Пигменты опсина на самом деле являются трансмембранными белками, которые содержат кофактор, известный как ретиналь. Ретиналь представляет собой углеводородную молекулу, связанную с витамином А. Когда фотон попадает на сетчатку, длинная углеводородная цепь молекулы биохимически изменяется. В частности, фотоны заставляют некоторые атомы углерода с двойными связями в цепи переключаться с цис на транс конформацию. Этот процесс называется фотоизомеризацией. Прежде чем взаимодействовать с фотоном, гибкие углероды сетчатки с двойными связями находятся в цис- конформация. Эта молекула обозначается как 11- цис -ретиналь. Фотон, взаимодействующий с молекулой, заставляет гибкие атомы углерода с двойными связями изменяться в конформацию транс –, образуя полностью транс -ретиналь, который имеет прямую углеводородную цепь ([ссылка]).
Изменение формы сетчатки в фоторецепторах инициирует зрительную трансдукцию в сетчатке. Активация белков сетчатки и опсина приводит к активации G-белка. Белок G изменяет мембранный потенциал фоторецепторной клетки, которая затем высвобождает меньше нейротрансмиттера во внешний синаптический слой сетчатки. Пока молекула сетчатки не изменится обратно на 11- цис — форма сетчатки, опсин не может реагировать на световую энергию, что называется обесцвечиванием. Когда большая группа фотопигментов обесцвечивается, сетчатка будет посылать информацию, как если бы воспринималась противоположная визуальная информация. После яркой вспышки света остаточные изображения обычно видны на негативе. Фотоизомеризация обращается вспять серией ферментативных изменений, так что сетчатка реагирует на большее количество световой энергии.
Изомеры сетчатки
Молекула сетчатки имеет два изомера: (а) один до взаимодействия с фотоном и (б) один, который изменяется в результате фотоизомеризации.
Опсины чувствительны к свету ограниченной длины волны. Родопсин, фотопигмент палочек, наиболее чувствителен к свету с длиной волны 498 нм. Три цветных опсина имеют пиковую чувствительность 564 нм, 534 нм и 420 нм, что примерно соответствует основным цветам: красному, зеленому и синему ([ссылка]). Поглощение родопсина у палочек гораздо более чувствительно, чем у опсинов колбочек; в частности, палочки чувствительны к зрению в условиях низкой освещенности, а колбочки чувствительны к более ярким условиям. При обычном солнечном свете родопсин будет постоянно обесцвечиваться, пока колбочки активны. В затемненной комнате света недостаточно для активации опсинов колбочек, и зрение полностью зависит от палочек. Палочки настолько чувствительны к свету, что один фотон может привести к возникновению потенциала действия от соответствующего RGC палочки.
Три типа опсинов колбочек, чувствительных к разным длинам волн света, обеспечивают нам цветовое зрение. Сравнивая активность трех разных колбочек, мозг может извлекать цветовую информацию из визуальных стимулов.
Сравнение цветовой чувствительности фотопигментов
Сравнение спектров пиковой чувствительности и поглощения четырех фотопигментов показывает, что они наиболее чувствительны к определенным длинам волн.
Посмотрите это видео, чтобы узнать больше о поперечном срезе мозга, на котором изображен зрительный путь от глаза к затылочной коре.
Как только любая сенсорная клетка преобразует стимул в нервный импульс, этот импульс должен пройти по аксонам, чтобы достичь ЦНС. Во многих особых чувствах аксоны, отходящие от сенсорных рецепторов, имеют топографическое расположение, что означает, что расположение сенсорного рецептора связано с расположением аксона в нерве.
Спинномозговые нервы
Как правило, спинномозговые нервы содержат афферентные аксоны от сенсорных рецепторов на периферии, например, от кожи, смешанные с эфферентными аксонами, направляющимися к мышцам или другим эффекторным органам. Когда спинномозговой нерв приближается к спинному мозгу, он разделяется на дорсальный и вентральный корешки. Дорсальные корешки содержат только аксоны сенсорных нейронов, тогда как вентральные корешки содержат только аксоны двигательных нейронов. Некоторые из ветвей образуют синапсы с локальными нейронами ганглия задних корешков, задних (дорсальных) рогов или даже передних (вентральных) рогов на уровне спинного мозга, куда они входят. Другие ветви будут перемещаться на небольшое расстояние вверх или вниз по позвоночнику, чтобы взаимодействовать с нейронами на других уровнях спинного мозга.
Черепные нервы
Черепные нервы передают специфическую сенсорную информацию от головы и шеи непосредственно в мозг. Для ощущений ниже шеи правая сторона тела связана с левой стороной мозга, а левая сторона тела — с правой стороной мозга. В то время как спинная информация является контралатеральной, системы черепных нервов в основном ипсилатеральны, а это означает, что черепной нерв на правой стороне головы связан с правой стороной мозга.
К органам чувств относятся обоняние (обоняние), вкус (вкус), соматосенсоризация (ощущения, связанные с кожей и телом), слух (слух), равновесие (равновесие) и зрение. За исключением соматоощущения, этот список представляет особые чувства или те системы тела, которые связаны с определенными органами, такими как язык или глаз. Соматоощущение относится к общим органам чувств, представляющим собой те чувствительные структуры, которые распределены по всему телу и в стенках различных органов. Все специальные чувства в первую очередь являются частью соматической нервной системы, поскольку они сознательно воспринимаются через мозговые процессы, хотя некоторые особые чувства способствуют вегетативной функции.
Клетки, преобразующие сенсорные стимулы в электрохимические сигналы нервной системы, классифицируются на основе структурных или функциональных аспектов клеток. Структурные классификации основаны либо на анатомии клетки, взаимодействующей со стимулом (свободные нервные окончания, инкапсулированные окончания или специализированная рецепторная клетка), либо на том, где клетка расположена относительно раздражителя (интерорецептор, экстерорецептор, проприоцептор). В-третьих, функциональная классификация основана на том, как клетка преобразует стимул в нервный сигнал. Хеморецепторы реагируют на химические раздражители и являются основой для обоняния и вкуса.
Нервы, передающие сенсорную информацию от периферии к ЦНС, представляют собой либо спинномозговые нервы, связанные со спинным мозгом, либо черепные нервы, связанные с головным мозгом. Спинномозговые нервы имеют смешанные популяции волокон; некоторые из них являются двигательными волокнами, а некоторые — сенсорными. Чувствительные волокна соединяются со спинным мозгом через задний корешок, который прикрепляется к заднему корешковому ганглию. Сенсорная информация от тела, которая передается через спинномозговые нервы, будет проецироваться на противоположную сторону мозга для обработки корой головного мозга.
Посмотрите это видео, чтобы узнать о докторе Даниэлле Рид из Центра химических ощущений Монелла в Филадельфии, штат Пенсильвания, которая в раннем возрасте заинтересовалась наукой из-за своего сенсорного опыта. Она признала, что ее чувство вкуса было уникальным по сравнению с другими людьми, которых она знала. Теперь она изучает генетические различия между людьми и их чувствительность к вкусовым раздражителям. На видео есть краткое изображение человека, высунувшего язык, покрытый цветной краской. Именно так доктор Рид может визуализировать и подсчитывать сосочки на поверхности языка. Люди делятся на две большие группы, известные как «дегустаторы» и «не дегустаторы», в зависимости от плотности сосочков на языке, что также указывает на количество вкусовых рецепторов.
Ответы могут быть разными, но типичный ответ может быть таким: я могу есть почти все (кроме грибов!), поэтому я не думаю, что я настолько чувствителен к вкусам. Вся моя семья любит есть разнообразную пищу, поэтому кажется, что у всех нас одинаковый уровень чувствительности.
[ссылка] Базилярная мембрана представляет собой тонкую мембрану, простирающуюся от центрального ядра улитки до края. Что прикреплено к этой мембране, чтобы их можно было активировать движением жидкости внутри улитки?
[ссылка] Волосковые клетки располагаются в кортиевом органе, расположенном на базилярной мембране. Стереоцилии этих клеток обычно прикреплены к текториальной мембране (хотя на микрофотографии они отделены из-за обработки ткани).
Посмотрите это видео, чтобы узнать больше о том, как структуры уха преобразуют звуковые волны в нервный сигнал, двигая «волоски» или стереоцилии улиткового канала. Определенные места по длине воздуховода кодируют определенные частоты или высоты тона. Мозг интерпретирует значение звуков, которые мы слышим, как музыку, речь, шум и т. д. Какие структуры уха отвечают за усиление и передачу звука от наружного уха к внутреннему уху?
Мелкие косточки среднего уха, косточки, усиливают и передают звук между барабанной перепонкой наружного уха и овальным окном внутреннего уха.
Посмотрите эту анимацию, чтобы узнать больше о внутреннем ухе и увидеть, как разворачивается улитка, основание которой находится сзади, а верхушка — впереди. Определенные длины волн звука вызывают вибрацию определенных областей базилярной мембраны, подобно тому, как клавиши пианино производят звук на разных частотах. Судя по анимации, какие частоты — от высоких до низких — вызывают активность волосковых клеток в улитковом канале?
Высокие частоты активируют волосковые клетки в направлении основания улитки, а низкие частоты активируют волосковые клетки в направлении вершины улитки.
Посмотрите это видео, чтобы узнать больше о поперечном срезе мозга, изображающем зрительный путь от глаза к затылочной коре. Первая половина пути представляет собой проекцию ГКС через зрительный нерв к латеральному коленчатому телу в таламусе с обеих сторон. Это первое волокно в проводящем пути синапсирует с таламической клеткой, которая затем проецируется в зрительную кору затылочной доли, где происходит «видение» или зрительное восприятие. Это видео дает сокращенный обзор зрительной системы, концентрируясь на пути от глаз к затылочной доле. В видео делается заявление (на 0:45), что «специализированные клетки сетчатки, называемые ганглиозными клетками, преобразуют световые лучи в электрические сигналы». Какой аспект обработки сетчатки упрощается этим утверждением? Поясните свой ответ.
Фоторецепторы преобразуют световую энергию или фотоны в электрохимический сигнал. Сетчатка содержит биполярные клетки и ГКС, которые в конечном итоге преобразуют их в потенциалы действия, которые передаются от сетчатки к ЦНС.
Какой тип рецепторных клеток отвечает за передачу болевых раздражителей?
- механорецептор
- ноцицептор
- осморецептор
- фоторецептор
B
Какой из этих черепных нервов является частью вкусовой системы?
- обонятельный
- блок
- тройничный нерв
- лица
D
Какая субмодальность вкуса чувствительна к рН слюны?
- умами
- кислый
- горький
- сладкий
B
Аксоны какого нейрона сетчатки образуют зрительный нерв?
- амакриновые клетки
- фоторецепторы
- биполярные клетки
- ганглиозных клеток сетчатки
D
Какой тип рецепторных клеток участвует в ощущении звука и равновесия?
- фоторецептор
- хеморецептор
- механорецептор
- ноцицептор
С
Подсластитель, известный как стевия, может заменить глюкозу в пище.
Молекула стевии похожа на глюкозу, поэтому она связывается с рецептором глюкозы в рецепторах, чувствительных к сладкому. Однако он не является субстратом для АТФ-генерирующего метаболизма внутри клеток.
Почему слепое пятно диска зрительного нерва в любом глазу не приводит к слепому пятну в поле зрения?
Поле зрения каждого глаза проецируется на сетчатку по мере того, как хрусталик фокусирует свет. Зрительная информация из правого поля зрения попадает на левую сторону сетчатки и наоборот. Диск зрительного нерва в правом глазу находится на медиальной стороне центральной ямки, которая будет левой стороной сетчатки. Однако диск зрительного нерва в левом глазу будет находиться на правой стороне этой ямки, поэтому правое поле зрения падает на сторону сетчатки в левом поле, где нет слепого пятна.
Глоссарий
- алкалоид
- вещество, обычно растительного происхождения, химически щелочное по отношению к pH и стимулирующее горькие рецепторы
- амакриновая клетка
- тип клеток в сетчатке, который соединяется с биполярными клетками вблизи внешнего синаптического слоя и обеспечивает основу для ранней обработки изображений в сетчатке
- ампула
- в ухе, структура в основании полукружного канала, которая содержит волосковые клетки и купол для передачи вращательного движения головы
- аносмия
- потеря обоняния; обычно в результате физического повреждения первого черепного нерва
- водянистая влага
- водянистая жидкость, заполняющая переднюю камеру, содержащую роговицу, радужную оболочку, цилиарное тело и хрусталик глаза
- прослушивание
- слух
- ушная раковина
- мясистая внешняя структура уха
- базилярная мембрана
- в ухе дно улиткового канала, на котором располагается кортиев орган
- биполярный элемент
Тип клеток- в сетчатке, который соединяет фоторецепторы с RGCs
- капсаицин
- молекула, которая активирует ноцицепторы, взаимодействуя с чувствительным к температуре ионным каналом, и является основой для «горячих» ощущений в острой пище
- хеморецептор
- сенсорная рецепторная клетка, чувствительная к химическим раздражителям, таким как вкус, запах или боль
- сосудистая оболочка
- сильно сосудистая ткань в стенке глаза, которая снабжает кровью наружную часть сетчатки
- цилиарное тело
- структура гладкой мускулатуры на внутренней поверхности радужной оболочки, которая контролирует форму хрусталика посредством волокон цинновых связок
- улитка
- слуховая часть внутреннего уха, содержащая структуры для преобразования звуковых раздражителей
- улитковый канал
- пространство внутри слуховой части внутреннего уха, содержащее кортиев орган и прилежащее с обеих сторон к барабанной и вестибулярной лестницам
- колбочковый фоторецептор
- один из двух типов рецепторных клеток сетчатки, которые специализируются на цветовом зрении за счет использования трех фотопигментов, распределенных по трем отдельным популяциям клеток
- контралатеральный
- слово, означающее «на противоположной стороне», как в аксонах, которые пересекают среднюю линию в волокнистом тракте
- роговица
- фиброзное покрытие передней области глаза, прозрачное для прохождения света
- купула
- специализированная структура в основании полукружного канала, изгибающая стереоцилии волосковых клеток при вращении головы за счет относительного движения заключенной жидкости
- инкапсулированное окончание
- конфигурация сенсорного рецепторного нейрона с дендритами, окруженными специализированными структурами, помогающими в передаче определенного типа ощущений, такими как пластинчатые тельца в глубоких слоях дермы и подкожной клетчатке
- равновесие
- чувство равновесия, включающее ощущение положения и движения головы
- наружное ухо
- структур на боковой поверхности головы, включая ушную раковину и слуховой проход сзади до барабанной перепонки
- экстерорецептор
- сенсорный рецептор, предназначенный для интерпретации стимулов из внешней среды, таких как фоторецепторы в глазах или соматосенсорные рецепторы в коже
- экстраокулярная мышца
- одна из шести мышц, берущих начало от костей глазницы и прикрепляющихся к поверхности глаза и отвечающих за движение глаза
- волокнистая оболочка
- наружный слой глаза, состоящий в основном из соединительной ткани, известной как склера и роговица
- ямка
- точный центр сетчатки, в котором зрительные стимулы фокусируются с максимальной остротой, где сетчатка наиболее тонкая, в которой нет ничего, кроме фоторецепторов
- свободное нервное окончание
- конфигурация сенсорного рецепторного нейрона с дендритами в соединительной ткани органа, например в дерме кожи, которые чаще всего чувствительны к химическим, термическим и механическим раздражителям
- общий смысл
- любая сенсорная система, которая распределена по всему телу и включена в органы многих других систем, например стенки органов пищеварения или кожа
- вкус
- чувство вкуса
- клетки вкусовых рецепторов
- сенсорных клеток во вкусовой почке, которые передают химические стимулы вкуса
- волосковые клетки
- механорецепторных клеток, обнаруженных во внутреннем ухе, которые передают стимулы органам слуха и чувства равновесия
- наковальня
- (также наковальня) косточка среднего уха, соединяющая молоточек со стремечком
- нижняя косая
- экстраокулярная мышца, отвечающая за боковое вращение глаза
- нижняя прямая мышца
- экстраокулярная мышца, отвечающая за взгляд вниз
- внутреннее ухо
- структура в височной кости, содержащая сенсорный аппарат слуха и равновесия
- внутренний сегмент
- в глазу, участок фоторецептора, который содержит ядро и другие основные органеллы для нормальных клеточных функций
- внутренний синаптический слой
Слой- в сетчатке, где биполярные клетки соединяются с RGCs
- интероцептор
- сенсорный рецептор, предназначенный для интерпретации стимулов от внутренних органов, таких как рецепторы растяжения в стенке кровеносных сосудов
- ипсилатеральный
- значение слова на той же стороне, что и в аксонах, которые не пересекают среднюю линию в волокнистом тракте
- ирис
- окрашенная часть передней части глаза, окружающая зрачок
- кинестезия
- чувство движения тела, основанное на ощущениях в скелетных мышцах, сухожилиях, суставах и коже
- слезный канал
- проток в медиальном углу глазницы, отводящий слезы в носовую полость
- слезная железа
- железа латеральнее орбиты, которая производит слезы для омывания поверхности глаза
- латеральная прямая мышца
- экстраокулярная мышца, отвечающая за отведение глаза
- линза
- компонент глаза, фокусирующий свет на сетчатке
- поднимающая верхнее веко
- мышца, вызывающая подъем верхнего века, контролируемая волокнами глазодвигательного нерва
- макула
- расширение в основании полукружного канала, при котором происходит передача равновесных раздражителей внутри ампулы
- молоточек
- (также молоточковая) косточка, прикрепляющаяся непосредственно к барабанной перепонке
- механорецептор
- Рецепторная клетка, преобразующая механические раздражители в электрохимический сигнал
- медиальная прямая мышца
- экстраокулярная мышца, отвечающая за приведение глаза
- среднее ухо
- пространство внутри височной кости между слуховым проходом и костным лабиринтом, где слуховые косточки усиливают звуковые волны от барабанной перепонки к овальному окну
- нервная оболочка
- слой глаза, содержащий нервную ткань, а именно сетчатку
- ноцицептор
- рецепторная клетка, воспринимающая болевые раздражители
- молекулы одоранта
- летучие химические вещества, которые связываются с рецепторными белками в обонятельных нейронах, чтобы стимулировать обоняние
- обоняние
- обоняние
- обонятельная луковица
- центральная мишень первого черепного нерва; расположен на вентральной поверхности лобной доли головного мозга
- обонятельный эпителий
- область носового эпителия, где расположены обонятельные нейроны
- обонятельный сенсорный нейрон
- рецепторная клетка обонятельной системы, чувствительная к химическим раздражителям обоняния, аксоны которой составляют первый черепной нерв
- опсин
Белок- , содержащий светочувствительный кофактор ретиналь для фототрансдукции
- диск зрительного нерва
- точка на сетчатке, в которой аксоны ГКС выходят из глаза и проходят кровеносные сосуды внутренней части сетчатки
- зрительный нерв
- второй черепной нерв, отвечающий за зрительное ощущение
- орган Корти
- структура в улитке, в которой волосковые клетки преобразуют движения звуковых волн в электрохимические сигналы
- осморецептор
- рецепторная клетка, воспринимающая разницу в концентрации телесных жидкостей на основе осмотического давления
- косточки
- три маленькие косточки в среднем ухе
- отолит
- слой кристаллов карбоната кальция, расположенный поверх отолитовой мембраны
- отолитовая мембрана
- студенистое вещество в маточке и мешочке внутреннего уха, содержащее кристаллы карбоната кальция и в которое встроены стереоцилии волосковых клеток
- внешний сегмент
- в глазу, участок фоторецептора, содержащий молекулы опсина, передающие световые стимулы
- внешний синаптический слой
- слой сетчатки, в котором фоторецепторы соединяются с биполярными клетками
- овальное окно
- мембрана у основания улитки в месте прикрепления стремени, отмечающая начало вестибулярной лестницы
- конъюнктива век
- мембрана, прикрепляемая к внутренней поверхности век, покрывающая переднюю поверхность роговицы
- сосочек
- для вкуса, бугоркообразного выступа на поверхности языка, содержащего вкусовые сосочки
- фотоизомеризация
- химическое изменение в молекуле ретиналя, которое изменяет связь таким образом, что она переключается с 11- цис-изомера -ретиналя на полностью- транс-изомер -ретиналя
- фотон
- индивидуальный «пакет» света
- фоторецептор
Рецепторная клетка- , специализирующаяся на ответе на световые раздражители
- проприоцепция
- чувство положения и движения тела
- проприоцептор
- рецепторная клетка, воспринимающая изменения положения и кинестетических аспектов тела
- ученик
- открытое отверстие в центре радужной оболочки, через которое свет проходит в глаз
- рецепторная клетка
- клетка, преобразующая стимулы окружающей среды в нервные сигналы
- сетчатка
- нервная ткань глаза, в которой происходит фототрансдукция
- сетчатка
- кофактор в молекуле опсина, претерпевающий биохимические изменения при попадании фотона (произносится с ударением на последнем слоге)
- ганглиозные клетки сетчатки (RGC)
- нейрон сетчатки, который проецируется вдоль II черепного нерва
- родопсин
Молекула фотопигмента- обнаружена в фоторецепторах палочек
- стержневой фоторецептор
- один из двух типов рецепторных клеток сетчатки, специализированных для зрения при слабом освещении
- круглое окно
- мембрана, обозначающая конец барабанной лестницы
- мешок
- структура внутреннего уха, отвечающая за преобразование линейного ускорения в вертикальной плоскости
- барабанная лестница
- часть улитки, простирающаяся от верхушки до круглого окна
- лестница вестибюля
- часть улитки, простирающаяся от овального окна до верхушки
- склера
- белок глаза
- полукружные каналы
- структур внутреннего уха, отвечающих за передачу информации о вращательном движении
- сенсорная модальность
- особая система для интерпретации и восприятия раздражителей окружающей среды нервной системой
- соматоощущение
- общий смысл, связанный с модальностями, объединенными вместе как осязание
- особый смысл
- любая сенсорная система, связанная с определенной структурой органа, а именно обоняние, вкус, зрение, слух и равновесие
- спиральный ганглий
- расположение тел нейронов, передающих слуховую информацию, по восьмому черепному нерву
- стремя
- (также стремечко) косточка среднего уха, прикрепляющаяся к внутреннему уху
- стереоцилии
- массив отростков апикальной мембраны в волосковой клетке, которые передают движения при изгибании
- субмодальность
- специфическое чувство в более широком смысле, например, сладкое как часть вкусового ощущения или цвет как часть зрения
- верхняя косая
- экстраокулярная мышца, отвечающая за медиальное вращение глаза
- верхняя прямая мышца
- экстраокулярная мышца, отвечающая за взгляд вверх
- вкусовые рецепторы
- структур внутри сосочка на языке, содержащих клетки вкусовых рецепторов
- текториальная мембрана
- компонент кортиева органа, лежащий над волосковыми клетками, в который встроены стереоцилии
- терморецептор
- сенсорный рецептор, специализирующийся на температурных раздражителях
- топографический
- относительно позиционной информации
- трансдукция
- процесс преобразования раздражителя окружающей среды в электрохимические сигналы нервной системы
- блок
- хрящевая структура, действующая как шкив для верхней косой мышцы
- барабанная перепонка
- барабанная перепонка
- умами
- вкусовая субмодальность на чувствительность к концентрации аминокислот; также называется пикантным чувством
- ученый
- структура внутреннего уха, отвечающая за преобразование линейного ускорения в горизонтальной плоскости
- сосудистая оболочка
- средний слой глаза состоит в основном из соединительной ткани с обильным кровоснабжением
- вестибулярный ганглий
- расположение тел нейронов, передающих информацию о равновесии, вдоль восьмого черепного нерва
- тамбур
- в ухе, часть внутреннего уха, отвечающая за чувство равновесия
- внутреннее чувство
- чувство, связанное с внутренними органами
- зрение
- особое зрение, основанное на передаче световых раздражителей
- острота зрения
- свойство зрения, связанное с резкостью фокуса, которое изменяется в зависимости от положения сетчатки
- стекловидное тело
- вязкая жидкость, заполняющая заднюю камеру глаза
- волокна зонулы
- фиброзные связи между цилиарным телом и хрусталиком
Физиология, цветовосприятие — StatPearls
Натаниэль Пасмантер; Сунил Мунакоми.
Информация об авторе
Последнее обновление: 18 сентября 2021 г.
Введение
У людей восприятие и способность различать цвета опосредованы различными механизмами сетчатки, а также мозга. Понимание физиологических основ цветового зрения необходимо для выявления аномалий и разработки методов лечения. В этой статье мы рассмотрим клеточные и генетические механизмы, лежащие в основе восприятия цвета, и применим эти механизмы для характеристики дефектов цветового зрения и способов их лечения.
Проблемы, вызывающие озабоченность
Нарушение цветового зрения может быть результатом различных аномалий, как системных, так и специфических внутри зрительной системы. Дефекты в генах, отвечающих за зрительную трансдукцию, часто приводят к врожденному дефициту цветового зрения. Любые аномалии сетчатки, зрительного нерва, зрительного тракта и зрительной коры могут вызывать дефекты цветового зрения. Таким образом, системные заболевания, такие как диабет, могут изменить цветовое зрение, как и глазные заболевания, такие как глаукома и катаракта.
Клеточная
Сетчатка большинства млекопитающих содержит только два типа колбочек (дихроматы), чувствительных к коротким длинам волн (S-колбочки, максимально чувствительные в синей части зрительного спектра) и средним длинам волн (M-колбочки, максимально чувствительные к зеленой длине волны). легкий). У людей, как у трихроматов, есть дополнительная L-колбочка, которая чувствительна к длинноволновому свету в красной части зрительного спектра. Дифференциальная стимуляция этих колбочек создает цветовые оси между красно-зеленым и сине-желтым, что позволяет визуализировать смеси цветов, попадающих в диапазон зрительного спектра. Сине-желтая цветовая ось несколько неверна, поскольку она более конкретно относится к способности отличать синий от зеленого, а также желтый от красного.[2]
Первым этапом зрительной трансдукции является опосредованное светом конформационное изменение 11- цис -ретиналя, которое активирует ассоциированный опсин, который действует как рецептор, связанный с G-белком.
Развитие
Исследования показывают, что к двухмесячному возрасту у младенцев появляется функциональное цветовое зрение. Кроме того, они способны различать несколько оттенков независимо от яркости и функции палочек, хотя их зрение отличается от зрения взрослых. Уточнение и постоянное развитие цветового зрения недостаточно изучены, равно как и возраст пикового функционирования, но, вероятно, зависит от усиления путей колбочковых фоторецепторов в результате продолжительного зрительного опыта.
Вовлеченные системы органов
Нормальное цветовосприятие является функцией нервной системы, зависящей от зрительного преобразования и передачи информации в зрительную кору.
Функция
Считается, что эволюция трехцветного цветового зрения помогла ранним приматам отличать красные, оранжевые и желтые плоды от зеленой листвы.[2] У современных людей цветовое зрение играет жизненно важную роль в выполнении домашних дел, вождении автомобиля и многих других взаимодействиях с окружающей средой. Потеря функционального цветового зрения может ухудшить эти повседневные задачи, включая производительность труда, и даже может помешать выбору конкретной карьеры.
Механизм
Глаз формирует изображения на основе различий в коэффициенте отражения света от внешних объектов. Небольшие возмущения, напротив, обрабатываются через систему центр-объем, где яркость окружающего фона вычитается из центрального сигнала, выделяя локальные особенности центрального сигнала.
Колбочки передают визуальную информацию через парвоцеллюлярные слои в латеральное коленчатое ядро (LGN) таламуса. Нейроны в LGN обрабатывают величину вклада сигналов колбочек оппонента и продолжают передавать сигнал в первичную зрительную кору V1. Механизмы восприятия цвета за пределами LGN не так хорошо охарактеризованы. Тем не менее, исследования фМРТ предполагают, что в V1 происходит дополнительное разделение и обработка как цветовой яркости, так и цветового контраста, а также дополнительных экстрастриарных областей зрения. Исследования фМРТ путей, участвующих в восприятии и обработке цветов, указывают на потенциальный нисходящий механизм взаимодействия височно-теменной коры со зрительными областями V2/3.
Различные дополнительные сложные механизмы влияют на восприятие цвета. Оттенок, насыщенность и яркость как центральной, так и окружающей области сильно влияют на восприятие центрального цвета, так что один и тот же стимул, представленный на разных фонах, может восприниматься совершенно как разные цвета. Индивидуальные различия в обработке цвета также влияют на то, как эти факторы влияют на восприятие цвета.
Хроматическое зрительное восприятие, по-видимому, подвержено высокой степени нейронной пластичности; это, по-видимому, опосредовано средней яркостью фона и уравновешивающими средними хроматическими стимулами (восприятие нормального цвета фона корректируется, чтобы дать равные вклады противоположных цветовых осей).
Все цвета получаются путем аддитивного или субтрактивного смешивания трех основных цветов.
Сопутствующее тестирование
Наиболее распространенным методом диагностики дефектов цветового зрения являются «псевдоизохроматические» тесты пластин, такие как пластины Исихара, из-за простоты доступа и использования. Однако эти тесты часто не позволяют дифференцировать различные типы дисхроматопсии. Другой диагностический инструмент, заказ тестов, требует, чтобы пациенты сортировали разные диски в зависимости от прогрессии цвета. По сравнению с пластинчатыми тестами эти тесты, как правило, более чувствительны при обнаружении нарушений цветового зрения, а также лучше при постановке конкретных диагнозов. Дополнительные диагностические методы, представляющие интерес, включают тесты на соответствие, привлекательные из-за возможности их проведения и оценки на компьютере.
Патофизиология
Нарушения цветового зрения обычно подразделяются на врожденные и приобретенные формы. Врожденные нарушения цветового зрения возникают из-за дефектов генов, кодирующих опсины колбочек, генов, кодирующих экспрессию опсинов колбочек, и генов белков, участвующих в фототрансдукции (таких как субъединицы канала PDE и CNG). Такие дефекты имеют широкий спектр возможных исходов и могут характеризоваться различной скоростью прогрессирования, дегенерацией сетчатки и потерей остроты зрения. Дефицит гуанилатциклазы может привести к врожденному амаврозу Лебера и тяжелой потере зрения в раннем возрасте. Напротив, палочковые дистрофии, приводящие к пигментному ретиниту, могут вызывать поздние нарушения цветового зрения, поскольку дегенерирующие палочки влияют на структуру и функцию колбочек.
Клиническое значение
Дефекты, влияющие на определенные типы опсинов колбочек, имеют связанные префиксы – протан для L колбочек, дейтан для М колбочек и Тритан для S колбочек. Например, если опсин L колбочек присутствует, но ненормальный, пациент будет классифицирован как «протаномальный трихромат». Напротив, полная потеря опсина L колбочек приведет к «дихромату протанопа». Такие дихроматы не смогут различать красный и зеленые видят только функциональные сочетания двух оттенков. В то же время аномальная трихромазия может варьироваться по степени тяжести от практически нормального цветового зрения до функциональной дихроматии.
Заболевания, которые в первую очередь поражают богатое колбочками пятно, такие как возрастная дегенерация желтого пятна (AMD), могут демонстрировать раннюю потерю цветоразличения до появления обнаруживаемых изменений морфологии сетчатки или остроты зрения.
Глаукома и глазная гипертензия, заболевания, которые могут привести к компрессии зрительного нерва, также могут вызывать дефекты цветового зрения. Восприятие цвета может измениться до того, как будут обнаружены другие функциональные или структурные изменения, которые, по-видимому, в первую очередь влияют на сине-желтую ось.
Многие другие расстройства, как специфичные для зрительной системы, так и общие, связаны с нарушениями цветового зрения. Разрывы и отслойки сетчатки могут привести к потере цветового зрения, а также к нарушениям, поражающим зрительный нерв, включая наследственную оптическую невропатию, неврит зрительного нерва и сдавление зрительного нерва. У многих пациентов с диабетической ретинопатией также наблюдается нарушение цветового зрения. Многие лекарства имеют значение при дисхроматопсии, включая ингибиторы ФДЭ-5, хлорохин, этамбутол и дигоксин, а также воздействие окружающей среды, такое как воздействие УФ-излучения, промышленных химикатов и гипоксии.
Генная терапия является перспективным направлением лечения многих врожденных дистрофий сетчатки. Многие генетические нарушения, присутствующие у нечеловеческих животных, аналогичны состояниям, наблюдаемым у людей. Трансляционная терапия замены гена, обычно опосредованная рекомбинантным аденоассоциированным вирусом (rAAV), оказалась успешной при лечении множественных врожденных дефектов, влияющих на цветовое зрение. Генетические дефекты RPE65, которые влияют на способность пигментного эпителия сетчатки перерабатывать ретиниловые эфиры и могут вызывать врожденный амавроз Лебера, успешно лечили с помощью генной терапии на моделях собак. С тех пор лечение было распространено на людей, что привело к первому одобренному FDA генно-терапевтическому лечению наследственной дистрофии сетчатки. Другие многообещающие методы лечения включают генную терапию врожденной ахроматопсии CNGB3, противодействующую отсутствию субъединицы канала CNG в колбочках, которая была успешной у собак [15], и генную терапию красно-зеленого дальтонизма у беличьих обезьян [16].
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Ссылки
- 1.
Симунович М.П. Приобретенный дефицит цветового зрения. Сурв Офтальмол. 2016 март-апрель;61(2):132-55. [PubMed: 26656928]
- 2.
Роу М.Х. Трихроматическое цветовое зрение у приматов. Новости физиол. 2002 июнь; 17:93-8. [PubMed: 12021378]
- 3.
Neitz M, Neitz J. Молекулярная генетика цветового зрения и дефектов цветового зрения. Арка Офтальмол. 2000 май; 118(5):691-700. [PubMed: 10815162]
- 4.
Браун А.М., Линдси Д.Т. Цветовое зрение и цветовые предпочтения младенцев: дань уважения Давиде Теллер. Vis Neurosci. 2013 ноябрь; 30 (5-6): 243-50. [PubMed: 23879986]
- 5.
Сугита Ю. Опыт в раннем детстве необходим для восприятия цвета. Карр Биол. 2004 г., 27 июля; 14 (14): 1267-71.
- 6.
Стюард Дж.М., Коул Б.Л. Что люди с дефектами цветового зрения говорят о повседневных задачах? Optom Vis Sci. 1989 г., май; 66 (5): 288–95. [PubMed: 2787492]
- 7.
Шепли Р., Хокен М. Нейронные механизмы восприятия цвета в первичной зрительной коре. Курр Опин Нейробиол. 2002 авг; 12 (4): 426-32. [PubMed: 12139991]
- 8.
Тинг Сиок В., Кей П., Ван В.С., Чан А.Х., Чен Л., Люк К.К., Хай Тан Л. Языковые области мозга участвуют в восприятии цвета. Proc Natl Acad Sci U S A. 200919 мая; 106(20):8140-5. [Бесплатная статья PMC: PMC2688888] [PubMed: 19416812]
- 9.
Horwitz GD. Что исследования макак рассказали нам о человеческом цветовом зрении. Неврология. 2015 18 июня; 296: 110-5. [Бесплатная статья PMC: PMC4401582] [PubMed: 25445192]
- 10.
Lotto RB, Purves D. Эмпирическая основа восприятия цвета. Сознательное Познание. 2002 Декабрь; 11 (4): 609-29.
- 11.
Neitz J, Carroll J, Yamauchi Y, Neitz M, Williams DR. Восприятие цвета опосредовано пластическим нейронным механизмом, который регулируется у взрослых. Нейрон. 2002 15 августа; 35 (4): 783-92. [PubMed: 12194876]
- 12.
Вемала Р., Шивапрасад С., Барбур Дж.Л. Выявление ранней потери цветового зрения при возрастной макулодистрофии — с акцентом на друзы и ретикулярные псевдодрузы. Invest Ophthalmol Vis Sci. 01 мая 2017 г .; 58 (6): BIO247-BIO254. [PubMed: 28846119]
- 13.
Папаконстантину Д., Георгалас И., Каланцис Г., Кармирис Э., Кутсандреа С., Диагуртас А., Ладас И., Георгопулос Г. Приобретенное цветовое зрение и дефекты поля зрения у пациентов с глазной гипертензией и ранняя глаукома. Клин Офтальмол. 2009 г.;3:251-7. [Бесплатная статья PMC: PMC2709000] [PubMed: 19668575]
- 14.
Миральди Утц В., Кусса Р.Г., Антаки Ф., Трабулси Э.И. Генная терапия заболеваний сетчатки, связанных с RPE65.
- 15.
Komáromy AM, Alexander JJ, Rowlan JS, Garcia MM, Chiodo VA, Kaya A, Tanaka JC, Acland GM, Hauswirth WW, Aguirre GD. Генная терапия восстанавливает функцию колбочек при врожденной ахроматопсии. Хум Мол Жене. 2010 01 июля; 19(13): 2581-93. [Бесплатная статья PMC: PMC2883338] [PubMed: 20378608]
- 16.
Mancuso K, Hauswirth WW, Li Q, Connor TB, Kuchenbecker JA, Mauck MC, Neitz J, Neitz M. Генная терапия для красно-зеленых дальтонизм у взрослых приматов. Природа. 2009 08 октября; 461 (7265): 784-7. [Бесплатная статья PMC: PMC2782927] [PubMed: 19759534]
Возможная физиологическая основа прерывистости сознания
Введение
Идея о том, что сознание человека не является непрерывным, а представляет собой серию дискретных фрагментов или моментов восприятия имеет долгую историю. Впервые оно появилось почти 2000 лет назад в виде буддийского учения о кшаникаваде (Пруден, 1988; Швейцер, 1994).
Пурвс и его коллеги сообщили, что иллюзия, напоминающая иллюзию, которая заставляет вращающиеся колеса телеги останавливаться и двигаться назад в старых ковбойских фильмах, также возникает в реальной жизни при естественном освещении. В фильмах иллюзия возникает из-за алиасинга или недостаточной дискретизации движения колеса, вызванного тем фактом, что немые фильмы воспроизводились со скоростью всего 16 кадров в секунду (т.
Однако исследования более физиологического характера — например, сообщения Леманна и его коллег о том, что данные ЭЭГ возникают в виде серии пространственно определенных «микросостояний» (Lehmann, 1971; Lehmann et al., 1987, 1998; Lehmann and Koenig , 1997) – также убедительно свидетельствуют о прерывности сознания. О подобных дискретных состояниях мозга сообщили Фриман и его коллеги (Freeman and Rogers, 2002; Freeman et al., 2003a,b; Freeman, 2004a,b), которые показали, что аналитическая мощность (AP) ЭЭГ с полосовой фильтрацией демонстрирует серии локальных минимумов или «нулевых пиков», при которых аналитическая фаза временного ряда ЭЭГ резко скачкообразно и относительно постоянный пространственный электромагнитный паттерн, наблюдаемый в любой данной области мозга, меняется на другой пространственный паттерн. Эти результаты привели Фримена (2006) к гипотезе о том, что переходы состояний, происходящие при минимумах ПД, могут обеспечить физиологическую основу для фреймов кинематографического сознания.
С небольшой модификацией, чтобы подчеркнуть разграничение кадров только глубокими минимумами в AP, эта гипотеза о нулевых пиках-затворах четко вписывается в теорию электромагнитного поля сознания (Pockett, 1999, 2000, 2002, 2007, 2011, в Нажмите). Теория электромагнитного поля сознания предполагает, что сознательные переживания идентичны определенным электромагнитным паттернам, генерируемым активностью мозга. Это означает, что периоды, когда электромагнитная активность, генерируемая мозгом, падает ниже некоторого порогового уровня, должны коррелировать с периодами, когда сознание выходит из строя — именно это и предлагает модифицированная гипотеза нулевых всплесков как затвора.
Идея о том, что электромагнитная активность должна превышать некоторое минимальное значение, чтобы обслуживать сознание, уже получила некоторую эмпирическую поддержку в нейромагнитных экспериментах Тонони и др. (1998). По словам этих авторов «1. Нервные ответы на соперничающие зрительные стимулы возникали в большом количестве областей коры как тогда, когда испытуемый сознательно воспринимал стимулы, так и когда он их не воспринимал.
На данном этапе важно прояснить, что минимальное количество электромагнитной активности предлагается более широкой теорией электромагнитного поля сознания и более узкой нулевой гипотезой о всплесках-затворах как необходимое , но недостаточное для сознания. Самая последняя формулировка теории электромагнитного поля (Pockett, 2011, в печати) также предлагает дополнительный фактор как необходимый, но недостаточный для сознания: особый паттерн электрического заряда, генерируемого LFP, в радиальном направлении, перпендикулярном поверхности мозг. Эта картина не может быть измерена с помощью поверхностных электродов, подобных тем, которые использовались в настоящем исследовании, которые могут только обеспечить измерение общей амплитуды электрических переходных процессов, происходящих в пределах нескольких миллиметров в любом направлении от каждого электрода, но его характеристика радиальной картины важна.
Гипотеза нулевых шипов как затвора относится только к одному из ряда факторов, которые накладывают ограничения на сознание. В нем говорится, что при отсутствии достаточной общей электромагнитной амплитуды не имеет значения, расположена ли та амплитуда, которая у вас есть, в правильном пространственном паттерне или нет — вы не можете иметь сознание. Конечно, обратное не подразумевается: существует множество возможных ситуаций, когда электромагнитная амплитуда слишком мала для поддержания сознания, и очень немногие из них когда-либо смогут генерировать электромагнитные паттерны, способные было бы достаточно для сознания.
При этом основная цель работы, представленной здесь, состоит в том, чтобы проверить правильность гипотезы нулевых шипов как затвора. Согласно классическому труду Поппера (1995) « Logik der Forschung », невозможно доказать научную гипотезу — лучшее, что можно сделать, — это не опровергнуть ее. Один из способов попытаться фальсифицировать гипотезу нулевых всплесков как затвора состоит в том, чтобы измерить частоту глубоких локальных минимумов в ПД данных ЭЭГ и сравнить ее с частотой предполагаемых фреймов сознания, о которых сообщается в психологической литературе. Если два набора частот окажутся радикально разными, гипотеза нулевых пиков как затвора будет фальсифицирована. Если бы существовала хорошая корреляция между двумя наборами частот, идея о том, что сознание возникает в кадрах, и гипотеза о том, что минимумы ПД служат для определения этих кадров, были бы поддержаны — хотя, конечно, не доказаны.
Однако есть одна большая трудность, связанная с проведением такого теста. Проблема в том, что Райс (1952) математически показал, что частота локальных минимумов в АП белого шума, отфильтрованного идеальным полосовым фильтром, линейно связана с полосой пропускания фильтра. В частности (Rice, 1952, стр. 9):
В большинстве случаев ЭЭГ больше напоминает коричневый или черный шум, чем белый шум, проанализированный Райсом, но и Freeman (2009), и Ruiz et al. (2010) эмпирически проверили взаимосвязь Райса и обнаружили, что, по крайней мере, для узких полос частот данные ЭЭГ удивительно хорошо соответствуют уравнению 1. Это затрудняет чистую эмпирическую проверку гипотезы об нулевых выбросах как затворе, потому что далеко не ясно, как (или даже являются ли) несемплированные электромагнитные продукты мозговой активности по своей природе ограниченными диапазонами. Большое количество данных свидетельствует о том, что различные полосы пропускания в пределах гамма-диапазона связаны с различными видами сознания (например, Таллон-Бодри и Бертран, 19).
Здесь мы решаем эту проблему, измеряя частоту глубоких минимумов АД в данных ЭКоГ у субъектов в сознании и без сознания, в расширенном ряду полос пропускания и для нескольких различных значений требуемого минимального АД. Затем мы сравниваем результаты этих измерений с длительностью предполагаемых фреймов сознания, описанных в психологической литературе.
Материалы и методы
Данные
Данные были зарегистрированы у четырех взрослых субъектов, которым были имплантированы внутричерепные электроды с клинической целью локализации эпилептических очагов. Испытуемые (три женщины) были в возрасте от 23 до 38 лет. У всех была рефрактерная к лечению локализованная эпилепсия, проявлявшаяся сложными парциальными и вторично-генерализованными припадками, и все они были кандидатами на операцию по поводу эпилепсии. Субъектов обследовали в Региональном центре эпилепсии Вашингтонского университета, расположенном в Медицинском центре Харборвью в Сиэтле, штат Вашингтон. Первоначальная неинвазивная оценка включала подробный анамнез, клиническое обследование, стандартную ЭЭГ, магнитно-резонансную томографию, нейропсихологическую оценку и долгосрочные ЭЭГ-видеомониторные исследования скальпа. После определения того, что инвазивные записи ЭЭГ необходимы для локализации приступов, каждому субъекту были выполнены долгосрочные внутричерепные субдуральные ЭЭГ-видеомониторные записи, которые включали размещение субдуральных 64-контактных электродов с сеткой 8 × 8 над выпуклостью интересующего полушария головного мозга.
Рисунок 1. Расположение записывающих электродов .
Данные были записаны с частотой дискретизации 500 Гц для двух испытуемых и 2000 Гц для двух других, с аналоговыми полосами пропускания 1–100 Гц.
Для сравнения с этими данными степенной искусственный шум был сгенерирован путем свертки случайного шума с фильтром 1/ f β , где β = 0 для белого шума, 1 для розового шума, 2 для коричневого шума и 3 для черного шума.
Обработка сигналов
Обработка сигналов выполнена в Matlab. Данные ЭКоГ визуально проверяли с помощью графиков временных рядов на каждом электроде, а случайный канал, содержащий значительные артефакты записи, отбрасывали. Остальные каналы были преобразованы в единое пространственное среднее по ансамблю (SEA) путем получения среднего значения всех каналов в каждый момент времени. Режекторный фильтр был применен для удаления всплеска линейного шума 60 Гц путем преобразования Фурье SEA, замены значения 60 Гц средним значением в двух соседних частотных элементах и последующего применения обратного преобразования Фурье.
Спектры плотности мощности ( P ) как функция частоты ( f ) были рассчитаны для данных SEA с надрезом с использованием многоконусного метода Томпсона (Matlab «pmtm.m»). Затем спектры мощности для всех четырех испытуемых усреднялись, а результат наносился на логарифмическую ось. Наклон этого графика был определен путем подгонки линии линейной регрессии к значениям log( P i ) и log( f i ) ( i = 1, 2, …, n ) для выбранного частотного диапазона с использованием схемы взвешивания, предназначенной для компенсации того факта, что на логарифмическом графике экспоненциально увеличивается количество точек на более высоких частотах.
, где вес w i равен
, а σ i является SE для логарифма средней плотности мощности, полученной из четырех отдельных логарифмических графиков. Фактор влияет на компенсацию неравномерно расположенных спектральных компонентов на логарифмическом графике. В случае постоянного σ i это гарантировало бы, что общий вес для точек в заданном диапазоне частот будет пропорционален изменению log( f ) в этом диапазоне (и, следовательно, вес за декаду будет постоянный).
SE в наклоне был рассчитан с использованием аналитического выражения для наклона β, преобразованного в виде линейной комбинации плотностей мощности и объединения SE компонентов плотности мощности в предположении, что нет корреляции ошибок между отдельными компонентами плотности мощности .
Затем временные ряды SEA с надрезами были отфильтрованы в серию все более широких полос пропускания вокруг выбранной центральной частоты.
Глубокие минимумы были выбраны в результирующем временном ряду AP, сначала выбирая все минимумы (путем сравнения каждой точки данных во временном ряду с точками данных по обе стороны от него), а затем выбирая только те минимумы, амплитуда которых упала ниже заданный порог, который выражался в процентах от средней амплитуды всех минимумов в этом наборе данных. В некоторых случаях этот алгоритм был расширен, чтобы игнорировать больший из двух минимумов, заключенных в скобки пика, который сам не смог подняться выше порога.
Результаты
Рисунок 2 служит интуитивным насосом для демонстрации значения АР. Аналитический сигнал реальной последовательности данных представляет собой комплексное преобразование, имеющее действительную часть, идентичную исходным данным, и мнимую часть, содержащую преобразование Гильберта, представляющее собой версию исходного сигнала, сдвинутую по фазе на 90°. Аналитическая мощность описывает огибающую квадратов (действительных + мнимых) частей аналитического сигнала.
Рисунок 2. Зависимость аналитической мощности от временного ряда .
На рис. 3 показаны спектры плотности мощности необработанной ЭКоГ бодрствования (рис. 3А) и необработанной ЭКоГ, зарегистрированной во время постиктального состояния бессознательного состояния (рис. 3В). Средний наклон логарифмического графика для всех субъектов на рисунке 3A составляет -2,93 ± 0,09 (среднее значение ± стандартное отклонение). Наклон большей части логарифмического графика на рисунке 3B составляет 0,07 ± 0,05.
Рисунок 3. Спектры плотности мощности для (A) ЭКоГ бодрствования и (B) ЭКоГ в бессознательном состоянии . Толстые линии: среднее значение по всем четырем предметам. Тонкие линии: плюс и минус одно стандартное отклонение от среднего.
На рисунке 4 показаны временные ряды AP сегментов (рисунок 4A) бодрствования и (рисунок 4B) постиктальных данных ЭКоГ с кружками вокруг минимумов, выбранных в нашем анализе.
Рисунок 4. Минимумы, выбранные в сегментах (A) бодрствования и (B) ЭКоГ в постиктальном бессознательном состоянии одного и того же субъекта . Порог, ниже которого минимумы должны опускаться, чтобы их можно было подсчитать (горизонтальная линия), установленный на уровне 30% от средней амплитуды для всех минимумов в этом наборе данных.
Еще одно наблюдение заключается в том, что примерно в моменты времени 0,64 и 0,73 с на рис. 4А выбираются два локальных минимума, которые (а) происходят очень близко друг к другу по времени и (б) заключают в себе максимум или пик, который сам по себе не может подняться выше установленного порог (горизонтальная линия). На том основании, что такие пики не могут служить сознанию больше, чем минимумы, которые их очерчивают, некоторые анализы, представленные ниже, были повторены с использованием алгоритма выбора минимумов, предназначенного для игнорирования большего из двух таких минимумов. Это включало исключительно сложный алгоритм, который, как правило, заставлял Matlab выдавать ошибки нехватки памяти, и, поскольку было обнаружено, что он практически не влияет на результаты дальнейшего анализа, они в основном выполнялись с использованием более простого кода выбора минимумов, описанного в разделе «Материалы и методы».
На рис. 5 показано, как эти наблюдения отражаются на «кривых Райса» данных бодрствования и постиктальных данных.
Рисунок 5. Кривые Райса для данных ЭКоГ в бодрствовании и в постприпадочном бессознательном состоянии . Минимумы аналитической мощности с порогом, установленным на уровне 30% (как на рисунке 4). Центральная частота всех полос пропускания составляет 50 Гц, поэтому полоса пропускания 40 Гц по горизонтальной оси соответствует полосе пропускания 30–70 Гц, а полоса пропускания 60 Гц соответствует полосе пропускания 20–80 Гц.
Таблица 1 иллюстрирует влияние изменения настройки порога на частоту, при которой происходит насыщение минимума ПД, показывая индивидуальные результаты для каждого субъекта. Можно видеть, что стандартное отклонение 20 наблюдений в области плато всех кривых Райса относительно невелико, что указывает на то, что плато довольно плоские. Для каждого субъекта среднее значение плато увеличивается предсказуемым образом по мере повышения порога, что позволяет подсчитывать больше минимумов. Для любого субъекта и условий частота минимумов ПД в SEA (показана) существенно не отличалась от таковой в данных случайного одиночного электрода в середине основной сетки (9).0011 p > 0,05, t — тест).
Таблица 1 . Влияние настройки порога на частоту плато минимумов аналитической мощности при ЭКоГ бодрствования .
Искусственный шум, построенный таким образом, чтобы спектры плотности мощности имели логарифмические наклоны, аналогичные нашим данным ЭКоГ, давали кривые Райса, аналогичные кривым данных.
Обсуждение
Заявленная цель этой серии анализов состояла в том, чтобы исследовать идею о том, что сознание возникает во фреймах, потому что ПД происходит в фреймах, путем сравнения наших физиологических измерений АД с психологическими данными в литературе о фреймах сознания.
Таблица 2 обобщает психологические данные, имеющиеся в литературе. В большинстве экспериментов сообщается, что зрение возникает в кадрах продолжительностью от 50 до 100 мс, т. Е. При частоте кадров 10–20 Гц. В таблице 1 приведены частоты, при которых на нашей ЭКоГ бодрствования наблюдается плато минимума глубокого АД, с центральной частотой кривой Райса 50 Гц (средняя гамма, которая считается важным частотным диапазоном в отношении сознания) и порогом, ниже которого АД минимум должен был упасть, чтобы его можно было считать установленным на уровне от 10 до 40% от среднего минимума в наборе данных. Частота плато минимумов АД составляет от 8,4 ± 1,3 Гц (для субъекта 1 с минимальным порогом АД, установленным на уровне 10%), до 23,4 ± 1,8 Гц (для субъекта 4 с минимальным порогом АД, установленным на уровне 40%).
Таблица 2 . Частоты фреймов зрительного сознания в психологической литературе .
Эти результаты получены из пространственно усредненной ЭКоГ, записанной у испытуемых, которые не выполняли зрительную задачу, поставленную экспериментатором, а просто спокойно отдыхали с открытыми глазами. Таким образом, один вопрос заключается в том, были ли измеренные нами минимумы АД просто артефактами пространственного усреднения, а другой вопрос заключается в том, действительно ли мы измеряли нейронные корреляты зрительного сознания.
Что касается первого вопроса, отдельные минимумы, безусловно, выпадали в разное время в SEA и в данных с отдельных электродов, но общие значения, при которых минимумы выходили на плато в двух наборах данных, существенно не отличались.
Второй вопрос настоящего анализа заключается в том, относятся ли наши данные конкретно к зрительному сознанию. Этот вопрос можно разделить на две части: во-первых, характерны ли данные для сознания, и, во-вторых, характерны ли они для зрительного сознания.
Первая часть этого вопроса посвящена нашему сравнению данных бодрствующих и постиктально бессознательных субъектов. Мы выбрали постиктальное состояние как наиболее безопасный пример общей бессознательности, потому что все другие предположительно бессознательные состояния не были представлены доступными нам данными — и действительно не могут быть надежно представлены какими-либо данными.
Бессознательное состояние обычно предполагается, если выполняются два критерия: субъект (а) нереактивен (не реагирует на внешние раздражители) и (б) амнезиачен (не помнит впоследствии, что был в сознании в рассматриваемое время). Что касается анестезии, то эти критерии на первый взгляд могут выполняться в большинстве случаев, но из этого нельзя разумно заключить, что в любой момент времени хирургическая анестезия обязательно приводила к потере сознания.
Другим важным состоянием, при котором можно заподозрить бессознательное состояние, является медленный сон. Но теперь известно, что медленный сон не обязательно проходит без сновидений (Hobson et al.
Напротив, постиктальное состояние, возникающее в течение нескольких минут после прекращения генерализованного тонико-клонического припадка, представляет собой состояние, при котором пациент не реагирует, амнезиачен и выдает данные ЭКоГ со спектрами мощности, полностью отличающимися от тех, которые вырабатываются мозгом в бодрствующем состоянии. (см. рис. 3A, B) или мозга в медленном сне (Freeman et al., 2006), что маловероятно, что он поддерживает какую-либо скрытую форму сознания. Когда испытуемый находится в этом состоянии, на рис. 5 видно, что на кривой Райса нет плато. Полное отсутствие плато кривой Райса в данных от бессознательных субъектов подтверждает идею о том, что плато кривой Райса может быть важным моментом в определении частоты кадров сознания.
Что касается вопроса о том, относятся ли наши данные конкретно к зрительному сознанию: наши испытуемые определенно не выполняли зрительных задач, поставленных экспериментатором.
Следовательно, мы утверждаем, что наши данные согласуются с гипотезой о том, что
(i) Некоторое минимальное значение АР – по нашим данным, вероятно, более 50 мкВ 2 /Гц – необходимо (хотя и недостаточно) для зрительного сознания и
(ii) Поскольку ПД, генерируемая бодрствующим мозгом, периодически падает ниже этого минимума с интервалами, которые в среднем не короче 50–100 мс, зрительное сознание возникает в кадрах или фрагментах длительностью не менее 50–100 мс.
Другими словами, глубокие минимумы ПД мозга вполне могут действовать как затвор в кинематографической модели сознания.
На вопрос, возникает ли эта связанная с АП дискретность сознания из-за какого-то специфически биологического фактора или же исключительно как следствие физических свойств сигналов 1/ f , отвечает наше исследование искусственного 1/ f шума. Повторение наших анализов искусственного шума показало, что искусственные сигналы 1/ f , построенные так, чтобы иметь те же спектры мощности, что и биологические данные, дают те же кривые Райса, что и биологические данные.
1. Локальные минимумы ПД встречаются во всех 1/ f (степенных) спектрах, независимо от того, имеют ли генераторы этих спектров какое-либо отношение к сознанию. Локальные минимумы — это просто физическая характеристика спектра 1/ f .
2. Все данные ЭЭГ имеют спектры 1/ f . В данных, полученных от субъектов, находящихся в сознании, показатель степени f находится между 2 и 3. В данных, полученных от наших субъектов, находящихся в бессознательном состоянии, показатель степени f в большей части частотного спектра практически равен 0. Но в обоих случаях спектры равны 1/. ф .
3. Следовательно, временные ряды ПД всех данных ЭЭГ – независимо от того, лежат ли они в основе сознания или нет – имеют локальные минимумы. AP мозга происходит в кадрах.
4. Следовательно, ЕСЛИ (как это предлагается как теорией электромагнитного поля сознания, так и гипотезой нулевых всплесков как жалюзи) для сознания необходимо определенное количество генерируемой мозгом ПД, сознание должно возникать в кадрах.
5. Нелогично предполагать обратное: что все данные с минимумами АД должны иметь какое-то отношение к сознанию.
Этот ответ подтверждается и расширяется при более внимательном рассмотрении кривых Райса, созданных различными показателями степени f в данных сознательного и бессознательного мозга. Можно видеть (рис. 5; табл. 1), что продолжительность кадров ПД в сознательном мозге достигает насыщения или плато примерно при той же продолжительности, о которой сообщалось в ряде более ранних психологических исследований кадров сознания (табл. 2). Психологические исследования более или менее согласны с тем, что сознание приходит в кадрах длительностью в диапазоне 50–100 мс. Кривые Райса в настоящем исследовании плато при длительности кадра около 50–100 мс. Это соответствие обеспечивает поддержку не только гипотезы нулевых всплесков как затвора, но и более широкой теории сознания с электромагнитным полем, обе из которых предсказывают, что сознание должно возникать в кадрах, потому что AP происходит в кадрах.
Напротив, когда данные, проанализированные на кривых Райса, поступают из бессознательного мозга, плато длительности кадра не наблюдается. Поскольку данные фильтруются во все более широких полосах пропускания, продолжительность кадров точки доступа становится все короче. Нет очевидного соответствия между продолжительностью кадров AP и продолжительностью ранее зарегистрированных кадров сознания. Это, очевидно, гораздо более слабый аргумент в пользу исследуемой теории и гипотезы, но несоответствие как минимум интересное.
Приведенный выше ответ на вопрос «почему сознание прерывисто?» вызывает ряд дополнительных вопросов. Например, было бы неплохо в первую очередь знать механизм, с помощью которого мозг генерирует спектры 1/ f .
На первый вопрос ответ просто неизвестен. Степенной закон (он же 1/ f ) спектры широко распространены в природе. Они порождены всевозможными процессами, которые на первый взгляд совершенно не связаны между собой. Никто не понимает почему. На самом деле, как выразились Уорд и Гринвуд (2007), «повсеместное распространение шума 1/ f было описано как одна из старейших загадок современной физики и науки в целом». Конечно, авторы настоящей статьи пытались, но не смогли понять синаптические механизмы, с помощью которых генерируются, в частности, спектры 1/ f ЭЭГ. Эта неудача, вероятно, неудивительна, учитывая, что точные синаптические механизмы, с помощью которых генерируется любой данный образец ЭЭГ, в настоящее время остаются неясными.
Что касается последнего вопроса, то ситуация несколько лучше.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Allport, DA (1968). Фемоменальная одновременность и гипотеза перцептивного момента.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Эндрюс, Т., и Пурвс, Д. (2005). Иллюзия колеса телеги в непрерывном свете. Познание тенденций. науч. 9, 261–263.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Эндрюс, Т.Дж., Первес, Д., Симпсон, В.А., и Ван Руллен, Р. (2005). Колеса продолжают вращаться: ответ Холкомбу и др. Тенденции Cogn. науч. 9, 560–561.
Полный текст CrossRef
Augenstine, LG (1955). «Доказательства периодичности выполнения задач человеком», в Теория информации в психологии: проблемы и методы , изд. Х. Квастлер (Гленко, Иллинойс: Free Press), 208–231.
Баарс, Б.Дж. (1988). Когнитивная теория сознания . Нью-Йорк, штат Нью-Йорк: Издательство Кембриджского университета.
Буш, Н. А., и Ван Руллен, Р. (2010). Спонтанные колебания ЭЭГ выявляют периодическую выборку зрительного внимания.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Крик, Ф. (1994). Удивительная гипотеза: научный поиск души . Нью-Йорк: Саймон и Шустер.
Crick, FHC, and Koch, C. (2003). Каркас для сознания. Нац. Неврологи. 6, 119–126.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Дехане, С., Чанже, Ж.-П., Наккаш, Л., Сакур, Дж., и Сержент, К. (2006). Сознательная, предсознательная и подсознательная обработка: проверяемая таксономия. Тенденции Cogn. науч. 10, 204–211.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Эфрон, Р. (1970). Минимальная продолжительность восприятия. Нейрофизиология 8, 57–63.
Фриман, В. Дж. (2004a). Происхождение, структура и роль фоновой активности ЭЭГ. Часть 1 Аналитическая амплитуда. клин. Нейрофизиол. 115, 2077–2088.
Полный текст CrossRef
Freeman, WJ (2004b).
Полный текст CrossRef
Freeman, WJ (2006). Кинематографическая гипотеза корковой динамики в восприятии. Междунар. Ж. Психофизиол. 60, 149–161.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Freeman, WJ (2009). Глубокий анализ восприятия через динамические структуры, возникающие в корковой активности из саморегулируемого шума. Познан. Нейродин. 3, 105–116.
Опубликован Аннотация | Опубликован полный текст | CrossRef Full Text
Freeman, WJ, Burke, B.C., and Holmes, MD (2003a). Апериодическая переустановка фазы в скальповой ЭЭГ бета-гамма-осцилляций переходами состояний с частотой альфа-тета. Гул. Карта мозга. 19, 248–272.
CrossRef Full Text
Freeman, WJ, Holmes, MD, Burke, B.C., and Vanhatalo, S. (2003b). Пространственные спектры ЭЭГ и ЭМГ скальпа бодрствующих людей.
CrossRef Full Text
Freeman, WJ, Holmes, M.D., West, G.A., and Vanhatalo, S. (2006). Динамика неокортекса человека, оптимизирующая его стабильность и гибкость. Междунар. Дж. Интелл. Сист. 21, 881–901.
Полный текст CrossRef
Freeman, WJ, and Rogers, LJ (2002). Точное временное разрешение аналитической фазы выявляет эпизодическую синхронизацию переходов состояний в гамма-ЭЭГ. J. Нейрофизиол. 87, 937–945.
Опубликован Аннотация | Полный текст в публикации
Гаттасс, Р., Насименто-Сильва, С., Соарес, Дж. Г. М., Лима, Б., Янсен, А. К., Диого, А. К. М., Фариас, М. Ф., Маркондес, М., Ботельо, Э. П., Мариани, О. С. , Аззи, Дж., и Фиорани, Ф.М. (2005). Корковые зрительные зоны у обезьян: расположение, топография, связи, столбцы, пластичность и корковая динамика. Филос. Транс. Р. Соц. В 360, 709–731.
CrossRef Full Text
Ghoneim, MM (2000). Сознание во время анестезии.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Хобсон, Дж. А., Пейс-Шотт, Э. Ф., и Стикголд, Р. (2000). Сновидения и мозг: к когнитивной нейробиологии состояний сознания. Поведение. наук о мозге. 23, 793–1121.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Холкомб, А.О., Клиффорд, К.В.Г., Иглман, Д.М., и Пакарян, П. (2005). Иллюзорное реверсирование движения в соответствии с датчиками движения. Тенденции Cogn. науч. 9, 559–560.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Джеймс, В. (1890 г.). Принципы психологии . Лондон: Макмиллан.
Клайн, К., Холкомб, А.О., и Иглман, Д.М. (2004). Иллюзорное изменение направления движения вызвано соперничеством, а не моментальными снимками поля зрения. Видение Рез. 44, 2653–2658.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Крейман Г.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Леманн, Д. (1971). Многоканальная топография альфа-полей ЭЭГ человека. Электроэнцефалогр. клин. Нейрофизиол. 31, 439–449.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Леманн, Д., и Кениг, Т. (1997). Пространственно-временная динамика альфа-электрических полей мозга и когнитивных режимов. Междунар. Ж. Психофизиол. 26, 99–112.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Леманн, Д., Одзаки, Х., и Пал, И. (1987). Серия альфа-карт ЭЭГ: микросостояния мозга с помощью пространственно-ориентированной адаптивной сегментации. Электроэнцефалогр. клин. Нейрофизиол. 67, 271–288.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Леманн Д.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Леопольд, Д. А., и Логотетис, Н. К. (1996). Изменения активности в ранней зрительной коре отражают восприятие обезьян во время бинокулярного соперничества. Природа 379, 549–553.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Логотетис, Н.К., и Шалл, Дж.Д. (1989). Нейрональные корреляты субъективного зрительного восприятия. Наука 245, 761–763.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Мэтьюсон, К. Э., Граттон, Г., Фабиани, М., Бек, Д. М., и Ро, Т. (2009). Видеть или не видеть: альфа-фаза престимула предсказывает визуальное осознание. J.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Мерфи, Г.С., Сокол, Дж.В., Вендер, Дж.С., Мэримонт, Дж.Х., и Аврам, М.Дж. (2002). Использование нервно-мышечных блокаторов в кардиохирургии у взрослых: результаты национального почтового опроса. Анест. аналг. 95, 1534–1539.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Пакарян, П., и Ясами, М. Т. (2003). Иллюзия колеса телеги при постоянном освещении: реальность или иллюзия? Восприятие 32, 1307–1310.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Pockett, S. (1999). Анестезия и электрофизиология слухового сознания. В сознании. Познан. 8, 45–61. Доступно по адресу: http://www.psych.auckland.ac.nz/webdav/site/psych/shared/about/our-people/documents/sue-pockett/TheNatureofConsciousnessAHypothesis.pdf
Pockett, S. (2000). Природа сознания: гипотеза .
Покет С. (2002). Трудности с электромагнитно-полевой теорией сознания. Дж. В сознании. Стад. 9, 51–56.
Покет С. (2003). Сколько длится «сейчас»? Феноменология и кажущееся настоящее. Феноменол. Познан. науч. 2, 55–68.
Полный текст CrossRef
Pockett, S. (2007). Трудности с теорией электромагнитного поля сознания: обновление. Нейроквантология 3, 271–275.
Покет С. (2011). Инициация преднамеренных действий и теория электромагнитного поля сознания. Humana Mente 15, 159–175.
Покет С. (в печати). Теория электромагнитного поля сознания: проверяемая гипотеза о характеристиках сознательных полей в отличие от бессознательных. Дж. В сознании. Шпилька .
Pockett, S., Bold, G.E.J., and Freeman, W.J. (2009). Синхронность ЭЭГ при выполнении перцептивно-когнитивной задачи: распространенная фазовая синхронность на всех частотах. клин. Нейрофизиол. 120, 695–708.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Покетт, С., и Холмс, доктор медицины (2009). Внутричерепные спектры мощности ЭЭГ и фазовая синхронизация в сознании и бессознательном состоянии. В сознании. Познан. 18, 1049–1055.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Поппер, К. Р. (1995). Логика научных открытий . Лондон: Рутледж.
Пруден, Л. М. (1988). Абхидхармакошабхасьям . Беркли, Калифорния: Asian Humanities Press.
Первес Д., Пайдарфар Дж. А. и Эндрюс Т. Дж. (1996). Иллюзия колеса телеги в кино и реальности. Проц. Натл. акад. науч. США 93, 3693–3697.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Кирога, Р. К., Редди, Л., Крейман, Г., Кох, К., и Фрид, И. (2005). Инвариантное визуальное представление одиночными нейронами в человеческом мозгу. Природа 435, 1102–1107.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Rice, SO (1952).
Руис Ю., Покетт С., Фриман В.Дж., Гонсалес Э. и Гуанг Л. (2010). Метод изучения глобальных пространственных паттернов, связанных с сенсорным восприятием, на ЭЭГ скальпа. J. Neurosci. Методы 191, 110–118.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Сакс, О. (2004). В Реке Сознания . Нью-Йоркское обозрение книг. Доступно по адресу: http://www.readability.com/read?url=http://www.nybooks.com/articles/archives/2004/jan/15/in-the-river-of-сознания/?page= 2
Швейцер, П. (1994). Мгновенное сознание и буддийская эпистемология. Дж. Индийский Филос. 22, 81–91.
CrossRef Full Text
Шейнберг Д.Л. и Логотетис Н.К. (1997). Роль височных областей коры в организации восприятия. Проц. Натл. акад. науч. США 94, 3408–3413.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Симпсон, В.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Страуд, Дж. М. (1955). «Тонкая структура психологического времени», в Теория информации в психологии: проблемы и методы , изд. Х. Квастлер (Гленко, Иллинойс: Free Press), 174–207.
Саммерфилд, К., Джек, А.И., и Берджесс, А.П. (2002). Индуцированная гамма-активность связана с сознательным осознанием существительных, замаскированных узором. Междунар. Ж. Психофизиол. 44, 93–100.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Tallon-Baudry, C. (2009). Роль колебательной синхронности гамма-диапазона в зрительном познании человека. Фронт. Бионауч. 14, 321–332.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Таллон-Бодри, К., и Бертран, О.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Тонони Г., Сринивасан Р., Рассел Д. П. и Эдельман Г. М. (1998). Исследование нейронных коррелятов сознательного восприятия с помощью частотно-меченных нейромагнитных ответов. Проц. Натл. акад. науч. США 95, 3198–3203.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
VanRullen, R., and Koch, C. (2003). Восприятие дискретно или непрерывно? Тенденции Cogn. науч. 7, 207–213.
Опубликован Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Ван Руллен, Р., Редди, Л., и Кох, К. (2005). Управляемая вниманием дискретная выборка восприятия движения. Проц. Натл. акад. науч. США 102, 5291–5296.
Опубликован Аннотация | Опубликован полный текст | Полный текст CrossRef
Варела Ф.