Моногибридное скрещивание. Первый и второй законы Менделя – онлайн-тренажер для подготовки к ЕНТ, итоговой аттестации и ВОУД
Основные закономерности наследования были изучены Г. Менделем и изложены в его книге «Опыты над растительными гибридами» (1865).
Он проводил скрещивание растений гороха, при котором родительские формы анализировались по одной паре альтернативных признаков. Такое скрещивание называется моногибридным.
Если у родительских форм учитываются две пары альтернативных признаков, скрещивание называется дигибридным, более двух признаков – полигибридным.
Прежде чем проводить опыты, Г. Мендель получил чистые линии горохов с альтернативными признаками, т. е. гомозиготные доминантные (АА) – желтые и гомозиготные рецессивные (аа) зеленые особи, которые в дальнейшем скрещивались друг с другом.
При анализе результатов скрещивания оказалось, что все потомки в первом поколении одинаковы по фенотипу (проявляется доминантный признак желтой окраски – закон доминирования) и генотипу (гетерозиготны).
Первый закон Менделя – закон единообразия гибридов первого поколения: при скрещивании гомозиготных особей, отличающихся по одной паре альтернативных признаков, наблюдается единообразие гибридов первого поколения как по фенотипу, так и по генотипу.
По фенотипу особи АА и Аа неотличимы (желтые), поэтому наблюдается расщепление в отношении 3:1 (три части потомков с желтыми семенами и одна часть – с зелеными). По генотипу соотношение будет: 1АА (одна часть – желтые гомозиготы):2Аа (две части – желтые гетерозиготы):1аа (одна часть – зеленые гомозиготы).
Второй закон Менделя – закон расщепления: при скрещивании гибридов первого поколения наблюдается расщепление в соотношении 3:1 по фенотипу и 1:2:1 по генотипу.
Доминантный ген не всегда полностью подавляет действие рецессивного гена. В таком случае все гибриды первого поколения не воспроизводят признаки родителей – имеет место промежуточный характер наследования.
Промежуточный характер наследования
Во втором поколении доминантные гомо- и гетерозиготы будут отличаться фенотипически и расщепление по фенотипу и генотипу одинаково (1:2:1). Например, при скрещивании гомозиготных растений ночной красавицы с красными (АА) и белыми (аа) цветками первое поколение получается с розовыми цветками (промежуточное наследование).
Во втором поколении расщепление по фенотипу, как и по генотипу, будет: одна часть растений с красными цветками, две части – с розовыми и одна часть – с белыми.
Гипотеза чистоты гамет. У гетерозигот проявляется лишь доминантный признак, рецессивный ген у гетерозиготного организма не сливается с доминантным, не разбавляется, не изменяется, а остается в чистом аллельном состоянии. Аллельные гены расположены в одинаковых локусах гомологичных хромосом и в процессе мейоза попадают в разные гаметы. Следовательно, в гамете может присутствовать одновременно только один из аллельных генов, определяющий развитие одного из альтернативных признаков, и они являются «чистыми» по данному признаку.
Анализирующее скрещивание. При полном доминировании судить о генотипе организма по его фенотипу невозможно, поскольку и доминантная гомозигота (АА), и гетерозигота (Аа) обладают фенотипически доминантным признаком. Для того чтобы отличить доминантную гомозиготу от гетерозиготной, используют метод, называемый анализирующим скрещиванием, т. е. скрещивание исследуемого организма с организмом, гомозиготным по рецессивным аллелям. В этом случае рецессивная форма (аа) образует только один тип гамет с аллелем а, что позволяет проявиться любому из двух аллелей исследуемого признака уже в первом поколении.
Биология Решение генетических задач
|
Генетика является точной наукой. В ней есть законы и правила, которые можно проверить через задачи. Генетика изучает закономерности изменчивости и наследственности. Каждый биологический вид воспроизводит себе подобные организмы. |
|
При решении задач используются символы. Латинской буквой Р обозначаются родители, буквой F — гибридное потомство. G- гаметы. Заглавными буквами обозначаем доминантные гены, а прописными — рецессивные гены. Одинаковыми заглавными буквами обозначаем доминантные гомозиготы, а прописными — рецессивные гомозиготы. Х — знак скрещивания. Знаком «зеркало Венеры» ♀ обозначают женский пол. Знаком «щит и копье Марса» ♂ обозначают мужской пол. |
|
Существуют специальные правила оформления задач по генетике. |
|
Первым принято записывать генотип женской особи, а затем — мужской. |
|
Гены одной аллельной пары всегда пишутся рядом. |
|
При записи генотипа буквы, обозначающие признаки, всегда пишутся в алфавитном порядке, независимо от того, какой признак — доминантный или рецессивный — они обозначают. |
|
Под генотипом всегда пишут фенотип. |
|
У особей определяют и записывают типы гамет, а не их количество. |
|
При решении задач на дигибридное скрещивание для определения генотипов потомства рекомендуется пользоваться решёткой Пеннета. По вертикали записываются типы гаметы от материнской особи, а по горизонтали — отцовской. На пересечении столбца и горизонтальной линии записываются сочетание гамет, соответствующие генотипу образующейся дочерней особи |
|
Рассмотрим правила при решении задач по генетике. Правило первое. Если при скрещивании двух фенотипически одинаковых особей в их потомстве наблюдается расщепление признаков, то эти особи гетерозиготны. |
|
Правило второе. Если в результате скрещивания особей, отличающихся фенотипически по одной паре признаков, получается потомство, у которого наблюдается расщепление по этой же паре признаков, то одна из родительских особей гетерозиготна, а другая — гомозиготна по рецессивному признаку. |
|
Задачи по генетике имеют единые принципы решения. Но чтобы правильно решать задачи, необходимо определить их тип. Задачи могут быть на моногибридное, дигибридное скрещивание. Рассмотрим технологию решения задач на моногибридное скрещивание. |
|
Условие задачи. У арбуза зеленая окраска плодов доминирует над полосатой окраской. |
|
Для решения этой задачи запишем объект исследования и обозначение генов. Нам дан объект исследования — арбуз. Признак для исследования — окраска арбуза. Доминантный признак зеленой окраски обозначаем заглавной буквой А, а рецессивный признак полосатый окраски — прописной буквой а. Нам известны генотипы родительских форм. Необходимо определить окраску плодов арбузов, то есть фенотип первого поколения. |
|
Так как исследуется только один признак — окраска, то задача на моногибридное скрещивание. Записываем формулу скрещивания для родительских форм. Определяем тип гамет. Записываем генотипы первого поколения. Определяем фенотипы первого поколения. Записываем ответ. В результате скрещивания растений имеющих гетерозиготные и гомозиготные генотипы, в первом поколении вероятность появления зеленых и полосатых арбузов равна 50% на 50%. |
|
Решим еще один тип задач на моногибридное скрещивание. Условие задачи. У мышей длинные уши наследуются как доминантный признак. Короткие уши наследуются как рецессивный признак. Скрестили гомозиготного самца с длинными ушами с самкой с короткими ушами. Определить генотип самца, самки и фенотип первого поколения. |
|
Нам дан объект исследования — мыши. Признак для исследования — длина уха. Доминантный признак длинное ухо обозначаем заглавной буквой А, а рецессивный признак — короткое ухо — прописной буквой а. Определяем генотипы родительских форм. У гомозиготного самца с длинными ушами обозначаем генотип двумя заглавными буквами АА, а у самки генотип двумя маленькими буквами аа. Необходимо определить фенотип первого поколения. Записываем формулу скрещивания для родительских форм. Определяем тип гамет. Записываем генотипы первого поколения. Определяем фенотипы первого поколения. Записываем ответ. |
|
Рассмотрим решение задач на дигибридное скрещивание. Послушайте условие задачи. У фигурной тыквы белая окраска плодов А доминирует над желтой а, а дисковидная форма В — над шаровидной b. Ответьте на вопрос: как будет выглядеть F1и F2 от скрещивания гомозиготной белой шаровидной тыквы с гомозиготной желтой дисковидной? |
|
Рассмотрим решение задачи. Сначала определяем объект исследования — это тыква, исследуемые признаки: цвет и форма плодов. Записываем и обозначаем цвет плодов: ген А — белый, ген а — желтый; форма плодов: ген В — дисковидная ген b — шаровидная . Определяем генотипы родительских тыкв. По условиям задачи, тыквы гомозиготны, следовательно, содержат две одинаковые аллели каждого признака. |
|
Запишем схему скрещивания родительских растений и определим генотип и фенотип первого поколения. Как вы видите из схемы скрещивания, генотипы первого поколения тыкв все будут гетерозиготны по двум признакам. А по фенотипу все белые и дисковидные. |
|
Далее находим генотипы и фенотипы второго поколения. Для этого строим решетку Пеннета и вносим в нее все возможные типы гамет: по горизонтали вносим гаметы женской особи, по вертикали — мужской особи. На пересечении получаем возможные генотипы потомства второго поколения. |
|
Выпишем расщепление гибридов по фенотипу. Они будут следующие: 9 белых дисковидных*, 3 белых шаровидных**, 3 желтых дисковидных, 1 желтая шаровидная***. |
|
Запишем ответ: первое поколение — все белые дисковидные. Во втором поколении — 9 белых дисковидных, 3 белых шаровидных, 3 желтых дисковидных, 1 желтая шаровидная. |
 Однако нет идентичных особей, все потомки в большей или меньшей степени отличаются от своих родителей. Генетика дает возможность прогнозировать и анализировать передачу наследственных признаков. Для этого нужно уметь решать задачи по генетике.
Однако нет идентичных особей, все потомки в большей или меньшей степени отличаются от своих родителей. Генетика дает возможность прогнозировать и анализировать передачу наследственных признаков. Для этого нужно уметь решать задачи по генетике.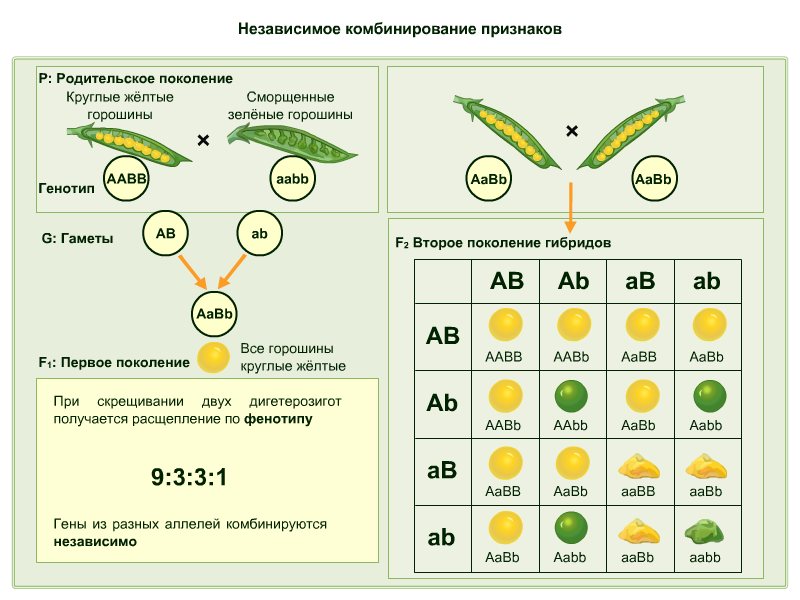 Предлагаем внимательно посмотреть на образец записи задачи.
Предлагаем внимательно посмотреть на образец записи задачи.





Соотношение фенотипов – определение и примеры
Соотношение фенотипов
сущ., множественное число: фенотипические соотношения
[fiːnə(ʊ)ˈtɪpɪk ˈreɪʃɪˌəʊ]
Определение: соотношение, основанное на фенотипах потомства и определяемое с использованием таких инструментов, как квадраты Пеннета.
Содержание
Определение соотношения фенотипов
Как определить соотношение фенотипов? Корреляция между количеством потомства, которое получит определенные признаки или комбинацию признаков, называется фенотипическое соотношение . Это соотношение обычно получается путем проведения тестового скрещивания, а затем с использованием информации из этого скрещивания, чтобы отметить, как часто будет проявляться признак или комбинация признаков на основе генотипа потомства.
См. примеры тестовых скрещиваний и фенотипов здесь: Что является ключом к распознаванию кодоминантности? J на нашем форуме. Открой для себя больше!
Что такое фенотипическое соотношение?
Фенотипическое соотношение равно количественному соотношению между фенотипами, показывающему, сколько раз частота одного фенотипа коррелирует с другим . Когда исследователь хочет получить экспрессию генов для поколений организма, он использует соотношение фенотипов, полученное в результате тестового скрещивания.
Тестовое скрещивание — это метод, используемый в генетике для изучения и получения возможных фенотипов и генотипов потомства организмов. Генотип организма — это его генетическая структура; он показывает аллели и гены, которые несет конкретный организм. Когда гены и аллели выражены в наблюдаемых признаках, это считается фенотипом. Фенотипы могут быть цветом глаз, ростом или даже текстурой волос. Генотипы можно использовать для определения фенотипов потомков организмов посредством тестового скрещивания и, в свою очередь, для получения соотношения фенотипов.
Фенотипы могут быть цветом глаз, ростом или даже текстурой волос. Генотипы можно использовать для определения фенотипов потомков организмов посредством тестового скрещивания и, в свою очередь, для получения соотношения фенотипов.
Например, если спариваться красный жук и синий жук, их потомство может быть красным, синим или фиолетовым (смесь обоих цветов). Нам нужно было бы найти соотношение фенотипов, чтобы определить, сколько раз наблюдается определенный фенотип по сравнению с другим фенотипом. Проще говоря, фенотипические отношения могут помочь нам определить вероятность того, что жук будет синим, красным или фиолетовым.
Биологическое определение:
Фенотипическое соотношение – это вероятность появления наблюдаемого признака при скрещивании. Соотношения фенотипов легче всего определить с помощью квадратов Пеннета или калькулятора соотношений фенотипов. Сравните: генотипическое соотношение
Важные генетические термины
Прежде чем узнать, как найти фенотипическое соотношение, необходимо знать некоторые генетические термины:
- Ген: Это то, что наследуется от родителя и передается потомству .

- Аллель: Разновидность гена, полученного от одного из двух родителей.
- Хромосома: Нитевидная структура, состоящая из нуклеиновых кислот и белков, несущих ген.
- Локус : Особое местонахождение гена в хромосоме.
- Гетерозигота : потомство, которое получает два разных аллеля, в частности, одного гена.
- Гомозиготный : Потомство, которое получает одинаковые аллели определенного гена от обоих родителей.
- Доминантный аллель: ген, который всегда будет проявляться как фенотип, даже когда он вступает в контакт с рецессивным.
- Рецессивный аллель : ген, который будет проявляться как фенотип только при контакте с другим рецессивным аллелем.
- Моногибрид : Это происходит, когда два родителя скрещиваются и производят потомство только с одним фенотипом.
- Дигибрид : Когда два родителя скрещиваются и получают потомство с комбинациями фенотипов родителей.
- Тригибрид: Когда два родителя скрещиваются и дают потомство, которое выражает более широкий спектр фенотипов, чем дигибрид.
- Punnett Square : Квадратная диаграмма используется в качестве инструмента для определения генотипа потомства при скрещивании конкретных родителей.


Как найти соотношение фенотипов
Как рассчитать соотношение фенотипов. Чтобы найти фенотипическое соотношение, мы смотрим на аллели родительских организмов и оцениваем, как часто эти гены будут проявляться у потомства. В большинстве случаев мы знаем, что будут выражать аллели и как они будут выглядеть. Соотношения фенотипов легче всего определить с помощью квадратов Пеннета или калькулятора соотношений фенотипов.
Какова формула соотношения фенотипов? Чтобы использовать формулу соотношения фенотипов, нужно сначала использовать частотную диаграмму – это можно сделать, если она не существовала до предоставления информации. Определите каждую желаемую черту и организуйте их в столбцы. Затем подсчитайте количество особей с определенными признаками, чтобы организм учитывался только один раз. Частоты ранжируются от наименьшей к наибольшей. Затем каждая частота будет разделена на наименьшую возможную частоту, а ответ будет отмечен в другом столбце таблицы. Эти ответы будут округлены и использованы в качестве соотношения фенотипов. Например, в приведенной ниже таблице 1 окончательное соотношение фенотипов будет равно 9.0003 9:3:1 , где 9 представляет черные волосы, 3 — каштановые волосы, а 1 — рыжие волосы.
Определите каждую желаемую черту и организуйте их в столбцы. Затем подсчитайте количество особей с определенными признаками, чтобы организм учитывался только один раз. Частоты ранжируются от наименьшей к наибольшей. Затем каждая частота будет разделена на наименьшую возможную частоту, а ответ будет отмечен в другом столбце таблицы. Эти ответы будут округлены и использованы в качестве соотношения фенотипов. Например, в приведенной ниже таблице 1 окончательное соотношение фенотипов будет равно 9.0003 9:3:1 , где 9 представляет черные волосы, 3 — каштановые волосы, а 1 — рыжие волосы.
Когда выражены оба пенотипа родителей, это называется кодоминантностью. Узнайте больше об этом здесь: Что является ключом к признанию кодоминантности? Дж на нашем форуме прямо сейчас!
Расчет соотношения фенотипов для типов скрещивания
Можно использовать либо калькулятор соотношения фенотипов, разработанный для конкретных скрещиваний, либо квадрат Пеннета. Во многих случаях расчеты могут быть затруднены, поскольку фенотипы видны при объединении множества аллелей. Однако следующие примеры будут выполнены с использованием одного аллеля, который даст только один признак.
Во многих случаях расчеты могут быть затруднены, поскольку фенотипы видны при объединении множества аллелей. Однако следующие примеры будут выполнены с использованием одного аллеля, который даст только один признак.
При использовании этих методов расчета мы можем получить результаты для фенотипов, которые появятся в первое дочернее поколение (F1) скрещивания и последующие поколения. Мы даже можем определить различные эффекты, которые могут возникнуть в этих последующих поколениях. Ранние заводчики лошадей и собак научились выращивать животных с разными чертами, даже не зная тех подробностей, которые мы знаем сегодня о генетике. Этот тип селекционного разведения привел к появлению огромного количества пород животных, которые мы имеем сегодня в нашем мире.
Некоторые фенотипические отношения могут быть простыми.
Что такое фенотипическое соотношение 1:1? Соотношение фенотипов 1:1 возникает, когда есть только два возможных фенотипа в качестве исходов при скрещивании организмов, и они оба имеют шанс появления 50/50. Как выглядит фенотипическое соотношение 3:1? Это происходит, когда каждый из двух гетерозиготных родителей передает по одному аллелю своему потомству, создавая два возможных фенотипа, даже если может быть несколько генотипов. Важно отметить, что генотипические и фенотипические соотношения не всегда будут одинаковыми. На рисунках 1 и 2 показаны примеры соотношения фенотипов 1:1 и 3:1 на площадях Пеннета соответственно.
Как выглядит фенотипическое соотношение 3:1? Это происходит, когда каждый из двух гетерозиготных родителей передает по одному аллелю своему потомству, создавая два возможных фенотипа, даже если может быть несколько генотипов. Важно отметить, что генотипические и фенотипические соотношения не всегда будут одинаковыми. На рисунках 1 и 2 показаны примеры соотношения фенотипов 1:1 и 3:1 на площадях Пеннета соответственно.
Рисунок 2. Пример соотношения фенотипов 3:1 на площади Пеннетт. Источник: Мария Виктория Гонзага из Biology Online.
В следующем разделе будут рассмотрены различные примеры соотношения фенотипов, которые являются более сложными. Это будет включать моногибридные, дигибридные и тригибридные скрещивания.
Соотношение фенотипов при моногибридном скрещивании
Моногибридное скрещивание происходит между двумя гомозиготными родителями, поэтому у их потомства образуется только один фенотип. Это также может произойти, когда генотипы обоих родителей являются полностью доминантными или полностью рецессивными, что при определенных генетических признаках приводит к противоположному фенотипу. Это легко определить с помощью квадрата Пеннета.
Это также может произойти, когда генотипы обоих родителей являются полностью доминантными или полностью рецессивными, что при определенных генетических признаках приводит к противоположному фенотипу. Это легко определить с помощью квадрата Пеннета.
На рисунке 3 ниже мы видим моногибридное скрещивание. В этом случае AA, родитель мужского пола обладает фенотипом высокого дерева, а аа, родитель женского пола обладает фенотипом низкорослого дерева. A является доминантным признаком, означающим, что когда он присутствует, организм всегда будет демонстрировать свой фенотип независимо от того, присутствует рецессивный ген или нет. Ген a является рецессивным и будет виден только в паре с другими аллель .
Когда родители размножаются, каждый из них передает своему потомству один из своих аллелей для формирования его хромосомы. Поскольку потомство должно получить по одному аллелю от каждого родителя и поскольку оба родителя гомозиготны, каждое потомство будет гетерозиготным. Это делает каждое потомство, которое они производят, становится высоким деревом, поскольку их генотип будет Aa . Таким образом, поскольку все 4 потомства имеют одинаковый фенотип, нет необходимости измерять соотношение фенотипов. Это связано с тем, что, несмотря на наличие двух возможных результатов (высокое или низкое дерево), только один из них является наблюдаемым признаком, поэтому вычисление фенотипического соотношения было бы излишним. Если необходимо показать фенотипическое соотношение, оно будет записано как 4:0 .
Поскольку потомство должно получить по одному аллелю от каждого родителя и поскольку оба родителя гомозиготны, каждое потомство будет гетерозиготным. Это делает каждое потомство, которое они производят, становится высоким деревом, поскольку их генотип будет Aa . Таким образом, поскольку все 4 потомства имеют одинаковый фенотип, нет необходимости измерять соотношение фенотипов. Это связано с тем, что, несмотря на наличие двух возможных результатов (высокое или низкое дерево), только один из них является наблюдаемым признаком, поэтому вычисление фенотипического соотношения было бы излишним. Если необходимо показать фенотипическое соотношение, оно будет записано как 4:0 .
Соотношение фенотипов дигибридного скрещивания
Дигибридное скрещивание вступает в игру, когда участвуют два фенотипа. Однако есть причина, по которой заводчики обычно не сосредотачиваются на использовании только одного фенотипа. Если они это сделают, им никогда не удастся изучить другие возможности и разработать гораздо более уникальные и многообещающие функции. Зачем разводить более крупных свиней ради большего количества мяса, если они просто унаследовали дефекты мозга от обоих родителей. Следовательно, генетики продолжают искать и продвигать полезные породы, а затем предотвращать разведение менее благоприятных. Они могут использовать калькулятор дигибридного скрещивания, чтобы получить соотношение фенотипов.
Если они это сделают, им никогда не удастся изучить другие возможности и разработать гораздо более уникальные и многообещающие функции. Зачем разводить более крупных свиней ради большего количества мяса, если они просто унаследовали дефекты мозга от обоих родителей. Следовательно, генетики продолжают искать и продвигать полезные породы, а затем предотвращать разведение менее благоприятных. Они могут использовать калькулятор дигибридного скрещивания, чтобы получить соотношение фенотипов.
На рис. 4 выше мы видим дигибридное скрещивание двух желтых горошин в поколении F1. Предыдущее или родительское поколение этого скрещивания состояло из двух гомозиготных родителей, одного доминантного ( RR YY ), одного рецессивного ( rr yy ). Доминантный RR выставлял круглый горошек и YY желтого цвета. Рецессивный рр выставлены морщинистым горошком, а yy, зеленого цвета.
В родительском скрещивании желтый округлый горох ( RRYY ) скрещен с зеленым морщинистым горохом ( rryy ). Это приводит к тому, что все их потомки становятся круглыми и желтыми, — моногибридным скрещиванием . Однако они несут гены, дающие желтые, зеленые, круглые или морщинистые аллели ( RrYy ), и поэтому являются гетерозиготными.
Когда два из этих потомков RrYy скрещиваются, они производят различные виды фенотипов. Они несут аллели как круглой ( R ), так и морщинистой ( r ) и аллели желтой ( Y ) и зеленой ( y) ).
Использование квадрата Пеннета для определения фенотипов потомства является простым и дает надежную визуализацию. Найти соотношение фенотипов легко с помощью калькулятора дигибридных квадратов Пеннета. На рис. 5 ниже показано, как легко можно подсчитать частоту генотипов и Соотношение 9:3:3:1 получается для этого кросса. Это может быть использовано для всех типов фенотипов.
5 ниже показано, как легко можно подсчитать частоту генотипов и Соотношение 9:3:3:1 получается для этого кросса. Это может быть использовано для всех типов фенотипов.
Соотношение фенотипов тригибридного скрещивания
Если добавляется еще один аллель, генетическая экспрессия и возможные фенотипические результаты расширяются еще больше. Калькулятор тригибридного скрещивания будет использоваться для расчета соотношения фенотипов для таких пород, как эта. Тригибридные кросс-отношения могут быть очень длинными из-за множества возможных результатов, которые можно получить от них.
Возьмем, к примеру, следующий пример:
Известно, что люди имеют несколько типов волос. В конкретном эксперименте генетики хотят увидеть, что произойдет, если они скрещивают людей по длине, цвету и текстуре волос. Доминантный аллель А даст длинные волосы, а рецессивный аллель а даст короткие волосы. Для черных волос ген представлен как доминантный B , а для каштановых волос — как рецессивный b . Наконец, прямые волосы будут представлены D и являются доминантным аллелем, тогда как кудрявые волосы представляют собой рецессивный аллель, представленный d .
Для черных волос ген представлен как доминантный B , а для каштановых волос — как рецессивный b . Наконец, прямые волосы будут представлены D и являются доминантным аллелем, тогда как кудрявые волосы представляют собой рецессивный аллель, представленный d .
Ученые начинают с моногибридного скрещивания фенотипа длины волос. Они скрещивают один ген, используя двух гетерозиготных родителей. Это можно увидеть на рисунке 6 ниже. Как было показано ранее в этой статье, это оставляет нам фенотипическое соотношение 3:1, производя потомство как с длинной, так и с короткой шерстью. Несмотря на то, что некоторые из наблюдаемых признаков потомства — длинные волосы, они несут рецессивный ген короткой шерсти. Вероятность потомства с длинными волосами намного выше, чем с короткими.
Рисунок 6: Моногибридное скрещивание по типам волос. Предоставлено: Университет штата Орегон.
Проведено второе скрещивание, теперь оно включает ген цвета волос. Это дигибридное скрещивание даст более двух фенотипических результатов, поскольку теперь два гена уступают место множественным фенотипическим результатам. Это приводит фенотипическое соотношение к 9:3:3:1 с возможными вариантами: длинные черные волосы, длинные каштановые волосы, короткие черные волосы и короткие каштановые волосы соответственно. Мы видим, что по мере того, как в разведение добавляется больше генов, мы получаем более крупные и сложные фенотипы.
Рис. 7. Показано дигибридное скрещивание по типам волос, включая длину и цвет волос. Предоставлено: Университет штата Орегон.
Наконец, добавляется третий ген, отвечающий за текстуру волос. Это тригибридное кросс-отношение можно получить с помощью калькулятора площади Пеннета, как и в случае моногибридных и дигибридных кроссов. В этом фенотипическом соотношении всего 8 возможных видимых признаков. Все они объединяются уникальным образом и производят особое потомство.
Наиболее популярным фенотипом будет сочетание всех доминантных аллелей – потомство человека черного, длинного, прямоволосого. Это связано с тем, что при объединении аллелей доминантный аллель всегда будет иметь приоритет, если он присутствует. Другими наблюдаемыми фенотипами будут:
- Длинные, прямые и каштановые волосы
- Длинные, вьющиеся и черные волосы
- длинные, вьющиеся и каштановые волосы
- Короткие, прямые и черные волосы
- Короткие, прямые и каштановые волосы
- Короткие, вьющиеся и черные волосы
- Короткие, вьющиеся и каштановые волосы
Как и ожидалось, наименее распространенным является тот фенотип, который включает все три рецессивных гена – aabbdd . Вероятность того, что это произойдет только один раз из шестидесяти четырех (64) возможных пересечений.
Рисунок 8: Тригибридное скрещивание типов волос с многочисленными фенотипическими результатами – цветом, длиной и текстурой волос. Предоставлено: Университет штата Орегон.
Предоставлено: Университет штата Орегон.Эти диаграммы показаны в качестве примера соотношения фенотипов с участием одного, двух и трех генов соответственно. Однако в действительности наследование признаков человеческого волоса является более сложным, поскольку фенотип определяется взаимодействием многих генов (аллелей) во многих локусах.
Посмотрите видео ниже, чтобы узнать больше о соотношениях фенотипов и генотипов.
Возник вопрос по теме? Спросите наше сообщество! Присоединяйтесь к нам здесь: Что является ключом к признанию кодоминантности? Будь частью нас!
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы уже узнали о фенотипических соотношениях.
Викторина
Выберите лучший ответ.
Фенотипическое соотношение
Генотипическое соотношение
Частота аллелей
2. Вариант генаХромосома
Локус
Аллель
3. Сколько признаков, вероятно, проявится при соотношении фенотипов 3:1?Один
Два
Три
4. Что из следующего приведет к фенотипическому соотношению 3:1?Аа х Аа
Аа х Аа
Аа х Аа
5. Аллель, который экспрессируется и маскирует эффект другого аллеля 9.0008Доминантный аллель
Рецессивный аллель
Множественный аллель
Отправьте результаты (необязательно)
Ваше имя
На электронную почту
NextРазличие генотипа и фенотипа: от менделевской генетики к биологии XXI века
Alberch P (1991) От генов к фенотипу: динамические системы и эволюционируемость. Genetica 84:5–11
Genetica 84:5–11
Статья КАС пабмед Google Scholar
Avery OT, MacLeod CM, McCarty M (1944) Изучение химической природы вещества, вызывающего трансформацию пневмококкового типа. J Exp Med 79(2):137–158
Статья КАС пабмед ПабМед Центральный Google Scholar
Авиталь Э., Яблонка Э. (2000) Традиции животных: наследование поведения в эволюции. Издательство Кембриджского университета, Кембридж
Книга Google Scholar
Бейтсон В. (1902) Принципы наследственности Менделя. Защита. Издательство Кембриджского университета, Кембридж
Книга Google Scholar
Bernard C (1865) Introduction à l’Etude de la Médecine Expérimental. Байер, Париж
Google Scholar
Берталанфи Л. В. (1952) Проблемы жизни оценка современной биологической и научной мысли. Harper & Brothers, Нью-Йорк
В. (1952) Проблемы жизни оценка современной биологической и научной мысли. Harper & Brothers, Нью-Йорк
Google Scholar
Буассо А. (В печати) La Causalité Du Développement Selon C. H. Waddington
Бондурянский Р. (2012) Снова переосмысление наследственности. Тренды Ecol Evol 27(6):330–336
Артикул КАС пабмед Google Scholar
Бондурянский Р., Дэй Т. (2018) Расширенная наследственность: новое понимание наследственности и эволюции. Издательство Принстонского университета, Принстон
Книга Google Scholar
Boveri T (1904) Ergebnisse Über Die Konstitution Der Chromatischen Substanz Des Zellkerns. Йена: Г. Фишер
Боулер П. (2001) Менделевская революция: появление наследственных концепций в современной науке и обществе. Международная издательская группа Continuum
Braun E (2015) Непредвиденная проблема: от генотипа к фенотипу в клеточных популяциях. Rep Prog Phys 78(3):1–51
Rep Prog Phys 78(3):1–51
Статья КАС Google Scholar
Шампанское F (2008) Эпигенетические механизмы и трансгенерационные эффекты материнской заботы. Front Neuroendocrinol 29(3):386–397
Статья КАС пабмед ПабМед Центральный Google Scholar
Корниш-Боуден А., Карденас М.Л. (2020) Противопоставление теорий жизни: исторический контекст, современные теории. В поисках идеальной теории. Биосистемы 1:104063
Артикул Google Scholar
Crick FH (1958) О синтезе белка. В: Сандерс Ф.К. (ред.) Симпозиумы Общества экспериментальной биологии, номер XII: Биологическая репликация макромолекул. Издательство Кембриджского университета, Кембридж. стр. 138–163
Danchin E et al (2019)Эпигенетически облегчённая мутационная ассимиляция: эпигенетика как центр инклюзивного эволюционного синтеза. Биол. Ред. 94 (259–282): 259. https://doi.org/10.1111/brv.12453
Биол. Ред. 94 (259–282): 259. https://doi.org/10.1111/brv.12453
Статья Google Scholar
Дарвин К. (1859 г.) О происхождении видов путем естественного отбора. Изменчивость животных и растений при одомашнивании. Джон Мюррей, Лондон
Google Scholar
Дарвин К. (1868) Изменчивость животных и растений при одомашнивании, том 2. Джон Мюррей, Лондон
Google Scholar
Дарвин К. (1871) Происхождение человека и отбор в отношении пола. 1-е изд. Том. 1–2. John Murray, London
De Vienne D (этот выпуск) Что такое фенотип? История и новости концепции. Genetica
De Vries H (1889) [1910] Внутриклеточный пангенезис. Издательство Open Court Publishing Co., Чикаго
Danchin E, Pocheville A, Rey O, Pujol B, Blanchet S (2011)Эпигенетически облегчаемая мутационная ассимиляция: эпигенетика как центр инклюзивного эволюционного синтеза. Biol Rev 94:259–282
Biol Rev 94:259–282
Статья Google Scholar
Danchin E, Pocheville A (2014) Наследование — это место, где физиология встречается с эволюцией. J Physiol 592 (11): 2307–2317. https://doi.org/10.1113/jphysiol.2014.272096
Статья КАС пабмед ПабМед Центральный Google Scholar
Danchin É, Wagner RH (2010) Инклюзивная наследуемость: объединение генетической и негенетической информации для изучения поведения и культуры животных. Oikos 119(2):210–218
Статья Google Scholar
Докинз Р. (2006) Эгоистичный ген, 3-е изд. Издательство Оксфордского университета, Оксфорд
Google Scholar
Добжанский Т. (1937) Генетика и происхождение видов. Издательство Колумбийского университета, Нью-Йорк
Google Scholar
Falk R (1984) Ген в поисках идентичности. Гум Жене 68 (3): 195–204. https://doi.org/10.1007/BF00418388
Гум Жене 68 (3): 195–204. https://doi.org/10.1007/BF00418388
Франклин Р.Э., Гослинг Р.Г. (1953) Молекулярная конфигурация тимононуклеата натрия. Природа 171 (4356): 740–741. https://doi.org/10.1038/171740a0
Статья КАС пабмед Google Scholar
Friedberg EC, Walker GC, Siede W, Wood RD (Eds.) (2005) Репарация ДНК и мутагенез. Американское общество микробиологии Press.
Forterre P, Filee J, Myllykallio H (2004) Происхождение и эволюция ДНК и механизмов репликации ДНК. В: Рибас де Пуплана Л. (ред.) Генетический код и происхождение жизни. Landes Bioscience, стр. 145–168
Фокс Келлер Э. (1983 [2003]) Чувство организма, издание, посвященное 10-летию: жизнь и работа Барбары МакКлинток. Macmillan
Fox Keller E (2000) Век ген. Издательство Гарвардского университета, Кембридж
Книга Google Scholar
Галеф Б. Г. младший, Лаланд К.Н. (2005) Социальное обучение у животных: эмпирические исследования и теоретические модели. Bioscience 55(6):489–499
Г. младший, Лаланд К.Н. (2005) Социальное обучение у животных: эмпирические исследования и теоретические модели. Bioscience 55(6):489–499
Гальтон Ф. (1865 г.) Наследственный талант и характер. Журнал Macmillan’s 12:157–166
Google Scholar
Гальтон Ф. (1872 г.) О кровном родстве. Proc Royal Soc Lond 20: 394–402
Гальтон Ф. (1876 г.) Теория наследственности. J Anthropol Inst GB Irel 5:329–348
Google Scholar
Гальтон Ф. (1887 г.) О кровном родстве. Proc R Soc Lond 20:394–402
Google Scholar
Гальтон Ф. (1886) Регрессия к посредственности в наследственном росте. J Anthropol Inst GB Irel 15:246–263
Google Scholar
Gayon J (1992) Darwin et l’après-Darwin: une histoire de l’hypothèse de sélection dans la théorie de l’évolution. Киме, Париж
Киме, Париж
Google Scholar
Gayon J (2004) Генетическая проблема на бис дисциплины? Медицина/науки 20(2):248–253
Статья Google Scholar
Gayon J (2000) От измерения к организации: философская схема истории концепции наследственности. В: Beurton P, Falk R, Rheinberger HJ (eds) Концепция гена в развитии и эволюции: исторические и эпистемологические перспективы. Издательство Кембриджского университета, Кембридж, стр. 69.–90
Глава Google Scholar
Gayon J, Petit V (2018) La connaissance de la vie aujourd’hui. Издания ISTE
Гилберт С.Ф. (2003) Биология развития, 7-е изд. Sinauer Associates, Сандерленд, Массачусетс
Google Scholar
Гилберт С.Ф., Саркар С. (2000) Охватывая сложность: организм для 21 века. Дев Дин 219(1):1–9
Артикул КАС пабмед Google Scholar
Гилберт С.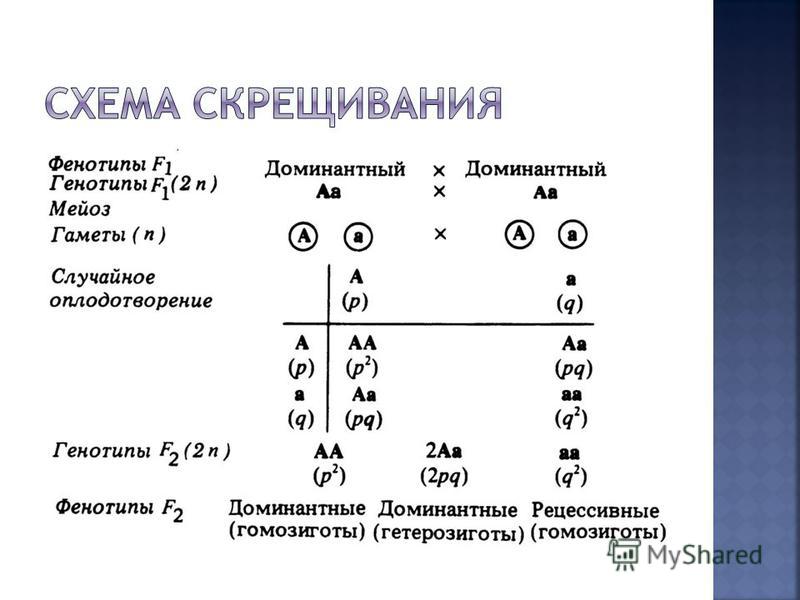 , Сапп Дж., Таубер А. (2012) Симбиотический взгляд на жизнь: мы никогда не были личностями. Q Rev Biol 87:325–341
, Сапп Дж., Таубер А. (2012) Симбиотический взгляд на жизнь: мы никогда не были личностями. Q Rev Biol 87:325–341
Статья пабмед Google Scholar
Griesemer J (2000) Развитие, культура и единицы наследования. Философия науки 67:S348–S368
Статья Google Scholar
Griffiths PE, Stotz K (2013) Генетика и философия: введение. Издательство Кембриджского университета, Кембридж
Книга Google Scholar
Haig D (2012) Комментарий: эпидемиология эпигенетики. Int J Epidemiol 41(1):13–16
Статья пабмед Google Scholar
Хемс Т. (2014) Случайность в биологии. Математическая структура Компьютерные науки 24 (3). https://doi.org/10.1017/S096012951200076X
Херд Э., Мартиенссен Р.А. (2014) Трансгенерационное эпигенетическое наследование: мифы и механизмы. Ячейка 157(1):95–109
Ячейка 157(1):95–109
Артикул КАС пабмед ПабМед Центральный Google Scholar
Хофмайстер Б.Т., Ли К., Рор Н.А., Холл Д.В., Шмитц Р.Дж. (2017)Стабильное наследование метилирования ДНК позволяет создавать карты эпигенотипов и изучать модели наследования эпиаллелей в отсутствие генетической изменчивости. Геном Биол 18(1):155. https://doi.org/10.1186/s13059-017-1288-х
Артикул КАС пабмед ПабМед Центральный Google Scholar
Корпус D (1988 г.) Interactors против транспортных средств. В: Плоткин Х.К. (ред.) Роль поведения в эволюции. MIT Press, Кембридж, Массачусетс, стр. 19–50
Google Scholar
Яблонка Э., Лэмб М. (1995) Эпигенетическое наследование и эволюция: ламарковское измерение. Издательство Оксфордского университета, Оксфорд
Google Scholar
Яблонка Э. , Лэмб М. (2005) Эволюция в четырех измерениях: генетические, эпигенетические, поведенческие и символические вариации в истории жизни. MIT Press, Кембридж
, Лэмб М. (2005) Эволюция в четырех измерениях: генетические, эпигенетические, поведенческие и символические вариации в истории жизни. MIT Press, Кембридж
Google Scholar
Яблонка Э., Раз Г. (2009) Трансгенерационное эпигенетическое наследование: распространенность, механизмы и последствия для изучения наследственности и эволюции. Quart Rev Biol 84(2):131–176
Джейкоб Ф. (1970) Логика жизни, история наследственности. Галлимар, Париж
Google Scholar
Джейкоб Ф., Моно Дж. (1961) Генетические регуляторные механизмы синтеза белков. J Mol Biol 3:318–356
Статья КАС пабмед Google Scholar
Джагер Дж., Монк Н. (2014) Биоаттракторы: теория динамических систем и эволюция регуляторных процессов. Дж Физиол 592(11):2267–2281
Статья КАС пабмед ПабМед Центральный Google Scholar
Johannsen WL (1909) Elemente der Exakten Erblichkeitslehre. Йена: Густав Фишер, 170
Йена: Густав Фишер, 170
Йоханнсен В.Л. (1911) Генотипическая концепция наследственности. Am Nat 45(531):129–159
Статья Google Scholar
Джоли Д., Грунау С. (редакторы) (2018) Перспективная эпигенетика, экология и эволюция. Une Prospective de l’Institut Ecologie et Environnement, 8, CNRS
Кант I (1790/1987) Критика суждения. Hackett Publishing, Индианаполис
Кауфман С. (2000) Исследования. Издательство Оксфордского университета, Оксфорд
Google Scholar
Китано Х (2002) Биология вычислительных систем. Природа 420:206–210
Статья КАС пабмед Google Scholar
Кун Т.С. (1970) Структура научных революций. University of Chicago Press, Чикаго
Google Scholar
Купец Дж. Дж. , Сониго П. (2000) Ni Dieu ni gène. Pour une autre théorie de l’heredité. Seuil, Paris
, Сониго П. (2000) Ni Dieu ni gène. Pour une autre théorie de l’heredité. Seuil, Paris
Laland KN, Odling-Smee JF, Myles S (2010) Как культура сформировала человеческий геном: объединение генетики и гуманитарных наук. Nat Rev Genet 11(2):137–148
Статья КАС пабмед Google Scholar
Ламарк Ж-Б де (1809) Философия зоологии или изложение соображений относительно родственников à l’histoire naturelle des animaux. Том. Премьера тома. 2 тт. Dentu, Paris
Lamm E, Jablonka E (2008) Воспитание природы: наследственная пластичность в эволюции. Philos Psychol 21(3):305–319
Статья Google Scholar
Леманн Л. (2008)Адаптивная динамика признаков построения ниши в пространственно разделенных популяциях: эволюционирующие посмертные расширенные фенотипы. Эволюция 62(3):549–566
Артикул пабмед Google Scholar
Ленай С. (1990) La découverte des lois de l’héredité (1862–1900), une anthologie. Presses Pocket, Париж
(1990) La découverte des lois de l’héredité (1862–1900), une anthologie. Presses Pocket, Париж
Google Scholar
Левонтин RC (1974) Дисперсионный анализ и анализ причин. Am J Hum Genet 26: 400–411. https://doi.org/10.1093/ije/dyl062
Статья КАС пабмед ПабМед Центральный Google Scholar
Левонтин Р.С. (1992) Генотип и фенотип. Ключевые слова эволюционной биологии, 4-е изд. Издательство Гарвардского университета, Гарвард, стр. 137–144
Google Scholar
Лонго Г., Микель П.А., Зонненшайн С., Сото А. (2012) Является ли информация подходящей наблюдаемой для биологической организации? Prog Biophys Mol Biol 109(3):108–114
Артикул КАС пабмед ПабМед Центральный Google Scholar
Лопес-Бельтран С. (1994) Ковка наследственности: от метафоры к причине, история овеществления. Исследования по истории и философии науки Часть а 25(2):221–235
Исследования по истории и философии науки Часть а 25(2):221–235
Статья Google Scholar
Махар Б. (2008) Случай отсутствия наследуемости. Природа 456(7218):18–21
Артикул КАС Google Scholar
Махер Б. (2008) Персональные геномы: случай отсутствия наследственности. Природа 456(7218):18–21
Артикул КАС пабмед Google Scholar
Матурана Х., Варела Ф. (1980) Автопоэзис и познание: реализация живого. Reidel, Бостон
Книга Google Scholar
Maynard SJ (1958) Теория эволюции, 3-е издание (1993). Издательство Кембриджского университета, Кембридж
Майр Э. (1961) Причина и следствие в биологии. Наука 134:1501–1506
Артикул КАС пабмед Google Scholar
Mayr E (1998) Пролог: некоторые мысли об истории эволюционного синтеза
Mayr E, Provine WB (Eds) The Evolutionary Synthesis: Perspectives on the Unification of Biology. Издательство Гарвардского университета, Кембридж. стр. 1–48
Издательство Гарвардского университета, Кембридж. стр. 1–48
Мендель Г. (1901 г.) Эксперименты по гибридизации растений (1865 г.) Прочитано 8 февраля и 8 марта 1865 г., собрания Брюннского общества естественной истории, перевод Уильяма Бейтсона. http://www.esp.org/foundations/genetics/classical/gm-65-a.pdf. (Мендель, Грегор. 1866. Versuche über Plflanzenhybriden. Verhandlungen des naturforschenden Vereines in Brünn, Bd. IV für das Jahr 1865, Abhandlungen, 3–47.)
Мерлин Ф. (2010) Эволюционная случайная мутация: защита консенсусной точки зрения современного синтеза. Филос Теория Биол 2 (201306): 1–22
Google Scholar
Мерлин Ф., Риболи-Саско Л. (2017) Картирование биологической передачи: эмпирический, динамический и эволюционный подход. Acta Biotheor 65(2):97–115
Статья пабмед Google Scholar
Монтевиль М., Моссио М. (2015) Биологическая организация как закрытие ограничений. J Theor Biol 372:179–191
(2015) Биологическая организация как закрытие ограничений. J Theor Biol 372:179–191
Статья пабмед Google Scholar
Монтевиль М., Моссио М., Пошевиль А., Лонго Г. (2016) Теоретические принципы биологии: вариации. Prog Biophys Mol Biol 122(1):36–50
Morange M (2003) Histoire de la biologie moléculaire. La Découverte, Париж
Забронировать Google Scholar
Морено А., Моссио М. (2015) Биологическая автономия. Философско-теоретическое исследование. Спрингер, Дордрехт
Google Scholar
Моссио М., Монтевиль М., Лонго Г. (2016) Теоретические принципы биологии: организация. Prog Biophys Mol Biol 122(1):24–35
Статья пабмед Google Scholar
Моссио М., Понтаротти Г. (2019) Сохранение функций между поколениями: наследственность в свете биологической организации. Br J Philos Sci. https://doi.org/10.1093/bjps/axz031
Br J Philos Sci. https://doi.org/10.1093/bjps/axz031
Артикул Google Scholar
Muller HJ (1922) Изменчивость вследствие изменения отдельного гена. Am Nat 56(642):32–50
Статья Google Scholar
Мюллер-Вилле С., Райнбергер Х.-Дж. (2007) Наследственность – формирование эпистемологического пространства. В: Müller-Wille S, Rheinberger HJ (ред.) Произведенная наследственность: на перекрестке биологии, политики и культуры, 1500–1870 гг. MIT Press, Кембридж, стр. 3–34
Глава Google Scholar
Noble D (2008) Клод Бернар, первый системный биолог и будущее физиологии. Exp Physiol 93(1):16–26
Статья пабмед Google Scholar
Одлинг-Сми Дж. (2010) Наследование ниши. В: Пиглиуччи М., Мюллер Г.Б. (ред.) Эволюция – расширенный синтез. MIT Press, Кембридж, стр. 175–208
MIT Press, Кембридж, стр. 175–208
Глава Google Scholar
Оргогозо В., Моризо Б., Мартин А. (2015) Дифференциальный взгляд на отношения генотип-фенотип. Фронт Жене 19(6):179. https://doi.org/10.3389/fgene.2015.00179
Статья КАС Google Scholar
Ояма С. (1985) Онтогенез информации: системы развития и эволюция. Издательство Университета Дьюка, Нью-Йорк
Google Scholar
Nature Reviews (2021), серия по восстановлению повреждений ДНК. www.nature.com/collections/hwnqqcstyj/
Перл Дж. (2009) Причинность: модели, рассуждения и выводы, 2-е изд. Издательство Кембриджского университета, Нью-Йорк
Книга Google Scholar
Питер Т., Левонтин Р. (2021) Различие генотипа и фенотипа, Стэнфордская философская энциклопедия (лето 2021 г. ), Zalta EN (ред.) https://plato.stanford.edu/archives/sum2021/entries /генотип-фенотип/
), Zalta EN (ред.) https://plato.stanford.edu/archives/sum2021/entries /генотип-фенотип/
Piaget J (1967) Biologie et Connaissance. Галлимар, Париж
Google Scholar
Пиглиуччи М. (2010) Сопоставление генотипа и фенотипа и конец метафоры «гены как план». Philos Trans R Soc Lond B Biol Sci 365 (1540): 557–566. https://doi.org/10.1098/rstb.2009.0241 PMID: 20083632; PMCID: PMC2817137
Pigliucci M, Murren CJ, Schlichting CD (2006) Фенотипическая пластичность и эволюция путем генетической ассимиляции. J Эксперт Биол 209(12):2362–2367
Артикул пабмед Google Scholar
Pocheville A (2010) Что такое строительство ниши (не). В: La Niche Écologique: концепции, модели, приложения. (Докторская диссертация), Арно Пошевиль, стр. 39–124. Ecole Normale Supérieure Paris, Paris
Pocheville A (2018) Биологическая информация как выбор и конструкция. Под редакцией Венди Паркер. Философия науки, Материалы двухгодичного собрания Ассоциации философии науки 2016 г., 85 (5).
Под редакцией Венди Паркер. Философия науки, Материалы двухгодичного собрания Ассоциации философии науки 2016 г., 85 (5).
Pocheville A (2019) Дарвиновская мечта: о времени, уровнях и процессах эволюции. В: Уллер Т., Лаланд К.Н. (ред.) Эволюционная причинность. Биологические и философские размышления, Венская серия теоретической биологии. MIT Press, Бостон
Google Scholar
Пошевиль А., Данчин Э. (2017) Генетическая ассимиляция и парадокс слепой изменчивости. В: Уолш Д.М., Хунеман П. (ред.) Вызов современному синтезу. Издательство Оксфордского университета, Оксфорд
Google Scholar
Понтаротти Г. (2016) Расширенное наследование как реконструкция расширенной организации: парадигматический случай симбиоза. Lato Sensu 3(1):93–102
Статья Google Scholar
Pontarotti G (2015) Расширенное наследование с организационной точки зрения. Hist Philos Life Sci 37:430–448
Hist Philos Life Sci 37:430–448
Статья пабмед Google Scholar
Портин П. (1993) Концепция гена: краткая история и современное состояние. Q R Biol 68:173–223
Статья КАС Google Scholar
Пишо А. (1993) История понятия жизни. Галлимар, Париж
Google Scholar
Pray L (2008) Репликация ДНК и причины мутации. Природоведение 1(1):214
Google Scholar
Радж А., ван Ауденарден А. (2008) Природа, воспитание или случайность: стохастическая экспрессия генов и ее последствия. Ячейка 135(2):216–226
Артикул КАС пабмед ПабМед Центральный Google Scholar
Richards C, Bossdorf O, Pigliucci M (2010) Какую роль играет наследственная эпигенетическая изменчивость в фенотипической эволюции? Биологические науки 60(3):232–237
Статья Google Scholar
Romanes GJ (1888) Ламаркизм против дарвинизма. Природа 38 (август): 413–413. https://doi.org/10.1038/038413a0
Природа 38 (август): 413–413. https://doi.org/10.1038/038413a0
Статья Google Scholar
Розен Р. (1991) Сама жизнь. Комплексное исследование природы. Издательство Колумбийского университета, Нью-Йорк, Origin and Fabrication of Life
Google Scholar
Слаткин М. (2009) Эпигенетическое наследование и проблема отсутствия наследуемости. Генетика 182(3):845–850
Артикул пабмед ПабМед Центральный Google Scholar
Сото А., Лонго Г., Микель П.А., Монтевиль М., Моссио М., Перре Н., Пошевиль А., Зонненшайн С. (2016) На пути к теории организмов: три основополагающих принципа в поисках полезной интеграции. Prog Biophys Mol Biol 122(1):77–82
Артикул пабмед ПабМед Центральный Google Scholar
Spencer H (1864) Принципы биологии Том 1. Williams and Norgate, Edinburgh
Williams and Norgate, Edinburgh
Stearns FW (2010) Сто лет плейотропии: ретроспектива. Генетика 186 (3): 767–773. https://doi.org/10.1534/genetics.110.122549. Опечатка в: Генетика. 2011 Январь; 187 (1): 355. PMID: 21062962; PMCID: PMC2975297
Стюарт Дж. (2004) Жизнь существует-т-эль?: Согласование генетики и биологии. Вюибер, Париж
Google Scholar
Stotz K (2008) Ингредиенты для постгеномного синтеза природы и воспитания. Филос Психол 21 (3): 359–381
Артикул Google Scholar
Stotz K (2017) Почему строительство ниши в целях развития не является селективным строительством ниши: и почему это важно. Интерфейс Focus 7:20160157
Артикул пабмед ПабМед Центральный Google Scholar
Шустар П. (2007) Понятие Крика о генетической информации и «центральная догма» молекулярной биологии. Br J Philos Sci 58 (1): 13–24
Br J Philos Sci 58 (1): 13–24
Артикул Google Scholar
Sutton WS (1903) Хромосомы в наследственности. Биол Булл 4(5):231–250
Артикул Google Scholar
Треротола М., Релли В., Симеоне П., Альберти С. (2015) Эпигенетическое наследование и отсутствующая наследуемость. Hum Genomics 9(1):1–12
Статья КАС Google Scholar
Урри Л.А., Рис Дж.Б., Каин М.Л., Вассерман С.А., Минорский П.В. (2016) Биология Кэмпбелла, 11-е изд. Пирсон, Нью-Йорк
Google Scholar
Waddington CH (1939) Введение в современную генетику. Macmillan, New York
Waddington CH (1942) Эпигенотип. Стремление 1:18–20
Google Scholar
Waddington CH (1953) Генетическая ассимиляция приобретенного признака.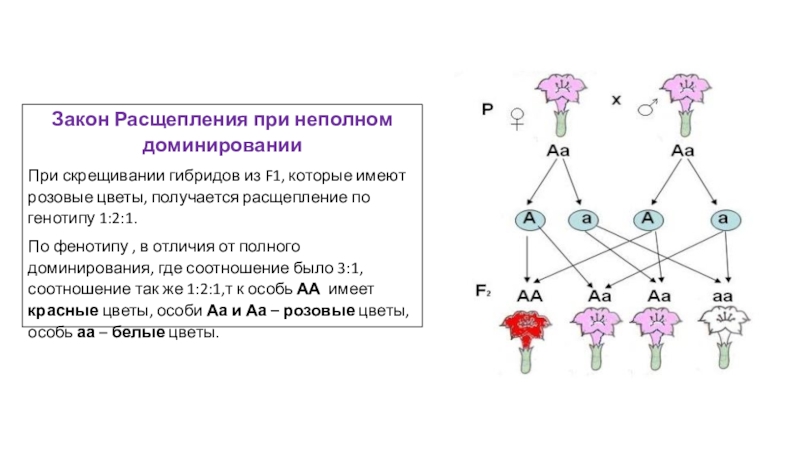 Эволюция 7(2):118–126
Эволюция 7(2):118–126
Артикул Google Scholar
Уолш Д.М. (2010) Два неодарвинизма. Hist Philos Life Sci 32 (2–3): 317–339
PubMed Google Scholar
Уолш Д. (2015) Организмы, деятельность и эволюция. Издательство Кембриджского университета, Кембридж
Книга Google Scholar
Watson J, Crick F (1953a) Молекулярная структура нуклеиновых кислот. Природа 171:737–738
Артикул КАС пабмед Google Scholar
Watson J, Crick F (1953b) Генетические последствия структуры дезоксирибонуклеиновой кислоты. Природа 171:964–967
Статья КАС пабмед Google Scholar
Вейсманн А (1883) [1990]. Де л’Эредите. В: Lenay C (ed) La découverte des lois de l’hérédité (1862–1900): une anthologie.