Ребят, помогите с летним дз по биологии, плиз! Мне нужно собрать гербарий листья простые и
Классификация такая и довольно сложная.
На простые и сложные листья делятся по количеству листовых пластинок.
• Простые — одна листовая пластина в листе,
• сложные — несколько листовых пластинок в листе. У каждой пластинки — свой черешок,прикрепляющийся к общему черешку (далее поймёте).
Простые листья.
Делятся на 2 группы:
1. Листья с цельной листовой пластиной,
2. листья с расчленённой листовой пластиной.
Простые листья с цельной листовой пластиной будут различаться по:
• Форме листовой пластины,
• краю листа,
• форме верхушки,
• основанию листа.
Как оно выглядит и примеры растений в файле. Для примера простого листа с цельной листовой пластиной сирень нарвите.
Простые листья с расчленённой листовой пластиной сортируются по следующим признакам:
1. Форма листовой пластинки:
• Если форма округлая и доли листа напоминают расставленные пальцы ладони ,то такие листья — пальчатые.
• Если форма листа овальная и доли листа напоминают перо птицы, то такие листья — перистые.
• Если расчленение происходит на глубину,меньшую,чем половина расстояния от края листа до центра — листья лопастные.
Пальчато-лопастные (клён),
Перисто-лопастные (дуб).
Клён,дуб в подмосковье есть — нарвёте.
• Если расчленение происходит на глубину большую половины расстояния от края до центра — листья раздельные:
Пальчато-раздельные (клещевина),
перисто-раздельные (одуванчик) уж его найдёте явно.
• Если расчление происходит до центра листовой пластины или до центральной жилки (у перистых) листья — рассеченные:
Пальчато-рассеченные (лютик),
Перисто-рассеченные (валериана) вряд ли встретите…
Сложные листья.
1. Тройчатосложные (клевер,земляника) их найдёте,клевер 100% землянику ток если в лесок или парк,
2. Пальчатосложные (конский каштан) простым языком это обычный каштан он на каждой улице,
3.
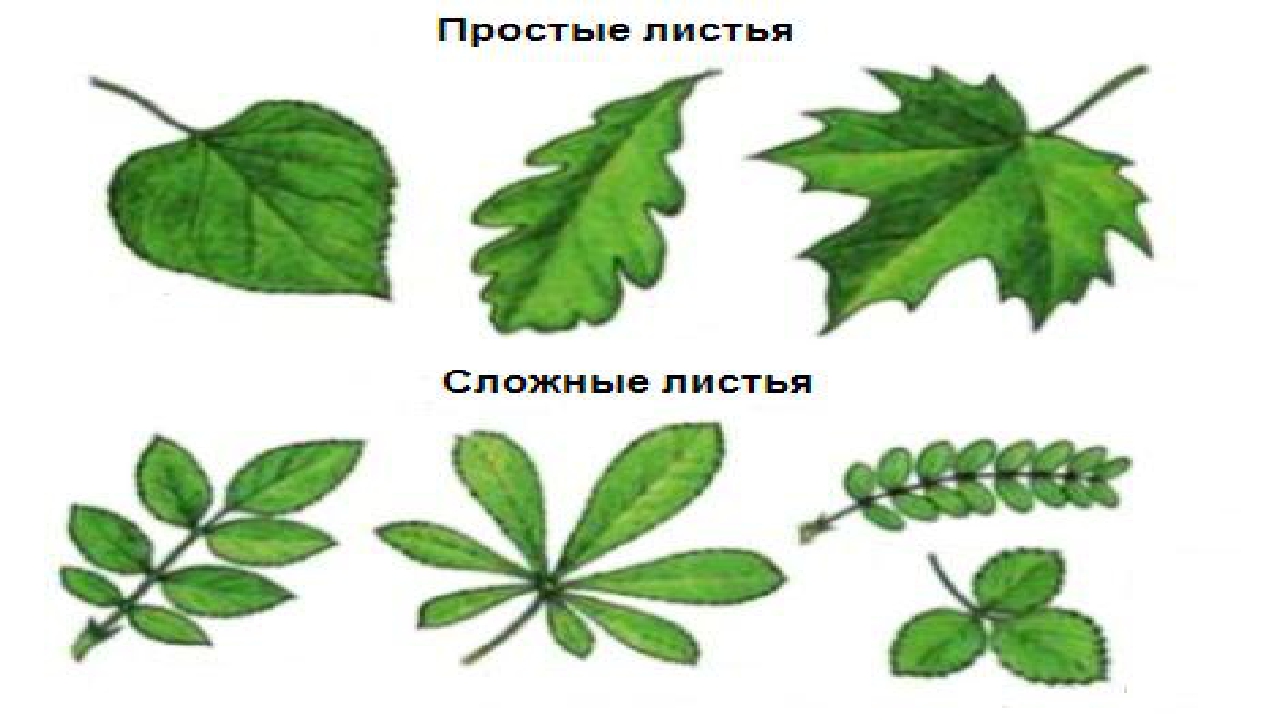 Перистосложные делятся на две подгруппы:
Перистосложные делятся на две подгруппы:а) непарноперистые — лист заканчивается непарным листом на верхушке (шиповник,рябина) их тож увидите — нарвёте,
б) парноперистые непарный лист на верхушке отсутствует (акация,горох) горох вряд ли разумеется,а акация белая вроде есть в подмосковье,вы главное на листья смотрите.
Все примеры че да как выглядит в файлах двух.
Первый файл простые с нерасчлененной пластиной,второй с расчленённой,во втором файле внизу сложные листья.
§ 6. Внешнее строение листа
Вопросы в начале параграфа1. Какие вегетативные органы различают у цветкового растения?
Для цветковых растений характерны вегетативные органы: корень, стебель и лист.
2. На каком органе цветкового растения расположены листья?
Листья у цветковых растений расположены на стебле.
Одинаковы ли размеры и форма листьев у разных растений?
Листья разных растений отличаются друг от друга и по форме, и по размеру, и по цвету, и по многим другим параметрам. При этом у всех листьев есть много общего.
Лабораторные работы
Лабораторная работа: Листья простые и сложные, их жилкование и листорасположение
1. Рассмотрите листья комнатных растений и образцов из гербария. Отберите простые листья. По какому признаку вы их отбираете?
Главный признак простых листьев — всего одна листовая пластина.
К комнатным растениям с простыми листьями относятся следующие растения:
2. Отберите сложные листья. По какому признаку вы это делаете? Какое жилкование у отобранных вами листьев?
Главный признак сложных листьев — наличие нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
Жилкование у всех отобранных листьев сетчатое — то есть сетчатым жилкованием отличаются листья ясеня, кислицы, земляники, клевера, рябины, шиповника, акации, вязеля, каштана, люпина и фасоли.
На территории России произрастают следующие растения со сложными листьями:
3. Какое листорасположение имеют просмотренные вами растения?
- Очередное — хлорофитум, гибискус, каланхоэ, герань, фикус, аспидистра, сансевиерия, гиацинт, юкка, рябина, шиповник, вязель, люпин;
- Супротивное — спатифиллум, аукуба, ясень, кислица, земляника, клевер, акация, каштан, фасоль;
- Мутовчатое — орхидея.
4. Заполните таблицу «Строение и расположение листьев у разных растений».
«Строение и расположение листьев у разных растений»
Название растения | Листья простые или сложные | Жилкование | Листорасположение |
Хлорофитум | Простые | Параллельное | Очередное |
Гибискус | Простые | Сетчатое | Очередное |
| Каланхоэ | Простые | Сетчатое | Очередное |
Герань | Простые | Сетчатое | Очередное |
| Спатифиллум | Простые | Сетчатое | Супротивное |
Фикус | Простые | Сетчатое | Очередное |
| Аспидистра | Простые | Дуговое | Очередное |
Аукуба | Простые | Сетчатое | Супротивное |
| Сансевиерия | Простые | Параллельное | Очередное |
Гиацинт | Простые | Параллельное | Очередное |
| Орхидея | Простые | Дуговое | Мутовчатое |
Юкка | Простые | Параллельное | Очередное |
| Ясень | Сложные | Сетчатое | Супротивное |
| Кислица | Сложные | Сетчатое | Супротивное |
Земляника | Сложные | Сетчатое | Супротивное |
| Клевер | Сложные | Сетчатое | Супротивное |
Рябина | Сложные | Сетчатое | Очередное |
| Шиповник | Сложные | Сетчатое | Очередное |
Акация | Сложные | Сетчатое | Супротивное |
| Вязель | Сложные | Сетчатое | Очередное |
Конский каштан | Сложные | Сетчатое | Супротивное |
Люпин | Сложные | Сетчатое | Очередное |
| Фасоль | Сложные | Сетчатое | Супротивное |
Вопросы в конце параграфа
1. Каково внешнее строение листа?
Каково внешнее строение листа?
Листья большей части растений (берёза, вишня, клён, яблоня) состоят из двух частей: листовой пластины и черешка. Это черешковые листья.
Листья некоторых растений (пшеница, цикорий, алоэ, лён) не имеют черешка а прикрепляются к стеблю основанием листовой пластины. Это сидячие листья.
Кроме того у некоторых листьев в основании развиваются небольшие выросты — прилистники.
По форме листья могут быть округлыми, овальными, сердцевидными, игольчатыми и т.д. Также листья могут отличаться по форме края пластины — лист может иметь зубчатый край, пильчатый, городчатый, цельный и т.д.
2. Какие листья называют сложными, а какие — простыми?
Листья называются простыми, если они состоят из одной листовой пластины.
Листья называют сложными, если состоят из нескольких листовых пластин соединённых с общим черешком небольшими черешками.
3. Как однодольные растения отличаются от двудольных по жилкованию листьев?
Для однодольных растений характерно параллельное или дуговое жилкование.
Для двудольных растений характерно сетчатое жилкование.
4. Какую функцию выполняют жилки листа?
Жилки — это проводящие пучки листа. Они проводят растворы питательных веществ поступающие из стебля, а также добавляют листьям прочность.
Подумайте
Можно ли только по жилкованию листьев определить, какое это растение — однодольное или двудольное?
Нет, нельзя. И среди однодольных, и среди двудольных растений существуют исключения. Например, у однодольного вороньего глаза жилкование сетчатое, а у двудольного подорожника — дуговое.
Задания
Составьте гербарий листьев с различной формой листовых пластинок и разным жилкованием.
Словарик
Листовая пластинка — это основная часть листа, выполняющая главные функции листа: фотосинтез, газообмен и испарение воды.
Черешок — это часть листа, которая соединяет листовую пластину со стеблем.
Черешковые листья — это листья, в строении которых есть черешок для соединения листовой пластины со стеблем.
Сидячие листья — это листья, в строении которых черешок отсутствует, а листья к стеблю крепятся основанием листовой пластины.
Простые листья — это листья состоящие из одной листовой пластины.
Сложные листья — это листья состоящие из нескольких листовых пластин и соединённых с общим черешком небольшими черешками.
Сетчатое жилкование — это жилкование листьев, жилки на которых многократно ветвятся и образуют сплошную сеть.
Параллельное жилкование — это жилкование листьев, жилки на которых расположены параллельно друг другу.
Дуговое жилкование — это жилкование листьев, жилки на которых расположены по дуге симметрично относительно центральной линии листа.
» Листья простые и сложные» ( 7класс)
Государственное казенное общеобразовательное учреждение «Школа № 3 Костромской области для детей с ограниченными возможностями здоровья»
Конспект урока
по биологии
На тему: «Листья простые и сложные»
для обучающихся 7 класса
Подготовила учитель
биологии высшей категории
Лебедева Оксана Арнольдовна
Цель: Формирование представлений о листьях простых и сложных.
Задачи:
Коррекционно-образовательные:
Познакомить обучающихся с простыми и сложными листьями;
Выявить общие и отличительные признаки строения простых и сложных листьев;
Учить различать простые и сложные листья на различных объектах;
Коррекционно-развивающие:
Развивать речь, память, внимание, мышление через умение сравнивать, обобщать, делать выводы;
Способствовать развитию навыков самостоятельной работы, совершенствование практических навыков;
Развивать познавательную активность, умение работать с гербарием, живыми объектами.


Коррекционно-воспитательные:
Способствовать воспитанию природоохранительного поведения;
Вырабатывать мотивацию к изучению биологии.
Методическое обеспечение урока:
карточки с заданиями разной степени сложности для самостоятельной работы учащихся,
гербарий с простыми и сложными листьями,
комнатные растения
презентация « Листья простые и сложные»
карточки с опорными словами
конверты с листьями
Тип урока: Комбинированный
Деятельность учителяДеятельность учащихся
Коррекционная работа
I Организационный момент Психологический настрой
(1-2 мин) Здравствуйте, ребята. Говорят, как день начнешь, так его и проведёшь. Давайте начнем сегодняшний день с улыбки, улыбнемся друг другу и с таким настроением проведём весь урок. Мне хочется, чтобы вы были на уроке внимательны, находчивы, а главное – чтобы вы показали, что знаете и как умеете работать. Проверка готовности класса.
Встают у парт, приветствуют учителя и гостей.
Рефлексия. Коррекция эмоционально-волевой сферы.
II. Актуализация знаний (1 мин)
Мы продолжаем знакомиться со строением цветкового растения.
1.Загадка
Для того, чтобы узнать о какой части растения мы будем говорить на уроке, отгадаем загадку
Под ногами лежат,
Зашагаешь — шуршат,
Яркие, цветные,
Краешки резные.
Ответы учащихся.
Листья.
Развитие логического мышления, через отгадывание загадки.
III Повторение пройденного материала ( 7-10 мин)
На прошлом уроке мы уже говорили о листьях, прежде чем познакомиться с новым материалом, вспомним, что уже знаем о листьях.
1.Беседа
Лист, это какой орган наземный или подземный?
Где он располагается?
Назовите основные части листа
По наличию черешка, на какие группы делятся листья?
Опорные слова вывешиваются на доске ( черешковые, сидячие)
Лист это наземный орган.
Лист располагается на стебле.
Широкая часть листа- листовая пластинка. Узкая часть листа- черешок.
Черешковые и сидячие.
Коррекция и развитие связной речи (полные ответы учащихся).
Коррекция памяти (выяснение степени усвоения материала предыдущих уроков).
2.Практическая работа
Определить на примере комнатных растений способы прикрепления листьев к стеблю
(герань, колеус, роициссус, традесканция, сеткреазия, столетник)
Определят черешковые и сидячие листья на примере комнатных растений.
Развитие познавательной активности, через умение работать с живыми объектами
Продолжение беседы
Что можно увидеть на обратной стороне листа?
Вспомните, какие виды жилкования листьев вы знаете?
Опорные слова вывешиваются на доску (сетчатое, параллельное, дуговое)
На обратной стороне листа расположены жилки.
Сетчатое, параллельное, дуговое.
Развитие долговременной памяти.
3.Самостоятельная работа
Определить жилкование листьев
Работа по карточкам
Молодцы, хорошо справились со всеми заданиями.
Выполняют задание.
Развитие самостоятельности и при выполнении задания.
IV. Изучение нового материала.(15 мин.)
1.Практическая работа
В конверте у детей – листья березы и рябины.
Назвать к каким растениям относятся листья.
Сравните:
Что общего есть у этих листьев?
Чем отличаются эти листья?
Вывод:
Если на черешке одна листовая пластинка—простой лист.
Если на черешке несколько листовых пластинок—сложный лист.
2.Запись темы урока на доске
Листья простые и сложные.
Сегодня на уроке мы будем учиться распознавать простые и сложные листья.
Узнают и называют, к каким растениям относятся листья
Есть черешок, листовая пластинка, жилки.
Разное количество пластинок на черешке
Слушают и запоминают.
Записывают тему урока в тетради.
Развитие логического мышления, через операции сравнения, обобщения.
Развитие зрительной и слуховой памяти
3.Работа с учебником
Читают текст на стр.71
Рассмотрите простые листья на рис.52
Рассмотрите сложные листья на рис.53.
Читают текст учебника.
Перечисляют, к каким растениям относятся простые листья.
Перечисляют сложные листья.
Контроль посадки за партами, сохранение осанки.
Развитие слухового и зрительного восприятия, внимания, памяти, активизация словаря.
4.Работа по презентации
Первичное закрепление
Читают текст по презентации
Развитие памяти
5.Фронтальная работа по презентации
Среди представленных листьев выберите номера простых листьев
Называют номера простых листьев
Развитие логического мышления, через операцию выбора.
V. Физминутка ( 1-2 мин)
(отдых для глаз)
Посмотрите в окно, можно ли сейчас увидеть на деревьях листья, если увидели, задержите свое внимание на них, посмотрите вдаль.
Смотрят вдаль.
Здоровьесбережение, профилактика нарушений осанки и зрения; снятие статического напряжения.
VI. Закрепление изученного материала.( 10мин)
1.Работа с гербарием.
В простенках в классе развешены листья из гербария.
Определить по гербарию простой лист или сложный.
Определяют по гербарию, какой лист простой или сложный.
Использование приемов технологии Базарного.
Развитие мышления через операцию классификация по признакам.
2.Самостоятельная работа
а).Для сильных учеников без опоры на наглядность, работа с перфокартой «Простые и сложные листья»
б).Для более слабых учеников, задание на карточке с опорой на наглядность.
Определить простые и сложные листья.
Выполняют задания.
Развитие самостоятельности и наблюдательности при выполнении задания.
Дифференцированный подход: задания разного уровня сложности
VII. Подведение итогов урока. Рефлексия. ( 1мин)
Чему учились сегодня на уроке?
Какое задание понравилось?
С каким настроением уходим с урока?
Учились различать простые и сложные листья.
Отвечают на вопросы.
Развитие мышления, памяти, речи. Развитие умения рассуждать, обобщать, анализировать свою работу на уроке.
Развитие смысловой памяти, усвоение поставленной задачи.
VIII. Домашнее задание.( 1мин)
Прочитать учебник на стр.71-73.
Письменно выполнить задание №3 на стр.73
Записывают домашнее задание в дневник.
Развитие зрительной и слуховой памяти.
IX. Выставление оценок (1 мин)
Слушают комментарии учителя.
Развитие самооценки.
Лабораторная работа по теме6 «Листья простые и сложные»
Лабораторная работа № 6
Листья простые и сложные, их жилкование и листорасположение
Цель:
— изучить строение простых и сложных листьев у различных растений,
— познакомиться с различными типами жилкования листьев
Оборудование: гербарии различных растений, комнатные растения, простые и сложные листья различных растений.
Ход работы
1. Рассмотрите листья комнатных растений и образцов из гербария. Отберите простые листья. По какому признаку вы их отбираете? Дайте ответ на вопрос.
2. Отберите сложные листья. Назовите эти растения.
3. Сделайте рисунок сложных листьев и подпишите название растения и основные части листа.
4. Отберите простые листья. Назовите эти растения.
5. Сделайте рисунок простых листьев и подпишите название растения и основные части листа.
6. Заполните таблицу «Строение и расположение листьев у разных растений»:
Название растенияЛистья (простые или сложные)
Жилкование
Листорасположение
7. Сделайте вывод о проделанной работе.
Примеры простых листьев у хлорофитума, сенполии, бегонии, колеуса(слева на право).
Простые листья состоят из одной листовой пластинки.
2. Отберите сложные листья. По какому признаку вы это делаете? Какое жилкование у отобранных вами листьев?
Сложные листья состоят из нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
У фатсии сетчатое жилкование, у робусты – параллельное.
3. Какое листорасположение имеют просмотренные вами растения?
Очередное листорасположение имеют фатсия, бегония, сенполия; супротивное – колеус, мутовчатое листорасположение – робуста, хлорофитум.
4. Заполните таблицу «Строение и расположение листьев у разных растений».
1. Каково внешнее строение листа?
Большая часть листьев состоит из двух частей: листовой пластинки и черешка. Черешок соединяет листовую пластинку со стеблем.
У листьев таких растений, как алоэ, пшеница, цикорий, лён, черешков нет, они прикрепляются к стеблю основанием листовой пластинки.
При основании черешка иногда развиваются выросты — прилистники
2. Какие листья называют сложными, а какие — простыми?
Какие листья называют сложными, а какие — простыми?
Сложные листья состоят из нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
Простые листья состоят из одной листовой пластинки.
3. Как однодольные растения отличаются от двудольных по жилкованию листьев?
Для однодольных растений характерно параллельное и дуговое жилкование.
Сетчатое жилкование типично для листьев двудольных растений.
4. Какую функцию выполняют жилки листа?
Жилки проводят растворы питательных веществ и придают листу прочность.
Подумайте
Можно ли только по жилкованию листьев определить, какое это растение — однодольное или двудольное?
Только по жилкованию листьев определить, какое это растение — однодольное или двудольное, нельзя, т.к. бывают исключения (например, у двудольного подорожника жилкование дуговое, а листья однодольного растения вороний глаз имеют сетчатое жилкование).
Листья простые и сложные. Листорасположение
admin 24.08.2010
Книга для чтения по ботанике
По своей форме листья различных растений не похожи друг на друга. Но даже самые разнообразные листья всегда можно объединить в две большие группы. Одну группу образуют простые листья, другую — сложные.
Как отличить простой лист от сложного? На черешке каждого простого листа только одна листовая пластинка. А сложные листья имеют несколько расположенных на одном черешке листовых пластинок, которые именуют листочками.
Среди простых листьев различают цельные, лопастные, раздельные и рассеченные.
Рис. 51. Листья сложные: 1 — малина; 2 — конский каштан; 3 — акация; 4 — земляника.
Цельные листья имеют многие деревья: береза, липа, тополь, яблоня, груша, вишня, черемуха, осина и другие. Лист считается цельным, если его пластинка цельнокрайняя или имеет неглубокие выемки.
Лопастным называют лист, у которого, как у дуба, вырезы-лопасти по краям пластинки доходят до одной четверти ее ширины.
Если надрезы листовой пластинки немного не доходят до средней жилки или основания листа, листья называют раздельными. Если же лист разрезан до средней жилки или до основания, он носит название рассеченного.
Лопастные листья — это листья клена, дуба, боярышника, смородины, крыжовника и некоторых других растений.
Возьмите несколько листьев разных растений, например: малины, рябины, ясеня, тополя, клена, дуба. Сравните листья рябины, малины, ясеня с листьями тополя, липы, клена и дуба. Чем они отличаются друг от друга? У листьев ясеня, рябины и малины на одном черешке расположено несколько листовых пластинок — листочков. Это сложные листья. Листья тополя, клена и дуба простые. У простых листьев листовая пластинка во время листопада отпадает вместе с черешком, а у сложных — отдельные листочки, составляющие лист, могут опадать раньше, чем черешок.
Сложный лист, состоящий из трех листовых пластинок, как у клевера, называют тройчатосложным или тройчатым.
Если лист образован несколькими листовыми пластинками, прикрепляющимися в одной точке, как, например, у люпина, его называют пальчатосложным. Если же листочки сложного листа прикрепляются по всей длине черешка, то такой лист — перисто-сложный.
Среди перистосложных листьев различают непарноперистые и парноперистые.
Непарноперистые листья — это те, что заканчиваются листовой пластинкой, не имеющей своей пары. Примером непарноперистых листьев будут листья рябины, ясеня, малины. Парноперистосложные листья встречаются реже, но все же некоторые растения с такими листьями вам известны. Это, например, горох посевной, мышиный горошек и душистый горошек.
Как простые, так и сложные листья двудольных и однодольных растений располагаются на стеблях в определенном порядке. Участки стебля, несущие лист, называются стеблевыми узлами, а участки стебля между узлами — междоузлиями.
Расположение листьев на стебле называют листорасположением.
Рис. 52. Листорасположение: 1 — очередное; 2 — супротивное; 3 — мутовчатое; а — междоузлие; б — узел.
Большинство растений имеет очередное листорасположение, например: рожь, пшеница, береза, яблоня, подсолнечник, фикус, роза. Листья у них расположены спирально вокруг стебля по одному, как бы чередуясь друг с другом, поэтому такое расположение и называют очередным.
Листья сирени, жасмина, клена, фуксии, глухой крапивы расположены на стебле не по одному, а по два: один лист против другого. Такое листорасположение называют супротивным.
Иногда встречаются растения с мутовчатым листорасположением. У них листья растут на стебле пучками, мутовками, располагаясь по три и более- листа в узле, и образуют как бы кольцо (мутовку) вокруг стебля. Среди комнатных растений мутовчатое листорасположение имеет олеандр, в аквариуме — элодея, среди диких растений — подмаренник северный, клевер люпиновый, вороний глаз четырехлистный и другие травянистые растения.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Просмотров: 29 412
Какие листья у алоэ простые или сложные — MOREREMONTA
Лист — это важнейший орган растения, главной его функцией является фотосинтез, т. е. синтез органических веществ из неорганических. Однако по внешнему строению листья растений разных видов отличаются. По форме листа часто можно определить, какому виду растений он принадлежит. Разнообразие внешнего строения листьев связано главным образом с тем, что растения приспособлены к разным условиям жизни.
Листья растений отличаются по размеру. Самые маленькие листья имеют размер меньше сантиметра (мокрица, ряска). Огромные листья характерны для некоторых тропических растений. Так у водного растения виктории диаметр листьев больше метра.
Во внешнем строении листьев большинства растений выделяют листовую пластинку и черешок. В листовой пластинке находится преимущественно фотосинтезирующая ткань, а черешок служит для соединения листовой пластинки со стеблем. Однако у некоторых видов растений листья не имеют черешков. Листья с черешками характерны для большинства деревьев (клен, липа, береза и др.). Листья без черешков свойственны алоэ, пшенице, кукурузе и др.
В листовой пластинке находится преимущественно фотосинтезирующая ткань, а черешок служит для соединения листовой пластинки со стеблем. Однако у некоторых видов растений листья не имеют черешков. Листья с черешками характерны для большинства деревьев (клен, липа, береза и др.). Листья без черешков свойственны алоэ, пшенице, кукурузе и др.
При внешнем осмотре листа на нем хорошо видны так называемые жилки. Лучше они видны на нижней стороне листа. Жилки образованы проводящими пучками и механическими волокнами. По проводящей ткани двигаются от корней вода и минеральные вещества, а в обратную сторону, от листьев, органические вещества. Механическая ткань придает листьям прочность и жесткость.
Жилкование листьев
При параллельном жилковании жилки в листовой пластине располагаются параллельно друг другу и выглядят как прямые линии.
При дуговом жилковании расположение жилок похоже на параллельное, но чем дальше от центральной оси листовой пластинки, тем больше жилка имеет форму дуги, а не прямой.
Параллельное и дуговое жилкование характерно для многих однодольных растений. Так многие злаки (пшеница, рожь) и лук имеют параллельное жилкование, а ландыш — дуговое.
При сетчатом жилковании жилки в листе образуют ветвящуюся сеть. Такое жилкование характерно для многих двудольных растений.
Существуют и другие типы жилкования листьев.
Простые и сложные листья
В зависимости от количества листовых пластин на одном черешке листья делят на простые и сложные.
У простых листьев на одном черешке развивается только одна листовая пластина (береза, осина, дуб).
У сложных листьев от одного общего черешка отрастают несколько или множество листовых пластин; при этом у каждого такого листочка есть свой маленький черешок, который его соединяет с общим черешком. Примерами растений со сложными листьями являются рябина, акация, земляника.
Сложный лист розы Простой лист осины
Расположение листьев
На стебле растения выделяют узлы и междоузлия. От узлов отрастают листья, а междоузлия — это участки стебля между узлами. Расположение листьев на стебле может быть различным в зависимости от вида растения.
Если листья располагаются по одному в узлах, при этом все вместе листья дают вид расположения как бы по спирали вдоль стебля, то говорят об очередном расположении листьев. Такое расположение характерно для подсолнечника, березы, шиповника.
При супротивном расположении листья растут по два в каждом узле, друг напротив друга. Супротивное расположение встречается у клена, крапивы и др.
Если в каждом узле растут более двух листьев, то говорят о мутовчатом листорасположении. Оно свойственно, например, для элодеи.
Существует также розеточное расположение листьев, когда междоузлий почти нет, а все листья растут как бы из одного места по кругу.
1. Какие вегетативные органы различают у цветкового растения?
Для цветковых растений характерны вегетативные органы: корень, стебель и лист.
2. На каком органе цветкового растения расположены листья?
Листья у цветковых растений расположены на стебле.
Одинаковы ли размеры и форма листьев у разных растений?
Листья разных растений отличаются друг от друга и по форме, и по размеру, и по цвету, и по многим другим параметрам. При этом у всех листьев есть много общего.
Лабораторные работыЛабораторная работа: Листья простые и сложные, их жилкование и листорасположение
1. Рассмотрите листья комнатных растений и образцов из гербария. Отберите простые листья. По какому признаку вы их отбираете?
Главный признак простых листьев — всего одна листовая пластина.
К комнатным растениям с простыми листьями относятся следующие растения:
2. Отберите сложные листья. По какому признаку вы это делаете? Какое жилкование у отобранных вами листьев?
Отберите сложные листья. По какому признаку вы это делаете? Какое жилкование у отобранных вами листьев?
Главный признак сложных листьев — наличие нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
Жилкование у всех отобранных листьев сетчатое — то есть сетчатым жилкованием отличаются листья ясеня, кислицы, земляники, клевера, рябины, шиповника, акации, вязеля, каштана, люпина и фасоли.
На территории России произрастают следующие растения со сложными листьями:
3. Какое листорасположение имеют просмотренные вами растения?
- Очередное — хлорофитум, гибискус, каланхоэ, герань, фикус, аспидистра, сансевиерия, гиацинт, юкка, рябина, шиповник, вязель, люпин;
- Супротивное — спатифиллум, аукуба, ясень, кислица, земляника, клевер, акация, каштан, фасоль;
- Мутовчатое — орхидея.
4. Заполните таблицу «Строение и расположение листьев у разных растений».
«Строение и расположение листьев у разных растений»
Название растения
Листорасположение
Вопросы в конце параграфа1. Каково внешнее строение листа?
Каково внешнее строение листа?
Листья большей части растений (берёза, вишня, клён, яблоня) состоят из двух частей: листовой пластины и черешка. Это черешковые листья.
Листья некоторых растений (пшеница, цикорий, алоэ, лён) не имеют черешка а прикрепляются к стеблю основанием листовой пластины. Это сидячие листья.
Кроме того у некоторых листьев в основании развиваются небольшие выросты — прилистники.
По форме листья могут быть округлыми, овальными, сердцевидными, игольчатыми и т.д. Также листья могут отличаться по форме края пластины — лист может иметь зубчатый край, пильчатый, городчатый, цельный и т.д.
2. Какие листья называют сложными, а какие — простыми?
Листья называются простыми, если они состоят из одной листовой пластины.
Листья называют сложными, если состоят из нескольких листовых пластин соединённых с общим черешком небольшими черешками.
3. Как однодольные растения отличаются от двудольных по жилкованию листьев?
Для однодольных растений характерно параллельное или дуговое жилкование.
Для двудольных растений характерно сетчатое жилкование.
4. Какую функцию выполняют жилки листа?
Жилки — это проводящие пучки листа. Они проводят растворы питательных веществ поступающие из стебля, а также добавляют листьям прочность.
ПодумайтеМожно ли только по жилкованию листьев определить, какое это растение — однодольное или двудольное?
Нет, нельзя. И среди однодольных, и среди двудольных растений существуют исключения. Например, у однодольного вороньего глаза жилкование сетчатое, а у двудольного подорожника — дуговое.
ЗаданияСоставьте гербарий листьев с различной формой листовых пластинок и разным жилкованием.
СловарикЛистовая пластинка — это основная часть листа, выполняющая главные функции листа: фотосинтез, газообмен и испарение воды.
Черешок — это часть листа, которая соединяет листовую пластину со стеблем.
Черешковые листья — это листья, в строении которых есть черешок для соединения листовой пластины со стеблем.
Сидячие листья — это листья, в строении которых черешок отсутствует, а листья к стеблю крепятся основанием листовой пластины.
Простые листья — это листья состоящие из одной листовой пластины.
Сложные листья — это листья состоящие из нескольких листовых пластин и соединённых с общим черешком небольшими черешками.
Сетчатое жилкование — это жилкование листьев, жилки на которых многократно ветвятся и образуют сплошную сеть.
Параллельное жилкование — это жилкование листьев, жилки на которых расположены параллельно друг другу.
Дуговое жилкование — это жилкование листьев, жилки на которых расположены по дуге симметрично относительно центральной линии листа.
Опубликовано 10.06.2017 по предмету Биология от Гость >>
Ответ оставил Гость
Алоэ: листорасположение очередное, листья простые, жилкование параллельное
Акация: листорасположение очередное, листья сложные, жилкование сетчатое
Нельзя всё время учиться. А для развлечения мы рекомендуем вам поиграть в отличную игру:
| Листья простые или сложные | Жилкование | Простые | Параллельное | Простые | Сетчатое | Очередное | |
| Каланхоэ | Простые | Сетчатое | Простые | Сетчатое | Очередное | ||
| Спатифиллум | Простые | Сетчатое | Простые | Сетчатое | Очередное | ||
| Аспидистра | Простые | Дуговое | Простые | Сетчатое | Супротивное | ||
| Сансевиерия | Простые | Параллельное | Простые | Параллельное | Очередное | ||
| Орхидея | Простые | Дуговое | Простые | Параллельное | Очередное | ||
| Ясень | Сложные | Сетчатое | |||||
| Кислица | Сложные | Сетчатое | Сложные | Сетчатое | Супротивное | ||
| Клевер | Сложные | Сетчатое | Сложные | Сетчатое | Очередное | ||
| Шиповник | Сложные | Сетчатое | Сложные | Сетчатое | Супротивное | ||
| Вязель | Сложные | Сетчатое | Сложные | Сетчатое | Сложные | Сетчатое | Очередное |
| Фасоль | Сложные | Сетчатое |
Параграф 6. Внешнее строение листа
1. Какие вегетативные органы различают у цветкового растения?
У цветкового растения различают следующие вегетативные органы: стебель, лист, корень.
2. На каком органе цветкового растения расположены листья?
Листья расположены на стебле.
3. Одинаковы ли размеры и форма листьев у разных растений?
Листья разных растений сильно отличаются по внешнему виду.
Лабораторная работа
Листья простые и сложные, их жилкование и листорасположение
1. Рассмотрите листья комнатных растений и образцов из гербария. Отберите простые листья. По какому признаку вы их отбираете?
Примеры простых листьев у хлорофитума, сенполии, бегонии, колеуса(слева на право).
Простые листья состоят из одной листовой пластинки.
2. Отберите сложные листья. По какому признаку вы это делаете? Какое жилкование у отобранных вами листьев?
Сложные листья состоят из нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
У фатсии сетчатое жилкование, у робусты – параллельное.
3. Какое листорасположение имеют просмотренные вами растения?
Очередное листорасположение имеют фатсия, бегония, сенполия; супротивное – колеус, мутовчатое листорасположение – робуста, хлорофитум.
4. Заполните таблицу «Строение и расположение листьев у разных растений».
Вопросы
1. Каково внешнее строение листа?
Большая часть листьев состоит из двух частей: листовой пластинки и черешка. Черешок соединяет листовую пластинку со стеблем.
У листьев таких растений, как алоэ, пшеница, цикорий, лён, черешков нет, они прикрепляются к стеблю основанием листовой пластинки.
При основании черешка иногда развиваются выросты — прилистники
2. Какие листья называют сложными, а какие — простыми?
Сложные листья состоят из нескольких листовых пластинок, соединённых с общим черешком небольшими черешками.
Простые листья состоят из одной листовой пластинки.
3. Как однодольные растения отличаются от двудольных по жилкованию листьев?
Для однодольных растений характерно параллельное и дуговое жилкование.
Сетчатое жилкование типично для листьев двудольных растений.
4. Какую функцию выполняют жилки листа?
Жилки проводят растворы питательных веществ и придают листу прочность.
Подумайте
Можно ли только по жилкованию листьев определить, какое это растение — однодольное или двудольное?
Только по жилкованию листьев определить, какое это растение — однодольное или двудольное, нельзя, т.к. бывают исключения (например, у двудольного подорожника жилкование дуговое, а листья однодольного растения вороний глаз имеют сетчатое жилкование).
Структура и эволюция листа| Цифровой атлас древней жизни
Избранные ссылки и дополнительная литература
Примечание: Издатель предоставляет бесплатный доступ к элементам, отмеченным зеленой звездочкой. Обратите внимание, что для повторного использования могут применяться ограничения.
Научные статьи и главы книг
* Дойл, Дж. 2013. Филогенетический анализ и морфологические инновации наземных растений. В B.A. Амвросий и М. Пуругганан (ред.), Эволюция формы растений. Ежегодные обзоры растений 45: 1–50. https://doi.org/10.1002/9781119312994.apr0486
* Melo, E., C.A. Сид Феррейра и Р. Грибель. 2019. Новый вид Coccoloba P. Browne (Polygonaceae) из бразильской Амазонки с исключительно большими листьями. Acta Amazonica 49. https://doi.org/10.1590/1809-4392201804771
* Ротвелл, G.W., S.E. Wyatt, A.M.F. Томеску. 2014. Эволюция растений на стыке палеонтологии и биологии развития: парадигма, ориентированная на организм. Американский журнал ботаники 101: 899–913. https://doi.org/10.3732/ajb.1300451
https://doi.org/10.3732/ajb.1300451
Schneider, H., A.R. Смит, К. Прайер. 2009. Действительно ли морфология расходится с молекулами при оценке филогении папоротника? Систематическая ботаника 34: 455–475. https://doi.org/10.1600/036364409789271209
Томеску, A.M.F. 2008. Мегафиллы, микрофиллы и эволюция развития листьев. Тенденции в растениеводстве 14: 5–12. https://doi.org/10.1016/j.tplants.2008.10.008
Tryon, R. 1960. Словарь терминов, относящихся к листу папоротника. Таксон 9: 104-109. https://doi.org/10.2307/1216244
* Васко А., Р.С. Моран, Б.А. Амвросий. 2013. Эволюция, морфология и развитие листьев папоротника. Frontiers in Plant Science 4, Статья 345. https://doi.org/10.3389/fpls.2013.00345
Книги, учебники и руководства
Коултер, Дж. М., К. Р. Барнс и Х. С. Cowles. 1910. Учебник ботаники для колледжей и университетов, вып.I. Морфология и физиология. Американская книжная компания, Нью-Йорк. Читайте бесплатно в Интернет-архиве: https://archive.org/details/textbookofbotan01coul
Эллис, Б., Д. К. Дейли, Л. Дж. Хики, К. Р. Джонсон, Дж. Д. Митчелл, П. Уилф и С. Л. Уинг. 2009. Учебник по архитектуре листа. Comstock Publishing Associates, Cornell University Press, Итака, Нью-Йорк.
Исав К. 1977. Анатомия семенных растений, 2-е изд. John Wiley & Sons, Inc., Нью-Йорк.
Эверт Р.Ф., С.Э. Эйххорн. 2013. Ворона Биология растений, 8-е изд. W.H. Freeman and Co., Нью-Йорк, Нью-Йорк.
Фостер, А.С., и Э.М. Гиффорд. 1974. Сравнительная морфология сосудистых растений, 2-е изд. W.H. Freeman and Co., Сан-Франциско.
Харрис, Дж. Г., и М. В. Харрис. 2001. Терминология идентификации растений, иллюстрированный глоссарий, 2-е изд. Издательство Спринг-Лейк, Спринг-Лейк, Юта.
Рабочая группа по архитектуре листьев (А. Эш, Б. Эллис, Л. Дж. Хики, К. Джонсон, П. Уилф и С. Винг).1999. Руководство по архитектуре листа: морфологическое описание и категоризация двудольных и однодольных покрытосеменных с сетчатыми жилками. Смитсоновский институт, Вашингтон, округ Колумбия.
Смитсоновский институт, Вашингтон, округ Колумбия.
Симпсон, М. 2010. Plant Systematics, 2 nd ed. Academic Press, Берлингтон, Массачусетс.
Уиллис, К.Дж., и Дж.К. МакЭлвейн. 2002. Эволюция растений, 2-е изд. Oxford University Press, Великобритания
Сайтов
* Гербарий Центра биоразнообразия Кофрина Университета Висконсин-Грин-Бей.Pteridophytes of Wisconsin: Папоротники и их союзники: Глоссарий избранной терминологии по птеридофитам. https://www.uwgb.edu/biodiversity/herbarium/pteridophytes/pteridophyte_glossary01.htm
* Лаборатория анатомии растений: Микрофотографии растительных клеток и тканей с пояснительным текстом (J.D. Mauseth): http://www.sbs.utexas.edu/mauseth/weblab/
* Лесная служба Министерства сельского хозяйства США: Структура папоротника. https://www.fs.fed.us/wildflowers/beauty/ferns/structure.shtml
Хороший образец — Австралийский национальный гербарий
Растения следует прессовать как можно скорее после сбора, до увядания.Образцы следует прижать и высушить между листами полупоглощающей бумаги (например, газеты), перемеженными листами толстого картона или легкой фанеры. Всю партию можно связать вместе в прессе для растений или аккуратно сложить грузы сверху. Важна хорошая циркуляция воздуха; хорошо прижимается к крышке автомобиля в хорошую погоду.
Чтобы дать вам как можно более точную идентификацию, нам необходимо знать:
- Где было собрано растение? Расстояние от города, по какой дороге и т. Д., И росло ли растение в густом кустарнике, нарушенном или полугородском кустарнике, на пастбище или в саду.Что это за страна для диких растений — влажная, сухая / кустарниковая, высокий лес, каменистые склоны, речная равнина и т. Д. Какие еще растения были вокруг? Также отметьте высоту, тип скалы, тип почвы, широту и долготу, если они известны.
- Как это выглядело? Было ли это дерево, куст, ниспадающий (плоский на земле) куст, или трава (недревесное растение), или альпинист? Какого цвета были цветы в свежем виде? Какого цвета и текстуры была кора? Неужели он корнеплод?
Обычно достаточно образцов длиной около 30 см.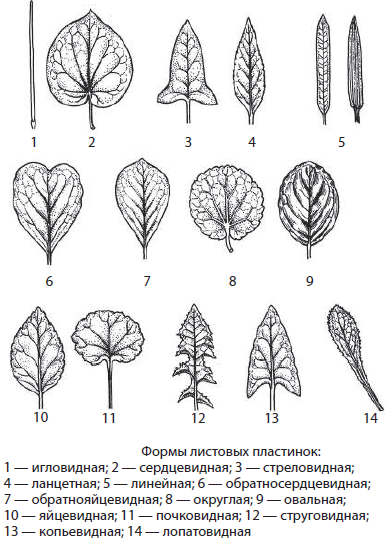 Они должны быть максимально полными; т.е. они должны включать бутоны, цветы и / или плоды, а также часть стебля с типичными листьями. Если форма или размер листьев на растении сильно различаются, сделайте заметку и возьмите отдельные части из разных частей растения, чтобы представить разницу. Для некоторых растений плоды и / или семена важны для точной идентификации; например Хакеа, ромашки и солончаки.
Они должны быть максимально полными; т.е. они должны включать бутоны, цветы и / или плоды, а также часть стебля с типичными листьями. Если форма или размер листьев на растении сильно различаются, сделайте заметку и возьмите отдельные части из разных частей растения, чтобы представить разницу. Для некоторых растений плоды и / или семена важны для точной идентификации; например Хакеа, ромашки и солончаки.
Некоторые группы предприятий трудно идентифицировать, и нам нужна специальная информация:
ЭвкалиптыСуществует более 800 видов, некоторые из которых очень трудно различить.Образцы должны включать взрослые листья, цветочные почки (лучше, чем открытые цветы!), Плоды и (если есть) молодые листья снизу на растении или от присосок у основания ствола. Вы должны сделать пометки с описанием:
- среда обитания (например, альпийская, вдоль берега реки, луга или леса, нарушенная территория)
- габитус (высотный, одно- или многоствольный)
- тип коры (гладкая, гладкая с каракулями, грубоватая, шероховатая, шелушащаяся, грубая и сильно ребристая и т. Д.).
Если кора даже частично шероховатая, как далеко она простирается (только носок у основания, до первой ветви, только по стволу и основным ветвям, а также по тонким веточкам)?
Если кора более или менее гладкая, какого цвета (цветов) и рисунка?
Акации
Еще больше видов, чем эвкалиптов. Соберите взрослые листья, молодые листья, если они есть, цветочные почки непосредственно перед открытием и, если возможно, зрелые стручки.
Грибки
Попытайтесь собрать ряд особей от незрелых до взрослых и записать цвета и субстрат.Выкопайте гриб целиком, включая основание стебля. Оберните пергаментной бумагой или поместите в коричневый бумажный (не пластиковый) пакет.
 Полезен отпечаток спор — срежьте шапочку взрослого человека (если есть) и положите жаберной стороной вниз на лист белой бумаги; накрыть тазом или ведром на пару часов, пока споры не упадут на бумагу; осторожно отправьте это вместе с образцом. Скорость важна, особенно для мясистых видов.
Полезен отпечаток спор — срежьте шапочку взрослого человека (если есть) и положите жаберной стороной вниз на лист белой бумаги; накрыть тазом или ведром на пару часов, пока споры не упадут на бумагу; осторожно отправьте это вместе с образцом. Скорость важна, особенно для мясистых видов.Травы (недревесные)
Для небольших трав соберите от одного до нескольких целых растений, включая любые подземные запасающие органы (луковицы или клубни), или, если растение является местным редким, оставьте их в земле и запишите, как эти части выглядели (это позволяет растению регенерировать — особенно актуально для редких видов, в том числе для многих орхидей).
Травы (и травянистые растения, включая камыш и осоки)
Соберите целое растение или небольшой пучок, включая корни. Что касается трав, обратите внимание, было ли растение легко выдергивать (однолетнее растение) или трудно вытащить (обычно многолетнее). Траву и осоку лучше всего собирать после того, как цветы распустятся, но до того, как плоды опадут.
Травяные деревья
Обратите внимание на длину цветущих и нецветущих частей колоса («копья») и длины ствола (если есть) под листьями.Следует собрать среднюю часть шипа (включая основание части цветения / плодоношения и верх гладкой части), а также несколько полных листьев, включая, если возможно, основания листьев. Будьте очень осторожны, чтобы не повредить растение при сборе листовых оснований; травянистые деревья (Xanthorrhoea) растут очень медленно и могут быть старше сотни лет.
Водные растения
Не хранить в воде после сбора. Слейте лишнюю воду и положите растение между газетными листами на несколько минут, затем прижмите и просушите между свежими листами.При необходимости замените бумагу.
Папоротники
Включает как плодородные (споровые), так и стерильные листья, а также часть корневища (ползучий «корень»), если он есть, и основание ножки (стебель листа).
 Для древовидных папоротников соберите часть плодородной вайи и основание ножки с чешуей или волосками.
Для древовидных папоротников соберите часть плодородной вайи и основание ножки с чешуей или волосками.Водоросли
Не будет идентифицирован гербарием ANBG. Небольшие (часто микроскопические) водоросли из цветущих рек или плотин можно собрать в водонепроницаемую емкость в воде, в которой они были обнаружены; Хранить в холодильнике.Их может определить местный орган управления водными ресурсами, который даст инструкции о том, как отправлять образцы. Крупные водоросли (например, водоросли) можно собирать так же, как и водоросли (см. Выше), и их следует отправлять в справочную службу, Национальный гербарий штата Новый Южный Уэльс, Mrs Macquaries Rd, Sydney NSW 2000.
Мхи, лишайники, печеночники
Постарайтесь включить плодовые тела (крошечные капсулы, диски или «зонтики» на стеблях или придатки сферической формы).Осторожно удалите несколько квадратных сантиметров растительного мата с субстрата или (если он плотно прилегает к коре, почвенной корке или камню) срежьте или сколите кусочки субстрата вместе с растением. Если образец большой или очень влажный, очень осторожно расплющите его. Не раздавливайте и не давите — поместите каждый образец в отдельный бумажный (не пластиковый) пакет с номером сбора и примечаниями и дайте ему высохнуть на воздухе. Хорошо упакуйте, чтобы не раздавить перед отправкой по почте.
Образцы для гербария
Ваши образцы, даже обычных видов, имеют научную ценность и могут быть включены в гербарий.верх
Распознавание контура листьев сложной формы
Получение изображения
Изображения листьев получаются с помощью трансиллюминатора или сканера для прозрачных пленок с фотографическим разрешением (300 точек на дюйм и уровни серого) и сохраняются в формате изображения без потерь (например, TIF, BMP или PNG). Форматы с потерями, такие как JPG, не подходят. Листья должны быть достаточно прижаты к сканеру, чтобы все части были в фокусе. Кроме того, листья не должны быть повреждены и в целом полностью развиваться.Однако критерий отбора листьев может изменяться в соответствии с целью исследования (таксономический, онтогенетический, эволюционный, связанный с геномом / транскриптомом и т. Д. [2]). Количество листьев зависит от уровня точности и необходимой погрешности, которая, в свою очередь, зависит от естественной изменчивости формы листьев, которая различается от вида к виду.
Листья должны быть достаточно прижаты к сканеру, чтобы все части были в фокусе. Кроме того, листья не должны быть повреждены и в целом полностью развиваться.Однако критерий отбора листьев может изменяться в соответствии с целью исследования (таксономический, онтогенетический, эволюционный, связанный с геномом / транскриптомом и т. Д. [2]). Количество листьев зависит от уровня точности и необходимой погрешности, которая, в свою очередь, зависит от естественной изменчивости формы листьев, которая различается от вида к виду.
Многоуровневая сегментация
На изображениях с просвечиваемым светом OL в результате представляют собой небольшие участки с более высокой плотностью по сравнению с остальной частью листовой пластинки (рис. 1C, 1D и 2A).Эти области могут быть выборочно сегментированы пороговыми методами на основе максимальной энтропии [9] (рис. 2B). Небольшие следы из-за толстых жилок могут быть удалены с помощью бинарной операции «закрытия». И наоборот, всю пластинку листа можно сегментировать с использованием автоматических (по умолчанию) пороговых методов, которые предполагают бимодальную гистограмму (рис. 2C). Если мы определим T1 инвертированную двоичную маску, полученную с помощью максимального порога энтропии, а T2 — двоичную маску, полученную с помощью порога по умолчанию, тогда операция XOR между T1 и T2 приведет к третьей маске, где OL, закрытые синусы и фон выглядят как отчетливые белые области, а остальная часть листовой пластинки — черная область.Однако в свете того факта, что все автоматизированные пороговые методы зависят от композиции изображения (т. Е. Соотношения между областями листа и фона) и освещенности (т. Е. Нелинейными настройками яркости и контрастности сканера), полученные значения T1 и Маски Т2 не могут быть оптимальными. Вместо этого может быть полезно иметь панель из нескольких изображений, подвергнутых XOR, с немного разными значениями T1 и T2. Например, как показано на рис. 2D, девять изображений получены с T1 ± 5% и T2 ± 5%, чтобы выбрать наиболее подходящую комбинацию. Все эти операции не касаются листьев, показывающих только TL, поскольку для этих листьев требуется только один порог по умолчанию (бимодальный).
Все эти операции не касаются листьев, показывающих только TL, поскольку для этих листьев требуется только один порог по умолчанию (бимодальный).
Получение и объединение нескольких контуров
Черно-белые контуры выбранной маски можно легко очертить с помощью классического инструмента «волшебная палочка» (рис. 3). Контуры автоматически сохраняются в виде массивов координат. Однако следует отметить, что координаты контура могут быть ориентированы по часовой стрелке или против часовой стрелки, в зависимости от положения контура, на который указывает «волшебная палочка».Этот момент имеет фундаментальное значение, поскольку для соединения контуров контуров TL и OL требуются разные ориентации и стратегии связывания. В частности, контуры OL показывают контур, который самопересекается два раза, следуя за своего рода слаломом через пару точек пересечения (рис. 3, красная рамка). И наоборот, контуры TL показывают контур, который не пересекается, а сам контактирует в соответствии с точкой касания (рис. 3, синяя рамка). Таким образом, сначала необходимо идентифицировать TL и OL, а затем на изображении локализовать точки пересечения и касания (рис. 4A).Следующая конкатенация контуров (рис. 4B) несколько сложна, но полностью автоматизирована. Алгоритм описан на рис. 5.
Рис 3. Определение точек пересечения и касания.
На изображении показаны семь контуров (помеченных от A до G), полученных из составной маски на рис. 1D. [A] — основная пластинка листа; [B] и [G] являются OL; [C] и [F] — пазухи листа, закрытые OL; [D] и [E] — пазухи листа, закрытые TL. В увеличенной красной рамке (упрощенно) зеленые и желтые стрелки показывают путь самопересечения или кривой слалома, которая объединяет контуры [A], [B] и [C], проходящие через две точки пересечения (красные точки).В увеличенной синей рамке зеленые и желтые стрелки показывают путь самоконтактной кривой, которая объединяет контуры [D] и [A], проходящую через точку касания (синяя точка).
https://doi. org/10.1371/journal.pone.0189427.g003
org/10.1371/journal.pone.0189427.g003
Рис. 4. Объединение контуров TL и OL.
(A) Расположение точек пересечения (красные точки) и точек касания (синие точки) между семью контурами на рис. 3. (B) Результат конкатенации контуров. Правильная связь контуров выделена последовательностью черных и зеленых следов.
https://doi.org/10.1371/journal.pone.0189427.g004
Рис. 5. Алгоритм конкатенации.
На схеме показаны ориентации координат (черные стрелки), необходимые для соединения соседних контуров ( A и B ), созданных двоичными контурами OL (левая панель) или TL (правая панель). Красные и синие точки обозначают точки пересечения и точки касания соответственно. Правильная ориентация контура и сцепление достигаются с помощью следующих шагов.
https://doi.org/10.1371/journal.pone.0189427.g005
Сначала мы идентифицируем в A и B :
- точки a c и b c , наиболее близкие к точке пересечения или касания
- точки a 1 и b 1 , предшествующие a c и b c , в индексе c ориентация -1)
- точки a 2 и b 2 , следующие за a c и b c , в ориентации контура 2 +1)
Пусть p будет отрезком линии, соединяющим точки a 1 и b 2, и q отрезком линии, соединяющим точки a 2 и б 1. Затем мы проверяем, пересекаются ли p и q .
Затем мы проверяем, пересекаются ли p и q .
Параметры регрессии сегментов p и q :
Общий диапазон координат p и q ограничен:
За исключением того, что в случае параллельности, прямые, которым принадлежат сегменты p и q , пересекаются в точке Z с координатами:
Но сегменты p и q пересекаются тогда и только тогда, когда координаты Z попадают в общий диапазон координат p и q , то есть:
На этом этапе контуры A и B , созданные OL, могут быть объединены, только если сегменты p и q пересекаются (красные стрелки).Если p и q не пересекаются, ориентация одного из двух контуров должна быть инвертирована. И наоборот, контуры A и B , созданные TL, могут быть объединены тогда и только тогда, когда p и q не пересекаются (синие стрелки). Если сегменты p и q пересекаются, ориентация одного из двух контуров должна быть инвертирована. В обоих случаях конкатенация достигается с помощью следующих шагов:
- контур A открывается в a c
- a c ссылки на b c
- координатная цепочка продолжает полный обход контура B , следуя его ориентации, до достижения b c снова
- b c снова подключается к a c
- координатная цепочка продолжает полный обход контура A , следуя его ориентации, до достижения a c снова
В результате получается новый замкнутый контур, который заменяет контуры A и B .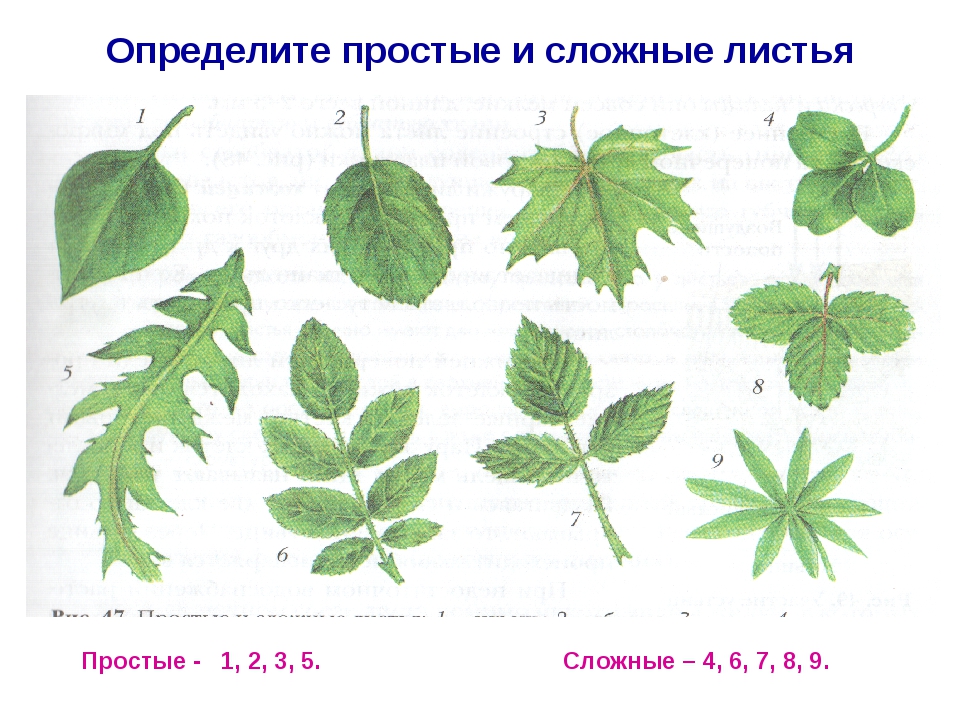 После каждой конкатенации количество контуров уменьшается на 1, так что алгоритм повторяется до тех пор, пока все контуры в конечном итоге не будут соединены для создания единого замкнутого контура листа.
После каждой конкатенации количество контуров уменьшается на 1, так что алгоритм повторяется до тех пор, пока все контуры в конечном итоге не будут соединены для создания единого замкнутого контура листа.
Чтобы оценить эффект рассмотрения или отказа от учета TL или OL, на рис. 6 показана коллекция листьев шести сортов виноградной лозы (A-F), демонстрирующих TL и OL, и шести сортов (G-L), не проявляющих TL или OL. Сорта A-F обрабатывались двумя способами: одним с использованием двойного порога, который распознает TL и OL (рис. 6, + столбцы), и другим с использованием метода порога по умолчанию, который не распознает OL или TL (рис. 6, — столбцы).Для статистического сравнения 90 контуров трех групп контуров (OL / TL +, OL / TL- и noOL / TL) были обработаны эллиптическим анализом Фурье (EFA) [10] и первые 20 эллиптических гармоник [3,11 ] были установлены в качестве входных переменных для анализа главных компонент (PCA), классического многомерного исследовательского метода. График PCA (рис. 7) четко отделяет группу OL / TL + от двух других групп, указывая на то, что TL и OL являются важными компонентами формы для различения двух групп сортов (A-F и G-L).Вклад эллиптических гармоник в формирование формы листа, включая TL и OL, можно визуализировать с помощью пошагового обратного преобразования эллиптических коэффициентов Фурье (рис. 8).
Рис. 6. Визуальное сравнение форм листьев, полученных с учетом или без учета TL или OL.
Листья шести сортов винограда, показывающих TL и OL [A-F], были получены с учетом (+ столбцы) или без учета (- столбцы) TL и OL. Для этого одни и те же изображения обрабатывались два раза: сначала с использованием метода двойного порога и контурирования, показанного в этой статье, а затем с использованием метода порогового значения по умолчанию.Другие сорта, не показавшие TL или OL [G-L], были обработаны с использованием метода пороговых значений по умолчанию.
https://doi.org/10.1371/journal.pone.0189427. g006
g006
Рис. 7. Статистическое сравнение форм листьев, полученных с учетом или без учета TL или OL.
Анализ главных компонентов первых 20 дескрипторов EFA (эллиптических гармоник) контуров 90 листьев, показанных на рис. 6. На графике показано резкое разделение форм листьев у сортов AF, достигнутых с учетом (красные точки) или без учета TL и OL (желтые точки).Последние смешаны с листьями сортов G-L, которые не показали TL или OL (синие точки).
https://doi.org/10.1371/journal.pone.0189427.g007
Рис. 8. Пошаговое обратное EFA-преобразование контура одного листа.
На изображении показана форма листа, повторно синтезированная с использованием первых 270 гармоник (гармоники с 1 по 15, следующие единичные шаги; гармоники с 60 по 270, следующие шаги 30). OL начинают появляться с 6 -й и 12 -й гармоники.
https://doi.org/10.1371/journal.pone.0189427.g008
До сих пор описанный метод можно также использовать для определения формы перекрывающихся листьев. Это, конечно, неуместно при работе со свежими образцами, но может представлять интерес для гербарных образцов, листья которых часто перекрываются, но их нельзя обрабатывать и перемещать из-за их жесткости и хрупкости. Однако следует отметить, что это приложение представляет две проблемы. Одним из факторов может быть контекстуальное присутствие в гербарных образцах объемных цветов и плодов, которые не позволяют использовать даже мягкое сжатие для расплющивания листьев на планшете для сканирования.Вторая проблема — относительная непрозрачность бумаги, на которой лежат образцы, что затрудняет просвечивание. Однако примерно в этот момент я обнаружил, что бумажные листы весом 120 г / м 2 или меньше позволяют получать достаточно хорошие изображения с помощью коммерческого сканера с увеличенной вручную яркостью.
Другие утилиты, присутствующие в макросе, позволяют повторно отслеживать контуры, накладывать их поверх и сохранять в виде растровых изображений или файлов координат. Координаты сохраняются в необработанном формате, таком же, как и в ImageJ, поэтому они могут быть напрямую импортированы и отображены ImageJ с помощью команд меню: Файл> Импорт> Координаты XY.Контуры тоже можно разделить. Это позволяет срезать черешки, отделить перекрывающиеся листья, прикрепленные к одной ветке (рис. 9), а также заделать небольшие разрывы границы листа (рис. 10). Полная блок-схема макроса показана на рис. 11. Макрос ImageJ / Fiji для получения сложных форм листьев доступен из файла S1. Макрос, выполняющий эллиптический анализ Фурье [10] для извлечения гармонических составляющих формы листьев [3,11,12], доступен в файле S2.
Координаты сохраняются в необработанном формате, таком же, как и в ImageJ, поэтому они могут быть напрямую импортированы и отображены ImageJ с помощью команд меню: Файл> Импорт> Координаты XY.Контуры тоже можно разделить. Это позволяет срезать черешки, отделить перекрывающиеся листья, прикрепленные к одной ветке (рис. 9), а также заделать небольшие разрывы границы листа (рис. 10). Полная блок-схема макроса показана на рис. 11. Макрос ImageJ / Fiji для получения сложных форм листьев доступен из файла S1. Макрос, выполняющий эллиптический анализ Фурье [10] для извлечения гармонических составляющих формы листьев [3,11,12], доступен в файле S2.
Рис. 9. Разделение листьев гербарных образцов.
(A) Оригинальный гербарный образец. (B) Двойная пороговая маска. (C) Полный контур образца. Пунктирными линиями обозначены точки разреза. (D) Контуры отдельных листьев, полученные с помощью функции обрезки и обведенные разными цветами. (E) Контуры отдельных листьев.
https://doi.org/10.1371/journal.pone.0189427.g009
Рис. 10. Утилита Cut.
Контуры можно разделить с помощью функции обрезки, чтобы удалить черешок с листа, «исправить» небольшие разрывы границы листа, а также отделить пересекающиеся листья, прикрепленные к одной и той же ветке, как показано на рис. 9.(A) Небольшой разрыв границы листа. Желтая пунктирная линия обозначает разделительную линию. (B) Получившийся «отремонтированный» контур.
https://doi.org/10.1371/journal.pone.0189427.g010
Рис. 11. Блок-схема макроса ImageJ.
На схеме показаны основные функции (показаны большими блоками, выделенными трехбуквенными акронимами жирным шрифтом) и основная последовательность операций (стрелки). Полный список операций, которые могут быть выполнены после завершения каждой функции, обозначен акронимами «СЛЕДУЮЩИЕ ОПЦИИ» под каждым блоком.Перезапустить макрос или выйти из него можно в любое время с помощью команды меню или клавиши ESC (не показана).
https://doi.org/10.1371/journal.pone.0189427.g011
О Herbaria 3.0 ~ Herbaria 3.0 Расскажите нам историю своего растения!
Гербарии 3.0 и рассказы о растениях
Растения есть повсюду в нашем мире и живут. Они украшают наши дома и укрепляют наши тела. Они дают нам лекарства и метафоры, духи и стихи. Но люди часто не замечают того факта, что растения существуют в мире сложных взаимоотношений, которые часто скрыты от человеческого взора.Они могут узнавать, общаться и даже сотрудничать друг с другом. Они проявляют сложное поведение в ответ на столь же сложные раздражители. У них есть свои желания, потребности и желания.
И все же, несмотря на наши существенные связи с зеленым миром, многие люди сейчас заметно «слепы» к растениям, которые не только поддерживают нас, но и разделяют наш мир.
Прочтите рассказы о растениях.
Что такое слепота растений?
Почти двадцать лет назад биологи диагностировали у людей эпидемию «растительной слепоты».Если мы не можем видеть растения, которые окружают нас каждый день — деревья, которые затеняют тротуары, лаванду, которая кормит пчел, комнатное растение, которое ползло по оконному стеклу, — мы также не сможем увидеть, что растения необходимы для нашей повседневной жизни.
Слепота растений делает нас нечувствительными как к частной жизни растений, так и к глубокой истории взаимоотношений растений и людей. Возможно, наиболее важным является то, что наша неспособность видеть растения локально делает нас слепыми к значительным последствиям деятельности человека для растительных сообществ во всем мире.
Что такое Гербария 3.0?
Herbaria 3.0 — это платформа для обмена историями о растениях и людях. Мы считаем, что эти истории могут привлечь наше внимание к взаимосвязанной природе взаимоотношений человека и растений. Обращение к этим отношениям помогает нам вспомнить растения и восстановить с ними связь, признавая ключевую роль растений в нашей жизни.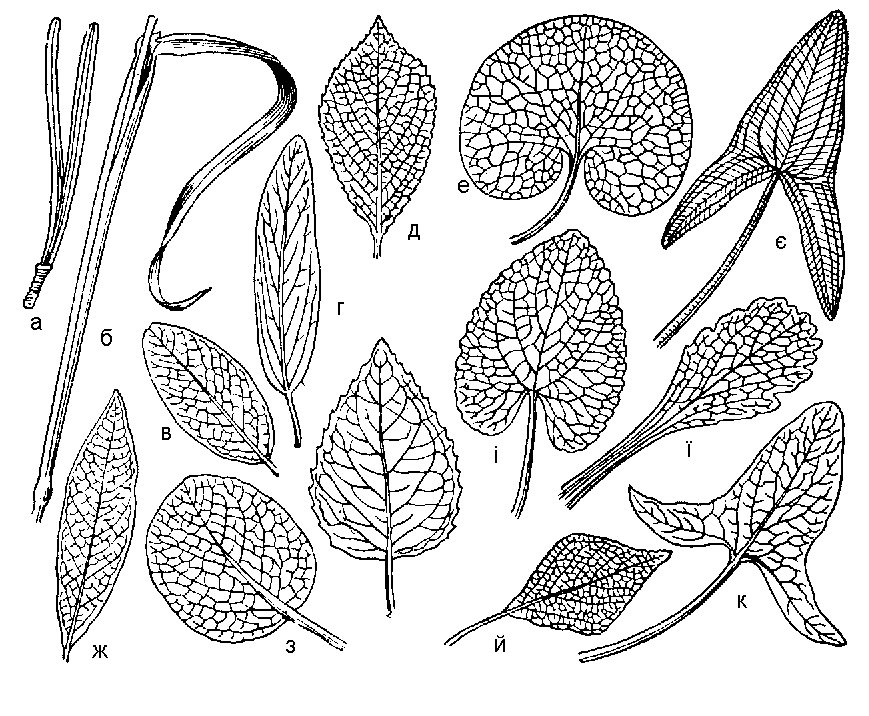
Что такое гербарии?
Zephyranthes treatiae, собранный Мэри Трит в 1877 году. Трит собрал этот вид в заповеднике Св.Река Джонс во Флориде. Профессор Гарварда Серено Уотсон назвала растение в ее честь в 1878 году.Гербарии — это коллекции консервированных образцов растений, состоящие из высушенных растений, помещенных на плотную белую бумагу, которые чаще всего готовятся коллекционерами растений или ботаниками для исследовательских целей. Первые гербарии были созданы в Италии в середине 1500-х годов как способ документирования лекарственных растений, и сегодня гербарии по-прежнему являются важным ресурсом для биологов-растений.
Лист с образцом гербария сохраняет, часто с любовью, отдельное растение.
Листы гербария, такие как левый, содержат важную информацию о растении: его строение, листья, корни, цветы; его название и классификационная информация; и процесс его сбора (кто собирал, где и когда). Если вы внимательно посмотрите на это изображение, полученное Мэри Трит, натуралистом XIX века и популярным писателем-натуралистом, вы увидите небольшой конверт, прикрепленный к листу; такие конверты могут содержать семена или другие важные части растения. Вы также увидите этикетки, которые отслеживают историю завода во времени, а также маркеры, используемые для сканирования и других цифровых целей.
Гербарии интересовали не только ботаников. На протяжении веков энтузиасты растений — часто обычные люди, которые в свободное время занимались сбором и сохранением растений — также создавали обширные гербарии, чтобы записывать и увековечивать растения, которые они находили во время своих прогулок. Гербарии записывают истории взаимодействия людей с растениями. Гербарий может быть личным документом, например, приведенным ниже примером из гербария поэтессы Эмили Дикинсон, или использоваться для совместного использования с друзьями и другими людьми.Но что верно для каждого примера, так это то, что каждый гербарный лист рассказывает историю. Каждый из них — живой объект, который меняет значение по мере своего движения во времени.
Каждый из них — живой объект, который меняет значение по мере своего движения во времени.
Вы увидите образцы гербарных образцов по всему сайту.
Почему Herbaria 3.0 ?
Оригинальный гербарный лист составляет «1.0» нашего проекта. Гербарии — мощные хроники исчезнувших и исчезающих миров. Они подробно описывают долгую и часто сложную историю взаимодействия растений и человека и, таким образом, служат эффективной метафорой для проекта этого места. Коллекция этих образцов в реальных и цифровых гербариях составляет «2.0». Вместе как образцы, так и их архивирование служат визуальным, тактильным и материальным хранилищем взаимодействий между растениями и людьми.Мы перезапустили концепцию гербариев в «Гербарии 3.0», предлагая место для сбора, обмена, картирования и архивирования современных встреч человека и растений, которые отражают глобальные перемещения растений и людей. Здесь мы стремимся раскрыть скрытые истории, вызвать новые повествования и создать светлое пятно надежды, точно так же, как растения показали устойчивость перед лицом перемен.
Скрытое вторжение и сокращение ниши, выявленное образцами гербариев в грибковом комплексе, вызывающем мучнистую росу дуба в Европе
Ajitomi A, Takushi T, Sato Y, Arasaki C, Ooshiro A (2020) Первое сообщение о мучнистой росе манго, вызванной Erysiphe quercicola в Японии.J Gen Plant Pathol 86: 316–321
CAS Google Scholar
Arnaud G, Foex E (1912) Sur la forme parfaite de l’Oïdium du chêne. C R Acad Sci 154: 124
Google Scholar
Базен Э, Мате-Юбер Х, Факон Б., Карлье Дж., Равинье В. (2014) Влияние системы спаривания на инвазивность: некоторая генетическая нагрузка может быть полезной при вторжении в новую среду. Биол Вторжения 16: 875–886
(2014) Влияние системы спаривания на инвазивность: некоторая генетическая нагрузка может быть полезной при вторжении в новую среду. Биол Вторжения 16: 875–886
Google Scholar
Behrens J (1921) Die Perithecien des Eiehenmeltaues в Германии.Z Pflanzenkrankh XXXI: 108–110
Google Scholar
Blackburn TM, Ewen JG (2017) Паразиты как движущие силы и пассажиры биологических инвазий, опосредованных человеком. EcoHealth 14: 61–73
PubMed Google Scholar
Blackwell M (2011) Грибы: 1, 2, 3. 5,1 миллиона видов? Am J Bot 98: 426–438
PubMed Google Scholar
Boddy L (2000) Межвидовые боевые взаимодействия между разлагающимися древесными базидиомицетами.FEMS Microbiol Ecol 31: 185–194
CAS PubMed Google Scholar
Boesewinkel HJ (1980) Идентификация плесени манго, Oidium mangiferae. Phytopathol Z 99: 126–130
Google Scholar
Boroń P, Lenart-Boroń A, Mullett M (2016) Распространение Dothistroma septosporum и типы его вязок в Польше. Форест Патол 46: 489–496
Google Scholar
Brasier CM, Buck KW (2001) Быстрые эволюционные изменения глобально вторгающегося грибкового патогена (болезнь голландского вяза).Биол Вторжения 3: 223–233
Google Scholar
Брюер М.Т., Кэдл-Дэвидсон Л., Кортеси П., Спану П.Д., Милгрум М.Г. (2011) Идентификация и структура локуса типа спаривания и разработка маркеров на основе ПЦР для типа спаривания в грибах мучнистой росы. Fungal Genet Biol 48: 704–713
CAS PubMed Google Scholar
Crooks JA (2005) Время задержки и экзотические виды: экология и управление биологическими инвазиями в замедленной съемке1. Экология 12: 316–329
Экология 12: 316–329
Google Scholar
Crous PW, Groenewald JZ, Slippers B, Wingfield MJ (2016) Глобальной продовольственной и волоконной безопасности угрожает текущая неэффективность идентификации грибов. Philos Trans R Soc London Ser B 371: 20160024
Google Scholar
DeBach P (1966) Принципы конкурентного вытеснения и сосуществования. Анну Преподобный Энтомол 11: 183–212
Google Scholar
Desprez-Loustau ML, Feau N, Mougou-Hamdane A, Dutech C (2011) Межвидовое и внутривидовое разнообразие мучнистой росы дуба в Европе: история коэволюции и адаптация к их хозяевам.Mycoscience 52: 165–173
Google Scholar
Desprez-Loustau ML, Hamelin F, Marçais B (2019) Экологическая и эволюционная траектория мучнистой росы дуба в Европе. В: Уилсон К., Фентон А., Томпкинс Д. (ред.) Экология болезней диких животных: связь теории с данными и применением. Cambridge University Press, Кембридж, стр. 429–457
Google Scholar
Desprez-Loustau ML, Robin C, Buee M, Courtecuisse R, Garbaye J, Suffert F et al (2007) Грибковое измерение биологических инвазий.Тенденции Ecol Evol 22: 472–480
PubMed Google Scholar
Desprez-Loustau ML, Massot M, Feau N, Fort T, de Vicente A, Torés JA, Ortuño DF (2017) Дальнейшее подтверждение сходства мучнистой росы дуба и манго и первое сообщение о Erysiphe quercicola и Erysiphe alphitoides на манго в континентальной Европе. Завод дис 101: 1086–1093
PubMed Google Scholar
Desprez-Loustau ML, Massot M, Toïgo M, Fort T et al.(2018) От листа к континенту: разномасштабное распространение инвазивного комплекса скрытых патогенов на дубе. Грибок Ecol 36: 39–50
Грибок Ecol 36: 39–50
Google Scholar
Данн А.М., Хэтчер М.Дж. (2015) Паразиты и биологические инвазии: параллели, взаимодействия и контроль. Тенденции Parasitol 31: 189–199
PubMed Google Scholar
Dutech C, Feau N, Lesur I, Ehrenmann F, Letellier T, Li B, Mouden C, Guichoux E, Desprez-Loustau ML, Gross A (2020) Простой и надежный метод выделения и проверки одиночного нуклеотида полиморфные маркеры из первого проекта генома Erysiphe alphitoides .Mycol Prog 19: 615–628
Google Scholar
Дворак М., Роткова Г., Ботелла Л. (2015) Обнаружение переносимого по воздуху инокулята Hymenoscyphus fraxineus и H. albidus во время сезонных колебаний, связанных с отсутствием апотециев. Леса 7: 1
Google Scholar
Edwards MC, Ayres PG (1982) Сезонные изменения устойчивости листьев Quercus petraea (скальный дуб) к Microsphaera alphitoides .Trans Br Mycol Soc 78: 569–571
Google Scholar
Enderle R, Stenlid J, Vasaitis R (2019) Обзор ясеня ( Fraxinus spp.) И болезни усмирения золы в Европе. CAB Rev 14: 1–12
Google Scholar
Feau N, Lauron-Moreau A, Piou D, Marçais B, Dutech C, Desprez-Loustau ML (2012) Разделение ниши генетических линий комплекса мучнистой росы дуба.Fungal Ecol 5: 154–162
Google Scholar
Фишер М.К., Гарнер Т.В., Уокер С.Ф. (2009) Глобальное появление Batrachochytrium dendrobatidis и хитридиомикоза амфибий в пространстве, времени и в организме хозяина. Annu Rev Microbiol 63: 291–310
CAS PubMed Google Scholar
Fisher MC, Henk DA, Briggs CJ, Brownstein JS, Madoff LC, McCraw SL, Gurr SJ (2012) Новые грибковые угрозы здоровью животных, растений и экосистем.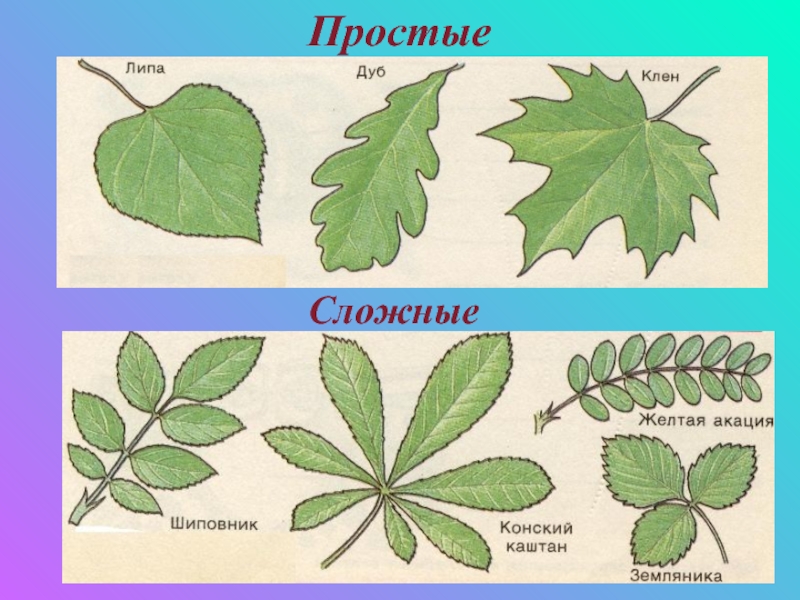 Nature 484: 186–194
Nature 484: 186–194
CAS PubMed Google Scholar
Фитт Б.Д., Хуанг Ю.Дж., ван ден Бош Ф., Вест Дж.С. (2006) Сосуществование родственных видов патогенов на пахотных культурах в пространстве и времени. Анну Рев Фитопатол 44: 163–182
CAS PubMed Google Scholar
Gao Y, Reitz SR (2017) Новые темы в нашем понимании смещения видов. Анну Рев Энтомол 62: 165–183
CAS PubMed Google Scholar
Gladieux P, Feurtey A, Hood ME, Snirc A, Clavel J, Dutech C et al (2015) Популяционная биология грибковых инвазий.Мол Экол 24: 1969–1986
CAS PubMed Google Scholar
Gorter GJMA (1984) Идентификация мучнистой росы дуба в Южной Африке. S Afr For J 129: 81–82
Google Scholar
Griffon E, Maublanc A (1912) Les Microsphaera des Chênes. Bull Soc Mycol F 28: 88–104
Google Scholar
Gross A, Han JG (2015) Hymenoscyphus fraxineus и два новых вида Hymenoscyphus , идентифицированных в Корее.Mycol Prog 14: 1–13
Google Scholar
Gross A, Holdenreider O, Pautasso M, Queloz V, Sieber TN (2014) Hymenoscyphus pseudoalbidus , возбудитель усыхания европейского ясеня. Мол Растение Патол 15: 109–117
Google Scholar
Гуревич Дж., Падилья Д.К. (2004) Являются ли инвазивные виды основной причиной исчезновения? Trends Ecol Evol 19: 470–474
PubMed Google Scholar
Hamelin FM, Bisson A, Desprez-Loustau ML, Fabre F, Mailleret L (2016) Дифференциация паразитов, разделяющих одно и то же растение-хозяин, по временной нише: мучнистая роса дуба в качестве примера. Экосфера 7: e01517
Экосфера 7: e01517
Google Scholar
Hariot P (1907) Note sur un oïdium du chêne. Bull Soc Mycol F 23: 157–159
Google Scholar
Hatcher MJ, Dick JT, Dunn AM (2012) Возникновение болезней и инвазии. Funct Ecol 26: 1275–1287
PubMed PubMed Central Google Scholar
Hawksworth DL, Lücking R (2017) Еще раз о разнообразии грибов: 2.От 2 до 3,8 миллиона видов. Царство грибов, стр. 79–95
Ioos R, Andrieux A, Marçais B, Frey P (2006) Генетическая характеристика естественного гибридного вида Phytophthora alni на основе ядерных и митохондриальных анализов ДНК. Fungal Gen Biol 43: 511–529
CAS Google Scholar
Исии Р., Хигаши М. (2001) Сосуществование, вызванное ограничением количества пыльцы у видов цветковых растений. Proc R Soc Lond Ser B 268: 579–585
CAS Google Scholar
Jolliffe PA (2000) Сменная серия.J Ecol 88: 371–385
Google Scholar
Kiss L (2008) Внутриклеточные микопаразиты в действии: взаимодействие между грибами мучнистой росы и Ampelomyces . В: Avery S, Stratford M, Van West P (eds) Стресс у дрожжей и мицелиальных грибов. Elsevier Academic Press, Кембридж, Массачусетс, США, стр. 37–52
Google Scholar
Кукол О., Ханацкова З., Дворжак М., Гаврдова Л. (2015) Не видны, но все еще присутствуют в Чехии: Hymenoscyphus albidus обнаружено с помощью ПЦР в реальном времени, но не с помощью интенсивного отбора проб.Mycol Prog 15: 6
Google Scholar
Ландольт Дж., Гросс А., Холденрейдер О., Паутассо М. (2016) Отмирание ясеня из-за Hymenoscyphus fraxineus : чему можно научиться из эволюционной экологии? Завод Патол 65: 1056–1070
Google Scholar
Limkaisang S, Cunnington JH, Wui LK, Salleh B, Sato Y, Divarangkoon R et al (2006) Молекулярно-филогенетический анализ показывает тесную взаимосвязь между грибами мучнистой росы на некоторых тропических деревьях и Erysiphe alphitoides , дубовым порошком. плесень.Mycoscience 47: 327–335
CAS Google Scholar
Linzmaier SM, Goebel LS, Ruland F, Jeschke JM (2018) Различия в поведении в сценарии чрезмерного вторжения: мраморный и остроконечный рак Ecosphere 9: e02385
Google Scholar
MacDougall AS, Gilbert B, Levine JM (2009) Инвазии растений и ниша. J Ecol 97: 609–615
Google Scholar
Маллон Калифорния, Ван Элсас Дж. Д., Саллес Дж. Ф. (2015) Микробные инвазии: процесс, закономерности и механизмы.Trends microbiol 23: 719–729
CAS PubMed Google Scholar
Marçais B, Kavkova M, Desprez-Loustau ML (2009) Фенотипические вариации в фенологии образования аскоспор между европейскими популяциями мучнистой росы дуба. Ann For Sci 66: 814
Google Scholar
Marçais B, Piou D, Dezette D, Desprez-Loustau ML (2017) Можно ли объяснить серьезность мучнистой росы дуба косвенным влиянием климата на состав патогенного комплекса Erysiphe ? Фитопатология 107: 570–579
PubMed Google Scholar
Mariette N, Mabon R, Corbière R, Boulard F, Glais I, Marquer B et al (2016) Фенотипические и генотипические изменения во французских популяциях Phytophthora infestans : являются ли инвазивные клоны наиболее агрессивными? Plant Pathol 65: 577–586
CAS Google Scholar
McKinney LV, Thomsen IM, Kjaer ED, Bengtsson SBK, Nielsen LR (2012) Быстрая инвазия агрессивного патогенного гриба ( Hymenoscyphus pseudoalbidus ) заменяет местный декомпозер ( Hymenoscyphus ). вымирание? Fungal Ecol 5 :: 663–669
Google Scholar
Mougou A, Dutech C, Desprez-Loustau ML (2008) Новое понимание личности и происхождения возбудителя мучнистой росы дуба в Европе.Для Pathol 38: 275–287
Google Scholar
Mougou-Hamdane A, Giresse X, Dutech C, Desprez-Loustau ML (2010) Пространственное распределение родословных грибов мучнистой росы дуба во Франции с использованием методов быстрого молекулярного обнаружения. Ann For Sci 67: 212–212
Google Scholar
Насир М., Могхал С.М., Мухтар Т., Аван М.З. (2014) Мучнистая роса манго: обзор экологии, биологии, эпидемиологии и управления.Crop Prot 64: 19–26
Google Scholar
Nishida S, Hashimoto K, Kanaoka MM, Takakura KI, Nishida T (2017) Различия в силе репродуктивного вмешательства от чужеродного родственника к местному виду в Taraxacum . J Plant Res 130: 125–134
PubMed Google Scholar
Паолетти М., Бак К.В., Бразье С.М. (2006) Селективное получение генов нового типа спаривания и вегетативной несовместимости посредством межвидового переноса генов в глобально вторгающемся эукариоте Ophiostoma novo-ulmi .Мол Экол 15: 249–262
CAS PubMed Google Scholar
Филибер А., Деспре-Лустау М.Л., Фабр Б., Фрей П., Халкетт Ф., Хассон С. и др. (2011) Прогнозирование успешной инвазии лесных патогенных грибов по признакам вида. J Appl Ecol 48: 1381–1390
Google Scholar
Rauschert ESJ, Shea K (2012) Инвазионное вмешательство из-за аналогичной межвидовой и внутривидовой конкуренции между захватчиками может повлиять на управление.Ecol Appl 22: 1413–1420
PubMed Google Scholar
Reitz SR, Trumble JT (2002) Конкурентное перемещение среди насекомых и паукообразных. Анну Рев Энтомол 47: 435–465
CAS PubMed Google Scholar
Ricciardi A, Blackburn TM, Carlton JT, Dick JT, Hulme PE, Iacarella JC и др. (2017) Наука о вторжении: обзор возникающих проблем и возможностей.Trends Ecol Evol 32: 464–474
PubMed Google Scholar
Rouxel M, Mestre P, Comont G, Lehman BL, Schilder A, Delmotte F (2013) Филогенетические и экспериментальные доказательства существования специализированных криптических видов в биотрофном оомицете. Новый Фитол 197: 251–263
PubMed Google Scholar
Рассел Дж. К., Сатаруддин Н. С., Херд А. Д. (2014) Чрезмерное вторжение функционально эквивалентных инвазивных видов.Экология 95: 2268–2276
PubMed Google Scholar
Scheele BC, Foster CN, Banks SC, Lindenmayer DB (2017) Сужение ниши у исчезающих видов: механизмы и последствия. Trends Ecol Evol 32: 346–355
PubMed Google Scholar
Шредер С., Телле С., Ник П., Тинес М. (2011) Тайное разнообразие Plasmopara viticola (Oomycota, Peronosporaceae) в Северной Америке.Org Divers Evol 11: 3–7
Google Scholar
Stroud JT, Giery ST, Outerbridge M, Feeley KJ (2019) Экологическое смещение характера изменяет результат приоритетных эффектов во время собрания сообщества. Экология 100: e02727
CAS PubMed Google Scholar
Takamatsu S, Braun U, Limkaisang S, Kom-Un S, Sato Y, Cunnington JH (2007) Филогения и таксономия мучнистой росы дуба Erysiphe alphitoides sensu lato .Mycol Res 111: 809–826
CAS PubMed Google Scholar
Тейлор Дж. В., Якобсон Д. Д., Крокен С., Касуга Т., Гейзер Д. М., Хиббетт Д. С., Фишер М. С. (2000) Филогенетическое распознавание видов и концепции видов у грибов. Fungal Genet Biol 31: 21–32
CAS PubMed Google Scholar
Torrend C (1909) L’Oidium du Chene en Portugal et a l’ile de Madere. Rev Sci Natur Collegio S Fiel Ser Bot VIII, Brotéria, стр. 103–113
Google Scholar
Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, Remm M, Rozen SG (2012) Primer3 — новые возможности и интерфейсы.Нуклеиновые кислоты res 40: e115 – e115
CAS PubMed PubMed Central Google Scholar
Viney R (1970) L’oïdium du Chêne: инцидент léger ou désastre. Ред. Для Fr
Viennot-Bourgin G (1968) Note sur les Erysiphacées. Bull Trim Soc Mycol France 84: 117–118
Google Scholar
Wu B, Hussain M, Zhang W, Stadler M, Liu X, Xiang M (2019) Текущее понимание разнообразия видов грибов и перспективы называния последовательностей ДНК грибов в окружающей среде.Микология 10: 127–140
PubMed PubMed Central Google Scholar
Xing Z, Zhang L, Wu S, Yi H, Gao Y, Lei Z (2017) Сравнение ниш между двумя инвазивными видами минеров и их паразитоидом Opius biroi : последствия для конкурентного вытеснения. Научный журнал 7: 1–6
Google Scholar
Ярвуд CE (1957) Мучнистая роса. Ботанический ред. 23: 235–301
Google Scholar
Йошида К., Бурбано Х.А., Краузе Дж., Тинес М., Вейгель Д., Камун С. (2014) Разработка гербариев для геномов патогенов растений: назад в будущее.Патогены PLoS 10: e1004028
PubMed PubMed Central Google Scholar
Zheng HD, Zhuang WY (2013) Hymenoscyphus albidoides sp. ноя и H. pseudoalbidus из Китая. Mycol Prog 13: 625–638
Google Scholar
Гетеробластная последовательность и развитие листьев у Leea guineensis по JSTOR
AbstractЛистья, зародившиеся в последовательных узлах гетеробластной серии у проростков Leea guineensis, становятся все более сложными в построении.Листья на первых четырех узлах простые, за ними следуют переходные формы между узлами 5 и 7. Эти переходные формы листьев сложны по своей природе, и промежуточные формы, такие как глубоко лопастные листья, не наблюдаются. В более высоких узлах сложная природа листьев становится все более сложной. Наиболее распространенная последовательность морфологий листа простая (узлы 1–4), от тройчатых до перистых (узлы 5–7), двупарчатых (узел 8) и трехлепестковых (узел 11). Морфология развития простых листьев в последовательности заметно отличалась от последующих сложных листьев.Однако ранние стадии развития сложных листьев, независимо от степени их сложности, были очень похожи. Следовательно, кажется, что существует поразительное различие в развитии между простыми листьями и сложными листьями в последовательности. Простые зачатки листьев становятся вогнутыми по центру и приобретают характерный складчатый вид, тогда как сложные листья имеют до трех порядков ветвления и имеют более сложную структуру. Однако сложные листья разного уровня сложности, по-видимому, обладают некоторыми общими особенностями развития.В максимуме сложную природу листа лучше всего описать в терминах семи листочков. Таким образом, сложность листа можно рассматривать как повторение базовой единицы. Увеличение сложности сложных листьев не влияет на скорость образования листьев.
Информация о журналеТекущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. С 1875 года Международный журнал наук о растениях (IJPS) представляет высококачественные, оригинальные, рецензированные исследования лабораторий по всему миру во всех областях науки о растениях.Охватываемые темы варьируются от генетики и геномики, биологии развития и клеточной биологии, биохимии и физиологии до морфологии и анатомии, систематики, эволюции, палеоботаники, взаимодействия растений и микробов и экологии. IJPS приветствует статьи, в которых представлены оценки и новые взгляды на актуальные области биологии растений.
Информация об издателеС момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета, University of Chicago Press взяла на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, способствующие образованию, развитию общественное понимание и обогащение культурной жизни.Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Измерение температуры с листьями: Структурированное исследование продолжительностью в семестр для студентов биологии растений | Американский учитель биологии
Недавние усилия по преобразованию и продвижению бакалавриата в области биологии постоянно подчеркивают преимущества перехода лабораторных упражнений от традиционных подходов «поваренных книг» к методике, основанной на запросах и исследованиях (NRC, 2003; AAAS, 2011).Такие усилия нацелены на то, чтобы научить студентов, как правильно практиковать науку, привлекая их к возможностям для использования мыслительных стратегий ученых, и подчеркивая открытый, ориентированный на открытия характер исследований, ведущих к неизвестным результатам (Chinn & Malhotra, 2002; Wallace et al., 2003; Weaver et al., 2008; Gormally et al., 2009; Kloser et al., 2011). Лаборатории, основанные на опросах, и опыт проведения курсовых исследований напрямую связаны с положительными результатами, включая повышение исследовательских навыков, научной грамотности, способности передавать знания, упорство в науке, повышение самоэффективности и более широкое вовлечение недостаточно представленных групп (Norris et al. al., 2003; Гормалли и др., 2009; Бангера и Браунелл, 2014 г .; Corwin et al., 2015). Несмотря на эти очевидные преимущества, существует множество препятствий на пути внедрения исследовательской педагогики на курсах бакалавриата по биологии, особенно проблемы, связанные с переводом студентов на открытую деятельность (Weaver et al., 2008). Этот переход часто чреват разочарованием, поскольку учащиеся сталкиваются с проблемами решения открытых вопросов без пошаговой схемы прогресса и успеха (Gormally et al., 2009). Без специального подхода к переходу между этими двумя различными лабораторными подходами студентам может просто не хватать знаний, опыта и абстрактных способностей мышления, чтобы полностью участвовать в аутентичных научных практиках (Lawson, 1980; Purser & Renner, 1983; Weaver et al., 2008). ; Gormally et al., 2009). Kloser et al. (2011) изложили несколько рекомендаций по эффективной исследовательской деятельности на основе курсов, которые могут обеспечить достаточную структуру для поддержки этого перехода. Они включают (1) ограниченный технический опыт, необходимый для сбора данных; (2) регулярные контрольные точки для обратной связи и исправления ошибок; (3) проекты, которые включают множество возможных переменных, позволяющих студентам формировать свои исследования в ограниченном объеме; и (4) оценки, которые имитируют распространение подлинных исследований, например, заключительный документ, отформатированный в соответствии со стандартами научного журнала.Эти рекомендации в сочетании с более структурированным исследовательским подходом, при котором ключевые параметры исследования определяются преподавателем, могут обеспечить баланс между исследованиями студентов и поддерживающими инструкциями и потенциально могут облегчить переход от традиционных лабораторий к аутентичным исследованиям (Weaver et al. ., 2008; Auchincloss et al., 2014).
Здесь я представляю программу исследований, разработанную для второго года курса биологии растений, которая объединяет ключевые элементы исследования и открытий, обеспечивая при этом структурированный подход к приобретению исследовательских навыков и научных компетенций.Этот учебный план был разработан для перехода студентов от «кулинарных» лабораторий, с которыми они сталкиваются на первом году обучения по общей биологии, к независимым исследовательским проектам, основанным на курсах, на третьем и четвертом году обучения, а также для устранения кажущейся нехватки хорошо разработанных исследовательских программ и основанные на исследованиях учебные программы для курсов биологии растений и ботаники. Учебная программа реализуется как семестр исследовательский проект, который включает в себя множество педагогических подходов с продемонстрированной эффективностью для повышения вовлеченности студентов, удержания и применения научной информации, включая обучение на открытом воздухе и полевые работы (Easton & Gilburn, 2012), экспериментальное обучение (Bauerle & Park, 2012) и проектной экологии (McCann & Miller, 2004; Purcell et al., 2007; Hall & Bauer-Armstrong, 2010), а также ключевые элементы аутентичного исследования и обучения на основе запросов. Учебная программа также включает сильный компонент количественного мышления, который включает статистическое моделирование и программирование на языке R, что соответствует цели курса, заключающейся в улучшении количественных знаний, навыков анализа данных и уверенности в применении количественных методов к биологическим данным.
Эта учебная программа была реализована в лабораторной части второго года курса биологии растений в Университете Джорджа Фокса, частном учебном заведении гуманитарных наук, в котором в настоящее время обучается 2389 студентов.Курс состоял из одного раздела из 17 специальностей биологии, каждый из которых завершил двух семестровый цикл общей биологии. Предварительное образование по темам биологии растений было минимальным для всех студентов. Учебный план лаборатории был реализован в виде трехчасового еженедельного лабораторного периода в течение 14 недель, при этом две субботние экскурсии заменили две еженедельные лабораторные встречи. Основная цель лабораторной программы — изучить, какие морфологические характеристики листьев наиболее сильно связаны со средней годовой температурой.Учебная программа охватывает девять результатов обучения, напрямую связанных с концепциями и компетенциями Vision и Change (AAAS, 2011; Таблица 1).
.