Вегетативная регуляция сердечной деятельности Текст научной статьи по специальности «Фундаментальная медицина»
УДК616.126.425-073.48
ВЕГЕТАТИВНАЯ РЕГУЛЯЦИЯ СЕРДЕЧНОЙ ДЕЯТЕЛЬНОСТИ
В.И. Шишко Кафедра факультетской терапии УО «Гродненский государственный медицинский университет»
В статье представлены современные представления о вегетативной регуляции сердечно-сосудистой системы, отражены вопросы иннервации и рефлекторной регуляции сердечной деятельности с позиций нормальной физиологии.
Ключевые слова: вегетативная нервная система, иннервация сердца, рефлекторная регуляция сердечной деятельности.
The article represents current views on vegetative regulation of cardiovascular system, and describes the aspects of innervation and reflex regulation of cardiac activity as viewed from physiology.
Key words: vegetative nervous system, cardiac innervation, reflex regulation of cardiac activity
Вегетативная нервная система (ВНС) — отдел нервной системы, регулирующий деятельность внутренних органов, желез внешней и внутренней секреции, кровеносных и лимфатических сосудов [10]. Первые сведения о структуре и функции ВНС принадлежат Галену (II век н.э.). J. Reil (1807) ввёл понятие «вегетативная нервная система», а J. Langley (1889) дал морфологическое описание ВНС, предложил деление её на симпатический и парасимпатический отделы, ввёл термин «автономная нервная система», учитывая способность последней самостоятельно осуществлять процессы регуляции деятельности внутренних органов. В настоящее время в русско, немецко-, франкоязычной литературе можно встретить термин вегетативная нервная система, а в англоязычной — автономная нервная система (АНС). Деятельность ВНС в основном непроизвольна и сознанием непосредственно не контролируется, направлена на поддержание постоянства внутренней среды и приспособление её к изменяющимся условиям внешней среды [8, 10].
Анатомия вегетативной нервной системы
С точки зрения иерархии управления, ВНС условно делят на 4 этажа (уровня). Первый этаж — интрамураль-ные сплетения, второй — паравертебральные и превер-тебральные ганглии, третий — центральные структуры симпатической нервной системы (СНС) и парасимпатической нервной системы (ПСНС). Последние представлены скоплениями преганглионарных нейронов в стволе мозга и спинном мозге. Четвёртый этаж включает высшие вегетативные центры (лимбико-ретикулярный комплекс — гиппокамп, грушевидная извилина, миндалевидный комплекс, перегородка, передние ядра таламуса, гипоталамус, ретикулярная формация, мозжечок, кора больших полушарий). Первые три этажа формируют сегментарный, а четвёртый — надсегментарный отделы ВНС. Кора головного мозга является высшим регуляторным центром интегративной деятельности, активируя как моторные, так и вегетативные центры. Лимбико-ретику-лярный комплекс и мозжечок отвечают за координацию вегетативных, поведенческих, эмоциональных, нейроэн-докринных реакций организма.
растает скорость проведения по специализированным проводящим тканям, увеличивается сократимость миокарда). Парасимпатический отдел ВНС оказывает трофот-ропное действие, способствуя восстановлению нарушенного во время активности организма гомеостаза, действует угнетающе на сердце (снижает ЧСС, атриовентри-кулярную проводимость и сократимость миокарда) [1, 4, 6, 9].
Ритм сердца определяется способностью специализированных клеток сердца спонтанно активироваться, так называемым свойством сердечного автоматизма. Автоматизм обеспечивает возникновение электрических импульсов в миокарде без участия нервной стимуляции. В нормальных условиях процессы спонтанной диастоли-ческой деполяризации, определяющие свойство автоматизма, наиболее быстро протекают в синоатриальном узле (СУ). Именно СУ узел задаёт ритм сердца, являясь водителем ритма 1 порядка. Обычная частота синусового импульсообразования — 60 — 100 импульсов в минуту, т.е. автоматизм СУ не является постоянной величиной, он может изменяться в связи с возможным смещением водителя ритма в пределах узла. В настоящее время ритм сердца рассматривается не только как показатель собственной функции ритмовождения СУ, но в большей степени как интегральный маркёр состояния множества систем, обеспечивающих гомеостаз организма. В норме основное модулирующее влияние на ритм сердца оказывает вегетативная нервная система (ВНС) [1, 6, 7].
Автоматизм обеспечивает возникновение электрических импульсов в миокарде без участия нервной стимуляции. В нормальных условиях процессы спонтанной диастоли-ческой деполяризации, определяющие свойство автоматизма, наиболее быстро протекают в синоатриальном узле (СУ). Именно СУ узел задаёт ритм сердца, являясь водителем ритма 1 порядка. Обычная частота синусового импульсообразования — 60 — 100 импульсов в минуту, т.е. автоматизм СУ не является постоянной величиной, он может изменяться в связи с возможным смещением водителя ритма в пределах узла. В настоящее время ритм сердца рассматривается не только как показатель собственной функции ритмовождения СУ, но в большей степени как интегральный маркёр состояния множества систем, обеспечивающих гомеостаз организма. В норме основное модулирующее влияние на ритм сердца оказывает вегетативная нервная система (ВНС) [1, 6, 7].
Иннервация сердца
Преганглионарные парасимпатические нервные волокна берут начало в продолговатом мозге, в клетках, которые находятся в дорсальном ядре блуждающего нерва (nucleus dorsalis n.
ЛЕКЦИИ И ОБЗОРЫ
дечных циклов.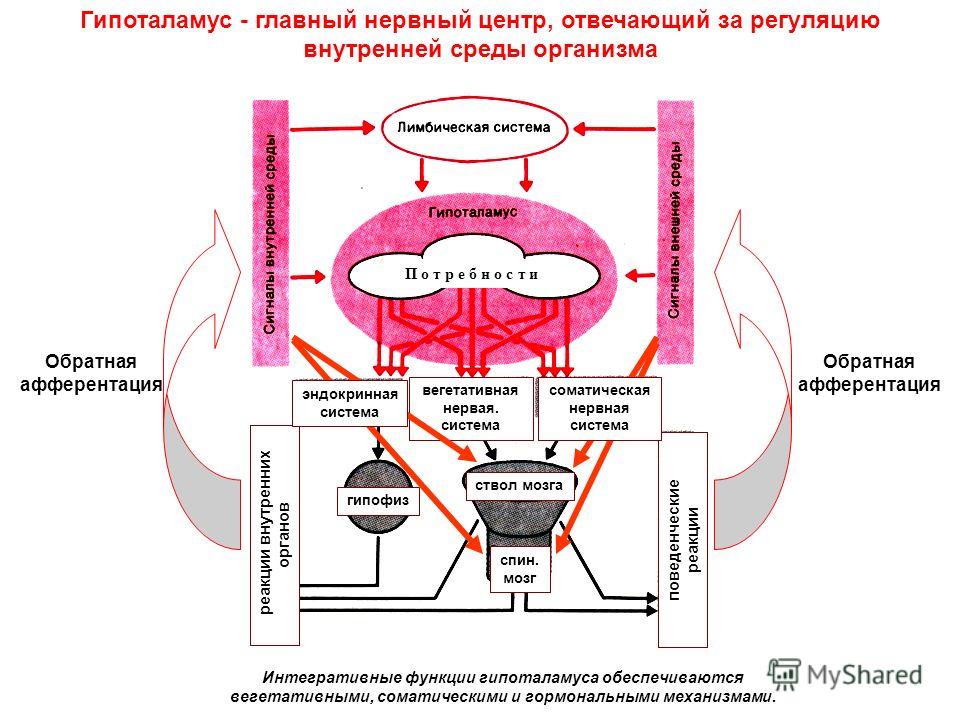 Однократная стимуляция блуждающего нерва или короткая серия импульсов оказывает влияние на ЧСС в течение последующих 15-20 с, с быстрым возвращением к контрольному уровню, благодаря быстрой деградации ацетилхолина в области СУ и АВС. Сочетание 2 характерных особенностей парасимпатической регуляции — короткого латентного периода и быстрого угасания ответной реакции, позволяет ей осуществлять быструю регуляцию и контроль за работой СУ и АВС практически при каждом сокращении [5, 7, 8].
Однократная стимуляция блуждающего нерва или короткая серия импульсов оказывает влияние на ЧСС в течение последующих 15-20 с, с быстрым возвращением к контрольному уровню, благодаря быстрой деградации ацетилхолина в области СУ и АВС. Сочетание 2 характерных особенностей парасимпатической регуляции — короткого латентного периода и быстрого угасания ответной реакции, позволяет ей осуществлять быструю регуляцию и контроль за работой СУ и АВС практически при каждом сокращении [5, 7, 8].
Волокна правого блуждающего нерва иннервируют преимущественно правое предсердие и особенно обильно СУ, а левого блуждающего нерва — АВС. В результате при раздражении правого блуждающего нерва более выражен отрицательный хронотропный эффект, а при стимуляции левого — отрицательный дромотропный. Парасимпатическая иннервация желудочков выражена слабо, в основном представлена в задненижней стенке левого желудочка. Поэтому при ишемии или инфаркте миокарда данной области отмечается брадикардия и гипотония, обусловленные возбуждением блуждающего нерва и описаны в литературе как рефлекс Бецольда-Яриша [4].
Преганглионарные симпатические волокна берут начало в интермедиалатеральных столбах 5-6 верхних грудных и 1-2 нижних шейных сегментах спинного мозга. Аксоны преганглионарных и постганглионарных нейронов образуют синапсы в трёх шейных и звёздчатом ганглиях. В средостении постганглионарные волокна симпатических и преганглионарные волокна парасимпатических нервов соединяются вместе, образуя сложное нервное сплетение смешанных эфферентных нервов, идущих к сердцу. Постганглионарные симпатические волокна достигают основания сердца в составе адвентиции крупных сосудов, где образуют обширное сплетение эпикарда. Затем они проходят сквозь миокард, вдоль коронарных сосудов. Нейромедиатором, выделяющимся из по-стганглионарных симпатических волокон, является норад-реналин, уровень которого одинаков как в СУ, так и в области правого предсердия [2, 7, 8, 9].
Повышение симпатической активности вызывает увеличение ЧСС, ускоряет диастолическую деполяризацию клеточных мембран, смещает водитель ритма к клеткам с самой высокой автоматической активностью. При стимуляции симпатических нервов ЧСС повышается медленно, латентный период реакции составляет 1-3 с, а установившийся уровень ЧСС достигается лишь через 3060 с от начала стимуляции. На скорость реакции влияет то, что медиатор вырабатывается довольно медленно нервными окончаниями, а воздействие на сердце осуществляется через относительно медленную систему вторичных мессинджеров — аденилатциклазу. После прекращения стимуляции хронотропный эффект исчезает постепенно. Скорость исчезновения эффекта стимуляции определяется снижением концентрации норадреналина в межклеточном пространстве, которая изменяется путём поглощения последнего нервными окончаниями, кардиомиоцитами и диффузией нейромедиатора в коронарный кровоток. Симпатические нервы практически равномерно распределены по всем отделам сердца, с максимальной иннервацией области правого предсердия. Симпатические нервы правой стороны преимущественно иннервируют переднюю поверхность желудочков и СУ, а левой — заднюю поверхность желудочков и АВС [7, 8].
При стимуляции симпатических нервов ЧСС повышается медленно, латентный период реакции составляет 1-3 с, а установившийся уровень ЧСС достигается лишь через 3060 с от начала стимуляции. На скорость реакции влияет то, что медиатор вырабатывается довольно медленно нервными окончаниями, а воздействие на сердце осуществляется через относительно медленную систему вторичных мессинджеров — аденилатциклазу. После прекращения стимуляции хронотропный эффект исчезает постепенно. Скорость исчезновения эффекта стимуляции определяется снижением концентрации норадреналина в межклеточном пространстве, которая изменяется путём поглощения последнего нервными окончаниями, кардиомиоцитами и диффузией нейромедиатора в коронарный кровоток. Симпатические нервы практически равномерно распределены по всем отделам сердца, с максимальной иннервацией области правого предсердия. Симпатические нервы правой стороны преимущественно иннервируют переднюю поверхность желудочков и СУ, а левой — заднюю поверхность желудочков и АВС [7, 8].
Афферентная иннервация сердца осуществляется в основном миелинизированными волокнами, идущими
в составе блуждающего нерва. Рецепторный аппарат в основном представлен механо- и барорецепторами, расположенными в правом предсердии, в устьях легочных и полых вен предсердий, желудочках, дуге аорты, синока-ротидном синусе [7]. По мнению большинства исследователей, регуляторные влияния ПСНС на СУ и АВС значительно превосходят влияния СНС.
Деятельность ВНС находится под влиянием центральной нервной системы (ЦНС) по механизму обратной связи. Обе системы тесно связаны между собой, а нервные центры на уровне ствола и полушарий головного мозга невозможно разделить морфологически. Самый верхний уровень взаимодействия осуществляется в сосудодвига-тельном центре, куда поступают и где обрабатываются афферентные сигналы из сердечно-сосудистой системы и где происходит регуляция эфферентной активности симпатической и парасимпатической нервной деятельности. Кроме интеграции на уровне ЦНС, важную роль играет также взаимодействие на уровне пре- и постси-наптических нервных окончаний, что подтверждено результатами анатомических и гистологических исследований. В последних исследованиях обнаружены особые клетки, содержащие большие запасы катехоламинов, на которых расположены синапсы, образованные терминальными окончаниями блуждающего нерва, что указывает на возможность прямого воздействия блуждающего нерва на адренергические рецепторы. Установлено, что часть внутрисердечных нейроцитов имеют положительную реакцию на моноаминоксидазу, что указывает на их роль в метаболизме норадреналина [7, 8].
Кроме интеграции на уровне ЦНС, важную роль играет также взаимодействие на уровне пре- и постси-наптических нервных окончаний, что подтверждено результатами анатомических и гистологических исследований. В последних исследованиях обнаружены особые клетки, содержащие большие запасы катехоламинов, на которых расположены синапсы, образованные терминальными окончаниями блуждающего нерва, что указывает на возможность прямого воздействия блуждающего нерва на адренергические рецепторы. Установлено, что часть внутрисердечных нейроцитов имеют положительную реакцию на моноаминоксидазу, что указывает на их роль в метаболизме норадреналина [7, 8].
Несмотря на разнонаправленное в целом действие СНС и ПСНС, при одновременной активации обоих отделов ВНС их эффекты не складываются простым алгебраическим способом и взаимодействие нельзя выразить линейной зависимостью. В литературе описано несколько типов взаимодействия отделов ВНС [1, 4, 8, 9, 12]. Согласно принципу «акцентированного антагонизма», ин-гибирующий эффект данного уровня парасимпатической активности тем сильнее, чем выше уровень симпатической активности, и наоборот. С другой стороны, при достижении определённого результата снижения активности в одном отделе ВНС происходит повышение активности другого отдела по принципу «функциональной синергии». При исследовании вегетативной реактивности необходимо учитывать «закон исходного уровня», согласно которому чем выше исходный уровень, тем в более деятельном и напряженном состоянии находится система, тем меньший ответ возможен при действии возмущающих стимулов.
С другой стороны, при достижении определённого результата снижения активности в одном отделе ВНС происходит повышение активности другого отдела по принципу «функциональной синергии». При исследовании вегетативной реактивности необходимо учитывать «закон исходного уровня», согласно которому чем выше исходный уровень, тем в более деятельном и напряженном состоянии находится система, тем меньший ответ возможен при действии возмущающих стимулов.
Состояние отделов ВНС претерпевает значительные изменения на протяжении жизни человека. В младенческом возрасте отмечается значительное преобладание симпатических нервных влияний при функциональной и морфологической незрелости обоих звеньев ВНС. Развитие симпатического и парасимпатического отделов ВНС после рождения происходит интенсивно, и к моменту полового созревания плотность расположения нервных сплетений в различных отделах сердца достигает наивысших показателей. При этом у лиц молодого возраста отмечается доминирование парасимпатических влияний, проявляющихся в исходной ваготонии в состоянии покоя. Начиная с 4-го десятилетия жизни, начинаются инволютивные изменения в аппарате симпатической иннервации, при сохранении плотности холинерги-ческих нервных сплетений. Процессы десимпатизации приводят к снижению симпатической активности и падению плотности распределения нервных сплетений на кар-
Начиная с 4-го десятилетия жизни, начинаются инволютивные изменения в аппарате симпатической иннервации, при сохранении плотности холинерги-ческих нервных сплетений. Процессы десимпатизации приводят к снижению симпатической активности и падению плотности распределения нервных сплетений на кар-
диомиоцитах, гладкомышечных клетках, способствуя гетерогенности потенциалзависимых свойств мембраны в клетках проводящей системы, рабочем миокарде, стенках сосудов, гиперчувствительности рецепторного аппарата к катехоламинам и могут служить основой аритмий, в том числе и фатальных. Имеются также и половые различия в состоянии вегетативного нервного тонуса. Так, у женщин молодого и среднего возраста (до 55 лет) отмечена более низкая активность симпатической нервной системы, чем у мужчин аналогичного возраста [11]. Таким образом, вегетативная иннервация различных отделов сердца неоднородна и несимметрична, имеет возрастные и половые различия. Согласованная работа сердца является результатом динамического взаимодействия отделов ВНС между собой [5, 6, 7, 9].
Рефлекторная регуляция сердечной деятельности
Артериальный барорецепторный рефлекс является ключевым механизмом краткосрочной регуляции артериального давления (АД). Оптимальный уровень системного артериального давления является одним из наиболее важных факторов, необходимых для адекватной работы сердечно-сосудистой системы. Афферентные импульсы от барорецепторов каротидных синусов и дуги аорты по ветвям языкоглоточного нерва (IX пара) и блуждающего нерва (Х пара) поступают к кардиоингибитор-ному и сосудодвигательному центру продолговатого мозга и другим отделам ЦНС. Эфферентное плечо баро-рецепторного рефлекса образуется симпатическими и парасимпатическими нервами. Импульсация от бароре-цепторов повышается на увеличении абсолютной величины растяжения и скорости изменения растяжения рецепторов. Повышение частоты импульсации от бароре-цепторов оказывает тормозящее влияние на симпатические центры и возбуждающее на парасимпатические, что приводит к снижению вазомоторного тонуса в резистив-ных и емкостных сосудах, уменьшению частоты и силы сердечных сокращений. Если среднее АД резко снижается, тонус блуждающего нерва практически исчезает, а рефлекторная регуляция осуществляется исключительно за счёт изменений эфферентной симпатической активности. При этом повышается общее периферическое сопротивление сосудов, увеличивается частота и сила сердечных сокращений, направленных на восстановление исходного уровня АД. И наоборот, если АД резко повышается, симпатический тонус полностью угнетается, а градация рефлекторной регуляции происходит только благодаря изменениям эфферентной регуляции вагу-са [3].
Если среднее АД резко снижается, тонус блуждающего нерва практически исчезает, а рефлекторная регуляция осуществляется исключительно за счёт изменений эфферентной симпатической активности. При этом повышается общее периферическое сопротивление сосудов, увеличивается частота и сила сердечных сокращений, направленных на восстановление исходного уровня АД. И наоборот, если АД резко повышается, симпатический тонус полностью угнетается, а градация рефлекторной регуляции происходит только благодаря изменениям эфферентной регуляции вагу-са [3].
Повышение давления в желудочках вызывает раздражение субэндокардиальных рецепторов растяжения и активацию парасимпатического кардиоингибиторного центра, что приводит к рефлекторной брадикардии и ва-зодилатации [7, 10].
Рефлекс Бейбриджа характеризуется повышением симпатического тонуса с повышением ЧСС в ответ на увеличение внутрисосудистого объема крови и повышение давления в крупных венах и правом предсердии. При этом происходит рост ЧСС, несмотря на сопутствующий подъём АД. В реальной жизни рефлекс Бейбрид-
При этом происходит рост ЧСС, несмотря на сопутствующий подъём АД. В реальной жизни рефлекс Бейбрид-
жа преобладает над артериальным барорецепторным рефлексом в случае увеличения объёма циркулирующей крови. Исходно и при уменьшении объёма циркулирующей крови барорецепторный рефлекс преобладает над рефлексом Бейбриджа [4, 10].
Ряд факторов, участвующих в поддержании гомеос-таза организма, влияет на рефлекторную регуляцию сердечной деятельности, при отсутствии значимых изменений активности ВНС. К ним относятся хеморецепторный рефлекс, изменения уровня электролитов крови (калия, кальция). На частоту сердечных сокращений оказывают влияние также фазы дыхания: вдох вызывает угнетение блуждающего нерва и ускорение ритма, выдох — раздражение блуждающего нерва и замедление сердечной деятельности [7].
Таким образом, в обеспечении вегетативного гомео-стаза участвует большое количество разнообразных ре-гуляторных механизмов. По мнению большинства исследователей, ритм сердца является не только показателем функции СУ, но и интегральным маркёром состояния множества систем, обеспечивающих гомеостаз организма, с основным модулирующим влиянием ВНС [1, 3, 9, 12]. Попытка выделить и количественно оценить влияние на ритм сердца каждого из звеньев — центрального, вегетативного, гуморального, рефлекторного — несомненно, является актуальной задачей в кардиологической практике, так как её решение позволит разработать дифференциально-диагностические критерии сердечно-сосудистой патологии на основании простой и доступной оценки состояния ритма сердца
По мнению большинства исследователей, ритм сердца является не только показателем функции СУ, но и интегральным маркёром состояния множества систем, обеспечивающих гомеостаз организма, с основным модулирующим влиянием ВНС [1, 3, 9, 12]. Попытка выделить и количественно оценить влияние на ритм сердца каждого из звеньев — центрального, вегетативного, гуморального, рефлекторного — несомненно, является актуальной задачей в кардиологической практике, так как её решение позволит разработать дифференциально-диагностические критерии сердечно-сосудистой патологии на основании простой и доступной оценки состояния ритма сердца
Литература
1 Баевский, P.M. Математический анализ изменений сердечного ритма при стрессе / Р.М. Баевский, О.И. Кириллов, С.З. Клецкин. — М.: Наука, 1984. — 220 с.
2 Вейн, А.М. Вегетативные расстройства / А.М. Вейн. — М.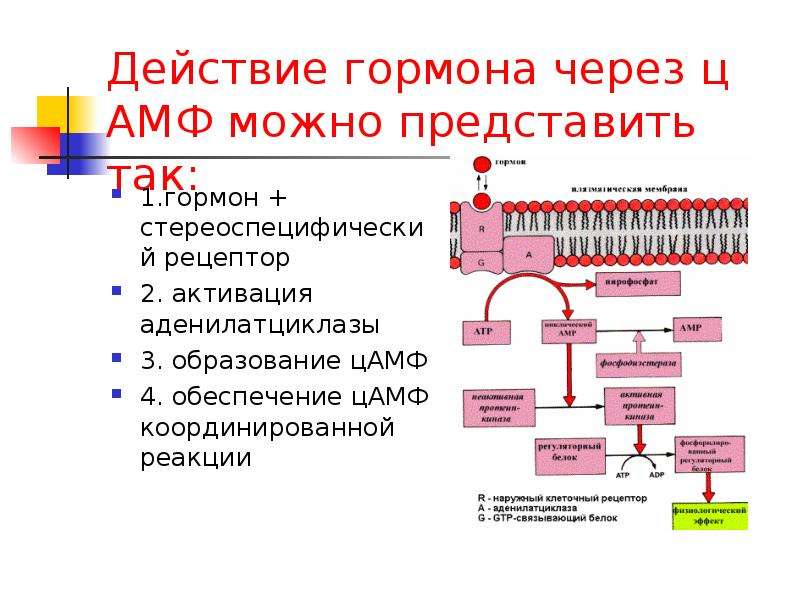 : Медицина, 1998. — 740 с.
: Медицина, 1998. — 740 с.
3 Михайлов, В.М. Вариабельность ритма сердца: опыт практического применения / В.М. Михайлов. — Иваново: Иван. гос. мед. академия, 2002. — 290 с.
4 Михайлов, С.С. Клиническая анатомия сердца / С.С. Михайлов. — М.: Медицина, 1987. — 287 с.
5 Морман, Д. Физиология сердечно-сосудистой системы / Д. Морман, Л. Хеллер. — Питер., 2000. 363 с.
6 Патофизиология заболеваний сердечно-сосудистой системы / под ред. Л. Лили; пер. с англ. — М.: БИНОМ. Лаборатория знаний, 2003. — 598 с.
7 Рябыкина, Г.В. Анализ вариабельности ритма сердца / Г.В. Рябыкина, А.В. Соболев // Кардиология. — 1996. — №10. — С. 8797.
8 Физиология и патофизиология сердца / под ред. Н.Спере-лакиса. — М.: Медицина, 1990. — Т. 2. — 624 с.
— М.: Медицина, 1990. — Т. 2. — 624 с.
9 Физиология человека / Н.А. Агаджанян [и др.]; СПб.: Со-тис,1998.- 527с.
I 0 Физиология человека: в 3 т.: пер. с англ.; под ред. Р.М. Шмидта и Г. Тевса. — М.: Мир, 1996.
II Швалёв, В.Н. Феномен ранней возрастной инволюции симпатического отдела вегетативной нервной системы / В.Н. Швалев, Н.Ф. Тарский // Кардиология. — 2001. — № 2. — С. 10-14.
12 Banister, R. Autonomic Failure: a Textbook of Clinical Disorders of the Autonomic Nervous System / R. Banister. — Oxford, 1983.
Поступила 10.03.09
симптомы, как лечить, чем опасно?
Все наши внутренние органы работают под воздействием сигналов со стороны вегетативной нервной системы. В случае поступления неправильных или нерегулярных сигналов работа внутренних органов нарушается, снижаются защитные функции организма. Это приводит к ухудшению общего самочувствия и дает симптоматику, сходную с мигренью, инфарктом миокарда, остеохондрозом и рядом других патологий.
В случае поступления неправильных или нерегулярных сигналов работа внутренних органов нарушается, снижаются защитные функции организма. Это приводит к ухудшению общего самочувствия и дает симптоматику, сходную с мигренью, инфарктом миокарда, остеохондрозом и рядом других патологий.
Подобные состояния могут развиться на фоне постоянных стрессов или быть ими спровоцированы, возникнув по какой-либо другой причине. Зачастую вегетативные нервные расстройства являются частью функциональных или органических поражений нервной системы в целом.
Типы вегетативной дисфункции
Часто дисфункцию вегетативной нервной системы путают с вегетососудистой дистонией (ВСД). Однако ВСД — лишь одно из проявлений различных заболеваний, в том числе и нервных.
- Соматоформное расстройство вегетативной нервной системы. Это разновидность невроза, проявляющаяся в виде симптомов различных хронических заболеваний, которых на самом деле у пациента нет.
 Возможны периодические приступы, клинически сходные с паническими атаками. Также часты психогенный кашель и одышка, нарушения пищеварения, головокружения и т.д. Этот вид вегетативной дисфункции встречается наиболее часто, как правило, обусловлен хроническим стрессом и поддается лечению лучше всего.
Возможны периодические приступы, клинически сходные с паническими атаками. Также часты психогенный кашель и одышка, нарушения пищеварения, головокружения и т.д. Этот вид вегетативной дисфункции встречается наиболее часто, как правило, обусловлен хроническим стрессом и поддается лечению лучше всего. - Поражение подкорковых структур. Возникает при различных травмах головного мозга, резидуальной патологии ЦНС. Для этого заболевания характерны полуобморочные состояния, снижение артериального давления, обильное мочеиспускание и диарея. По окончании приступа больной ощущает слабость и вялость.
- Постоянное раздражение периферических вегетативных структур. Может возникнуть при предменструальном синдроме, мочекаменной болезни, шейной дорсопатии с поражением симпатического шейного сплетения.
 Возможны периодические приступы, клинически сходные с паническими атаками. Также часты психогенный кашель и одышка, нарушения пищеварения, головокружения и т.д. Этот вид вегетативной дисфункции встречается наиболее часто, как правило, обусловлен хроническим стрессом и поддается лечению лучше всего.
Возможны периодические приступы, клинически сходные с паническими атаками. Также часты психогенный кашель и одышка, нарушения пищеварения, головокружения и т.д. Этот вид вегетативной дисфункции встречается наиболее часто, как правило, обусловлен хроническим стрессом и поддается лечению лучше всего.Вегетативная дисфункция любого типа является проявлением серьезных проблем в организме, поэтому при первых же симптомах обязательно следует выявить ее причину и начать лечение.
Причины и симптомы расстройства ВНС
Вегетативными нарушениями в современном мире страдает около 70% взрослых и 25% детей. Такая статистика никого не удивит, если взглянуть на список возможных причин вегетативной дисфункции:
- хронический стресс;
- наследственность;
- гормональные сбои или возрастные гормональные изменения в организме;
- малоподвижный образ жизни;
- неправильное питание;
- злоупотребление алкоголем и табаком;
- травмы, ранения, хирургические операции, нарушающие целостности нервных связей;
- интоксикация нервной и кровеносной систем в результате воспалительных процессов;
- длительный прием сильнодействующих лекарств, самолечение;
- аллергические заболевания.
Кстати
Обычно проявления вегетативной дисфункции дают о себе знать в возрасте 20–30 лет, женщинам ставят подобный диагноз в 2–3 раза чаще, чем мужчинам. Однако это не говорит о том, что мужчины заболевают реже. Просто женщины больше склонны обращаться за медицинской помощью. Мужчины же, во-первых, предпочитают не жаловаться, а во-вторых, обычно задействуют «мужские» методы решения проблем — например, алкоголь.
Симптомы расстройства вегетативной нервной системы могут быть следующими:
- Сердечно-сосудистый синдром. Нарушение сердечного ритма, скачки артериального давления, нарушение периферического кровообращения, внезапный дискомфорт в области сердца.
- Гипервентиляционный синдром. Учащенное дыхание, ощущение недостатка воздуха, головокружение, нарушение чувствительности в конечностях, мышечные спазмы.
- Синдром раздраженного кишечника. Спазмы и ноющие боли внизу живота, частые позывы к дефекации, повышенное газообразование, диарея.
- Расстройства со стороны пищеварительного тракта. Тошнота и рвота, затрудненное глотание («комок в горле»), боль и дискомфорт под ложечкой, нарушение аппетита.
- Повышенная потливость. Как правило, в области ладоней и подошв.
- Цисталгия. Частое болезненное мочеиспускание, не связанное с болезнями мочеполового тракта.
- Сексуальные расстройства. Нарушения эрекции и эякуляции у мужчин, вагинизм и аноргазмия у женщин, снижение либидо.
- Нарушение терморегуляции. Ознобы, повышение температуры.
К части симптомов больные способны привыкнуть или списывать их на другие заболевания. Однако со временем количество проявлений вегетативной дисфункции растет, они учащаются, и игнорировать их уже невозможно.
К кому обратиться при расстройстве вегетативной нервной системы и как его лечить?
Проблемами ВНС занимается врач-невролог, который после ряда диагностических процедур уточняет тип расстройства и назначает необходимую терапию. Главное условие успешного лечения, вне зависимости от причин заболевания, — снижение тревожности и борьба со стрессом.
Амедикаментозные методики
Не стоит рассчитывать на то, что проблема решится приемом лекарств трижды в день. От пациента потребуется заинтересованность в лечении и работа над собой.
- Изменение образа жизни. Прежде всего следует исключить нервные перегрузки на работе и дома, наладить режим труда и отдыха. Противопоказан ненормированный рабочий день, работа в авральных режимах. Необходимо пересмотреть рацион питания, включить в него больше натуральной белковой пищи, овощей и фруктов. Необходимо чаще бывать на свежем воздухе, показаны пешие прогулки, йога, занятия в бассейне.
- Физиотерапия. Стоунтерапия, массаж, акупунктура, электрофорез, грязевые ванны направлены на релаксацию, снятие мышечных зажимов, улучшение кровообращения и оттока лимфы. Все это благотворно влияет на общее состояние пациентов, снимает стрессовое напряжение.
- Работа с психологом. Зачастую заболевание развивается на фоне особенностей психологии. Самый распространенный пример — трудоголизм. Увлеченный своей деятельностью человек, привыкший работать в стрессовом режиме, не замечает симптомов болезни, считает их недостойными внимания и резко отрицает необходимость не только в санаторно-курортном лечении, но и в элементарном ежедневном отдыхе. Проблему может решить только работа с психологом.
Фармакотерапия
Лекарственные средства назначаются с несколькими целями. Необходимо нормализовать обмен веществ, укрепить иммунитет, восстановить работу сердечно-сосудистой системы, снять тревожные и депрессивные состояния.
- Фитопрепараты, витамины, БАДы. Успокоительные чаи и сборы, настойки, экстракты пришли к нам из народной медицины. Наиболее популярными являются фитопрепараты на основе ромашки, валерианы, пустырника. Витаминные комплексы для нервной системы должны включать в себя витамины С и Е, а также витамины группы В. Прием биоактивных добавок к пище может быть направлен как на достижение успокоительного эффекта, так и на укрепление иммунитета. Для этого подходят препараты эхинацеи.
- Лекарства для нормализации работы сердечно-сосудистой системы. Наиболее часто назначаются «Корвалол» и «Валокордин», оказывающее стабилизирующее влияние не только на работу сердца при некоторых видах нарушения ритма, но и обладающие седативным эффектом.
- Антидепрессанты и седативные препараты. Назначаются комплексно или по отдельности, в зависимости от клинической картины. Среди современных безрецептурных анксиолитических препаратов (противотревожных) стоит выделить «Афобазол». Он, в отличие от сильнодействующих рецептурных средств, не вызывает привыкания, сонливости, снижения внимания.
«Афобазол» успешно применяется при расстройствах адаптации (стрессе) и тревожных расстройствах различного генеза. В том числе возникших на фоне соматических заболеваний. «Афобазол» помогает снизить или устранить такие симптомы, как мышечное напряжение, сердечно-сосудистые симптомы, приступы удушья, чувство «комка в горле», стрессовые расстройства ЖКТ, сухость во рту, потливость, ощущение приливов, головокружение, головные боли, напряжение, кожный зуд. «Афобазол» уменьшает проявления вегетативной дисфункции у 89% пациентов. Препарат обычно назначается курсом с продолжительностью 2–4 недели. Особенность этого лекарственного средства состоит в том, что его действие направлено на восстановление нормальной работы нервной системы и защиту организма от стресса в различных его проявлениях.
При расстройстве вегетативной нервной системы успех лечения невозможен без самодисциплины пациента. Соблюдая график лечения и не пропуская назначенные процедуры, вы можете быть уверены в скором облегчении своего состояния.
Вегетативная регуляция ритма сердца и показатели сигнал-усредненной ЭКГ у больных гипертонической болезнью | Рузов
1. Мазур Н.А. Внезапная смерть Клин мед 1988, 11 28-35
2. Татарченко И.П., Позднякова Н.В., Морозова О.И., Шевырев В.А. Прогноз больных ишемической болезнью сердца — клинико-инструментальные аспекты Пенза ЭЛМА, 2002, с 21-3
3. Потапова Н.П., Иванов Г.Г., Буланова Н.А. Современные неинвазивные методы оценки и прогнозирования развития потенциально опасных и угрожающих жизни аритмий состояние проблемы и перспективы развития Кардиология 1997, 2 70-5
4. Легконогое A.B. Результаты и перспективы изучения поздних потенциалов желудочков Кардиология 1997, 10 57-65
5. Чирейкин Л.В., Быстров Я.Б., Шубин Ю.В. Поздние потенциалы желудочков в современной диагностике и прогнозе течения заболеваний сердца Вестн аритмол 1999, 13 61-74
6. Palatim Р., Julius S. Heart rate and cardiovascular risk Journal Hypertension 1990, 81 850-9
7. Barron H.V., Lesh M.D. Autonomic nervous system and sudden cardiac death J Am Cardiol 1996, 27 1053-60
8. Vester E.G., Kuhls S., Penngs С. Ventricular late potentials m hypertensive petients with and without left ventricular hypertrophy PASE 1993, 16 913-26
9. Wiseman M.N., Levy M.N. Electrophysiologic mechanisms for ventricular arrhythmic m left ventricular dysfunction electrolytes, catecholamines and drug J Clm Pharmacol 1991,32 1053-60
10. Guidelines Subcommitee 1999 World Health Organization Interna tional Sosiety of Hypertension guidelines for the management of hypertension Journal of Hypertension 1999, 17 151-83
11. Simson M.B. Use of signals m the terminal QRS complex to identify patients with ventricular tachycardia after myocardial infarction Circulation 1981, 64 235-42
12. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology Heart rate variability Standards of measurement, physiological interpretation and clinical use Circulation 1996, 93 1043-56
13. Шляхто Е.В., Конради А.О. Причины и последствия активации симпатической нервной системы при артериальной гипертензии Артериальная гипертензия 2003, 9 (3) 81 -8
14. Letу M.N., Schwarts P.J. et al. Vagal control of the heart Experimental basis and clinical implications Arrnonk Future 1994, 78-84
15. Lown B., Vemer R.L. Neural activity and ventricular fibrillation Engl J Med 1976, 249 1165-70
16. Беленков Ю.Н. Ремоделирование левого желудочка комплексный подход Сердеч недостаточность 2002, 3 (4) 161-3
Статья «Вегетативная дистония у детей»
Все органы и системы организма находятся под регулирующим воздействием нервной системы, состоящим из центрального отдела (головной мозг) и вегетативного отдела. Вегетативная нервная система при взаимодействии с корой головного мозга и эндокринной системой (щитовидная железа, гипофиз, надпочечники) принимает непосредственное участие в адаптации организма, регулирует реакции на любые стрессорные воздействия. Вегетативная нервная система состоит из 2 отделов: симпатический и парасимпатический. Симпатический отдел включаются в ситуациях, требующих напряженной психической и физической деятельности. Парасимпатический отдел проявляет свою основную функцию вне периода наряженной активности организма, преимущественно в период «отдыха».
«Синдром вегетативной дистонии» — это состояние, которое характеризуется нарушениями вегетативной (нервной) регуляции работы внутренних органов: сердечно-сосудистой системы, желудочно-кишечного тракта, органов дыхания, желез внутренней секреции. Изменения со стороны всех этих систем носят функциональный, т.е. обратимый характер и не представляют угрозы для жизни и здоровья ребенка. Причиной развития вегетативной дистонии являются наследственно — конституциональные факторы.
Дети с дистонией могут предъявлять самые разнообразные жалобы
Самые частые из них — головные боли, боли в области сердца и живота. Как правило, такие дети плохо переносят транспорт, душные помещения, могут быть головокружения и даже кратковременные потери сознания (обмороки). Нередко отмечается нестабильное артериальное давление, повышенная утомляемость, беспокойный сон, перемена настроения. Часто бывают внезапные ощущения проколов в левой половине грудной клетки, особенно на вдохе, сопровождающиеся чувством затрудненного дыхания, сердцебиения, бледностью.
Со стороны органов дыхания могут отмечаться приступы внезапной одышки, без видимых причин появляются глубокие «вздохи», так называемый «дыхательный невроз», приступы невротического, спазматического кашля.
При дистонии могут появляться боли и неприятные ощущения в области мышц ног, чувства онемения, преимущественно по вечерам, перед засыпанием — симптом «беспокойных ног».
Существует 2 варианта вегетативной дистонии:
- по ваготоническому типу
- по симпатикотоническому типу
При ваготоническом типе дети жалуются на частые головные боли, связанные с понижением артериального давления, боли в области сердца, редкий пульс, обморочные состояния в душных помещениях, чрезмерную потливость, зябкость, длительные сохранения субфебрильной (до 37.5) температуры после перенесенных инфекций. Часто бывают немотивированные боли в животе, тошнота, метеоризм. Такие дети быстро устают, у них повышенная тревожность, сонливость, утомляемость, склонность к депрессиям.
При симпатикотоническом типе дистонии у детей наоборот отмечаются сердцебиения, ускоренный пульс, склонность к повышенному давлению.
Верхние пределы артериального давления у детей:
- 7-9 лет 125/75
- 10-13 лет 130/80
- 14-17 лет 135/85
У таких детей часто бывают проявления «термоневроза» — повышение температуры тела на фоне стресса, колебания в течение суток, субфебрильная температура при отсутствии признаков заболевания. Такие дети обычно худые, несмотря на нормальный аппетит.
Рекомендации для родителей
В основе лечения вегетативной дистонии лежат немедикаментозные методы лечения.
Они включают в себя: правильную организацию труда и отдыха, соблюдение распорядка дня, занятия физической культурой, рациональное питание, водолечение и бальнеотерапию, физиотерапию, психотерапию. Необходимо устранить гиподинамию, так как физическая активность повышает тонус сердечно-сосудистой системы, улучшает обменные процессы и микроциркуляцию.
Прогулки на свежем воздухе должны быть не менее 2 часов в день. Освобождение от занятий физкультуры в школе не требуется.
Рекомендуемые занятия спортом: плавание, коньки, лыжи, игровые виды спорта, велосипед, легкая атлетика.
Не рекомендуется: бокс, все виды борьбы, тяжелая атлетика.
Ночной сон должен составлять не менее 8-10 часов, так как сон является основным регулятором циркадных биоритмов организма. Занятия за компьютером должны составлять не более 40 — 60 минут в день в зависимости от возраста ребенка.
Питание детей должно быть рациональным, с достаточным количеством минеральных веществ и витаминов. Нужно избегать избытка мучных продуктов, копченостей, животных жиров. При склонности к повышению АД необходимо ограничение соли, жидкости, высококалорийных продуктов. При ваготонической дистонии можно включать в рацион кофе, шоколад, крепкий чай, солености.
Очень эффективны водные процедуры:
- плавание
- лечебные ванны (солено — хвойные, кислородные, жемчужные)
- души (циркулярный, контрастный, подводный, душ Шарко, игольчатый)
В лечении дистонии широко используются физиотерапия и массаж. Занятия с психологом помогают детям снять излишнее напряжение, скоррегировать отношения с окружающими, снизить уровень тревоги.
ВЛИЯНИЕ ВЕГЕТАТИВНОЙ НЕРВНОЙ РЕГУЛЯЦИИ НА РАЗВИТИЕ СЕРДЕЧНЫХ АРИТМИЙ У БОЛЬНЫХ ИШЕМИЧЕСКОЙ БОЛЕЗНЬЮ СЕРДЦА | Опубликовать статью ВАК, elibrary (НЭБ)
Лысенкова Н.О.1, Румянцев М.И.2, Жилина А.Н.3, Кратнов А.Е.4
1Кандидат медицинских наук, ассистент кафедры, 2Кандидат медицинских наук, ассистент кафедры, 3Кандидат медицинских наук, ассистент кафедры, Доктор медицинских наук, Профессор, заведующий кафедрой, Ярославский государственный медицинский университет
ВЛИЯНИЕ ВЕГЕТАТИВНОЙ НЕРВНОЙ РЕГУЛЯЦИИ НА РАЗВИТИЕ СЕРДЕЧНЫХ АРИТМИЙ У БОЛЬНЫХ ИШЕМИЧЕСКОЙ БОЛЕЗНЬЮ СЕРДЦА
Аннотация
У больных сердечно-сосудистыми заболеваниями, особенно ишемической болезнью сердца (ИБС), отмечается выраженное ремоделирование миокарда, происходит перестройка вегетативной нервной системы сердца. Это ведет к нарушению как симпатических, так и парасимпатических регуляторных влияний. Рассогласования между электрофизиологическими процессами в миокарде, эфферентной импульсацией из центральных структур и состоянием афферентных связей, а также дисфункция самих центральных нервных структур могут служить причиной электрической нестабильности миокарда и способствовать возникновению фатальных нарушений сердечного ритма, приводящих к наступлению кардиальной смерти.
Ключевые слова: вегетативная нервная система, ишемическая болезнь сердца, электрическая нестабильность миокарда, нарушения ритма сердца.
Lysenkova N.O.1, Rumyancev M.I.2, Zhilina A.N.3, Kratnov A.E.4
1MD, assistant of the Department of therapy of pediatric faculty, 2MD, assistant of the Department of therapy of pediatric faculty, 3MD, assistant of the Department of therapy of pediatric faculty, 4MD, Professor, Head of the Department of therapy of pediatric faculty, Yaroslavl State Medical University
THE INFLUENCE OF AUTONOMIC NERVOUS REGULATION ON THE DEVELOPMENT OF CARDIAC ARRHYTHMIAS IN PATIENTS WITH CORONARY HEART DISEASE
Abstract
In patients with cardiovascular diseases, especially coronary heart disease, there is a marked remodeling of the myocardium, there is a rearrangement of the autonomic nervous system of the heart. This leads to disruption of both sympathetic and parasympathetic regulatory influences. The mismatch between electrophysiological processes in the myocardium, with efferent impulses from the central agencies and the state of the afferent connections, and also themselves dysfunction of the central nervous structures can cause electrical instability of the myocardium and lead to fatal cardiac arrhythmias leading to cardiac death occurrence.
Keywords: autonomic nervous system, coronary heart disease, electrical instability of the myocardium, arrhythmias.
Одной из актуальных проблем современной кардиологии является своевременное определение у больных ИБС риска развития угрожающих жизни аритмий и наступления кардиальной смерти [1]. Данные клинических и морфологических исследований доказывают тесную взаимосвязь развития кардиальной смерти с наличием фатальных нарушений ритма, дисфункцией ВНС, нейрогуморальными механизмами, и сниженной ФВ левого желудочка у больных ИБС [17]. Определяющим условием для возникновения фатальных аритмий признается наличие структурной патологии сердца, которое под действием различных функциональных факторов становится электрически нестабильным [25]. Кроме того, существуют данные, согласно которым нестабильность электрофизиологических процессов в миокарде у пациентов, страдающих ИБС, в первую очередь связана с нарушением регуляции вегетативного и электролитного баланса [7, 19].
Сердце иннервируется вегетативной нервной системой (ВНС), состоящей из симпатических и парасимпатических нервов. Симпатические нервы, стимулируя β-адренорецепторы синусового узла, увеличивают частоту сердечных сокращений (ЧСС). Раздражение блуждающего нерва, в свою очередь, стимулирует М-холинорецепторы синусового узла, вследствие чего развивается брадикардия. Синусовый и атриовентрикулярный узлы находятся в основном под влиянием блуждающего нерва и, в меньшей степени, симпатического, в то время как желудочки контролируются преимущественно симпатическим нервом [28, 37]. Деятельность ВНС контролируется центральной нервной системой и рядом гуморальных влияний. В продолговатом мозге расположен сердечно-сосудистый центр, который объединяет парасимпатический, симпатический и сосудодвигательный центры. Регуляция этих центров осуществляется подкорковыми узлами и корой головного мозга. На ритмическую деятельность сердца влияют импульсы, исходящие из сердечно-аортального, синокаротидного и других сплетений. Также на сердечно-сосудистый центр действуют гуморальные нарушения, изменения в крови (парциального давления углекислого газа и кислорода, кислотно-основного состояния), хеморецепторный рефлекс [11]. В состоянии покоя доминирует тонус вагуса и изменчивость сердечной периодики в значительной степени зависит от его влияний. Вагусная и симпатическая активность находятся в постоянном взаимодействии. Поскольку синусовый узел богат холинэстеразой, действие любого вагусного импульса краткосрочно, так как ацетилхолин быстро гидролизируется. Преобладание парасимпатических влияний над симпатическими может быть объяснено двумя независимыми механизмами: холинергически индуцируемым снижением высвобождения норадреналина в ответ на симпатическую стимуляцию и холинергическим подавлением ответа на адренергический стимул. Афферентная вагусная стимуляция приводит к рефлекторному возбуждению эфферентной вагусной активности и ингибированию эфферентной симпатической активности [37]. Эффекты противоположно ориентированного рефлекса опосредуются стимуляцией афферентной симпатической активности [33]. Эфферентная вагусная активность также находится под тоническим сдерживающим влиянием афферентной кардиальной симпатической активности [16]. Эфферентная симпатическая и вагусная импульсации, направленные на синусовый узел, характеризуются разрядом, преимущественно синхронизированным с каждым сердечным циклом.
Литературные данные о роли вегетативной регуляции в аритмогенезе неоднозначны. В последнее время активно обсуждается положение о том, что повышение активности симпатического звена ВНС при ишемии миокарда приводит к возникновению нарушений ритма, тогда как активация парасимпатического звена обладает протективным эффектом [36]. Однако, при гистологическом исследовании миокарда у пациентов, погибших внезапно, было выявлено нарушение автономной регуляции, обширные очаги истощения катехоламинов в адренергических сплетениях миокарда и изменения вегетативных нервных ганглиев. Показано, что ишемические изменения в области нижней стенки левого желудочка вызывают активацию парасимпатического отдела ВНС, а в передней стенке приводят к повышению тонуса симпатических афферентных нервов [3].
Известно, что у больных ИБС, отмечается выраженное ремоделирование миокарда, происходит перестройка вегетативной нервной системы сердца. Это ведет к нарушению как симпатических, так и парасимпатических регуляторных влияний. Особого внимания заслуживает активация симпатоадреналовой системы и снижение активности парасимпатического отдела ВНС, связанные как с развитием общего адаптационного синдрома, так и со значительной структурной перестройкой миокарда, что обуславливает увеличение электрической нестабильности, склонность к возникновению фатальных нарушений сердечного ритма [2, 9, 10, 18], тем самым повышается риск кардиальной смерти.
Известно, что вагусное влияние понижает порог возникновения угрожающих жизни желудочковых аритмий и обеспечивает “антиаритмическую защиту”, возможно, путем снижения возбудимости кардиомиоцитов, а симпатическое, напротив, повышает этот порог, что приводит к более частым аритмическим осложнениям. Выявлено, что в остром периоде инфаркта миокарда (ИМ) наблюдается повышение тонуса симпатической нервной системы и снижение тонуса парасимпатической. В ряде исследований выявлено, что снижение вагусной активности или нарушение баланса влияний ВНС на синусовый ритм в пользу симпатического отдела, наблюдаемое уже в ранние сроки обострения ИБС, сопряжено с тяжестью заболевания и сохраняется не менее 6-12 месяцев [12, 15, 29]. Есть основания полагать, что этот дисбаланс связан с повышенным риском развития тяжелых осложнений и смерти: так, существуют свидетельства проаритмического эффекта уменьшения вагусных или повышения симпатическихвлияний на сердце, а также защитного действия противоположных изменений активности ВНС [10].
Исследования показали, что большинству эпизодов ишемии миокарда (61,8%) предшествовали значимые изменения тонуса ВНС (симпатической – 61,9%, парасимпатической – 38,1%). Остальные эпизоды (38,8%) возникали на фоне неизмененной активности ВНС. Установлено, что повышение активности симпатической нервной системы достоверно уменьшает продолжительность эпизодов безболевой ишемии миокарда и увеличивает длительность приступов спонтанной стенокардии [4]. Органические изменения миокарда, особенно ИМ, вносят значительные изменения в регуляторные процессы автономной нервной системы вследствие развития “вегетативной денервации” сердца. Нарушение вегетативной регуляции сердечно-сосудистой системы может быть обусловлено усилением симпато-симпатических [13, 31, 32] и симпато-вагальных рефлексов [37]. Изменение геометрических свойств пораженного миокарда приводит к усилению афферентной симпатической стимуляции вследствие механического раздражения нервных окончаний и рефлекторному ослаблению парасимпатических влияний на сердце [31]. Также снижается ответ клеток синусового узла на изменение нейрорегуляторных механизмов.
Достаточно хорошо освещен обсуждаемый в течение длительного времени вклад ВНС в генезис нарушений ритма сердца и внезапной кардиальной смерти у больных ИБС [14]. Известно, что по мере прогрессирования ИБС зоны ишемии, а также фиброза и кардиосклероза становятся более чувствительными к воздействию катехоламинов и поэтому реагируют на малейшие изменения симпатического тонуса [27]. В последние годы выявлено, что очаговые повреждения нервных волокон в желудочках сердца у больных с перенесенным ИМ могут приводить к нарушению реполяризации и, таким образом, способствовать электрической нестабильности миокарда [7, 8].Обнаружено, что при ИМ и нестабильной стенокардии нарушения симпатической иннервации сердца превышают по площади и глубине зоны нарушенной перфузии, что объясняется большей чувствительностью нейрональных окончаний к ишемии. Предполагается, что усиление адренергических влияний на кардиомиоциты у больных с ОКС происходит вследствие снижения захвата симпатическими окончаниями сердца поступающего из кровотока норадреналина [7].
Известно, что нервная регуляция стабилизирует электрическую активность сердца. Повреждения внутрисердечных нервных волокон и ганглиев могут лежать в основе серьезных нарушений образования импульсов, проведения возбуждения и нарушения процессов реполяризации миокарда. Длительные сравнительные клинические и патологоанатомические наблюдения свидетельствуют о том, что у больных с ВКС нервные волокна часто изменены вблизи проводящей системы сердца [20, 21]. Также встречаются очаговые повреждения нервных волокон в желудочках сердца, которые могут приводить к нарушению реполяризации и способствовать электрической нестабильности миокарда [42]. В литературе описаны связи между нервными волокнами и кардиомиоцитами, особенно расположенными вблизи проводящей системы, где в большом количестве проходят периферические нервы [22, 23, 43]. Не исключено, что поражение нервных сплетений в желудочках сердца, где определяется большое количество симпатических нервов [41], вызывают нарушения реполяризации из-за изменения адренергических нервных влияний. Вблизи синусового узла могут повреждаться как симпатические, так и парасимпатические нервные элементы, и естественно предположить, что поражения нервных волокон разной медиаторной природы будет вызывать противоположное действие.
Таким образом, эпидемиологические, клинические и морфологические данные убедительно доказывают, что регуляция ВНС является одним из наиболее важных механизмов, стабилизирующих электрическую активность сердца. А повреждения внутрисердечных нервных волокон и ганглиев у больных ИБС, могут лежать в основе серьезных нарушений ритма сердца.
Литература
- Bojcov S.A., Nikulina N.N., Jakushin S.S. i dr. Vnezapnaja serdechnaja smert’ u bol’nyh IBS: rasprostranennost’, vyjavljaemost’ i problemy statisticheskogo ucheta // Rossijskij kardiologicheskij zhurnal. -2011.-№2.-S.59-64.
- Bokerija L.A., Revishvili A.Sh., Neminushhij N.M. Vnezapnaja serdechnaja smert’. – M.: GJeOTAR-media, 2011. – 272 s.
- Vahljaev V.D., Nedostup A.V., Caregorodcev D.A. i soavt. Rol’ gumoral’nyh faktorov v patogeneze aritmij serdca // Rossijskij medicinskij zhurnal. -2000.-№2.-S. 54-57
- Dzizinskij A.A., Smirnova Ju.Ju., Beljalov F.I. Ocenka aktivnosti vegetativnoj nervnoj sistemy pri pristupe ishemii miokarda s pomoshh’ju issledovanija variabel’nosti ritma // Kardiologija. -1999.-№1.-S. 34-37.
- Ivanov G.G., Smetnev A.S., Syrkin A.L. i dr. Osnovnye mehanizmy, principy prognoza i profilaktiki vnezapnoj serdechnoj smerti // Kardiologija.-1998.-№12.-S. 64-73
- Kalinkin M.N., Dubrovin I.A., Chelnokov V.S. Strukturno-metabolicheskie osnovy central’nyh mehanizmov vnezapnoj serdechnoj smerti // Kardiologija. -2000.-№4.-S. 30-33
- Nifontov E.M., Shihaliev D.R., Bogachev M.I. i dr. Antiaritmicheskaja jeffektivnost’ omega-3 polinenasyshhennyh zhirnyh kislot u bol’nyh stabil’noj ishemicheskoj bolezn’ju serdca s zheludochkovymi narushenijami ritma // Kardiologija. -2010.-№12.-S.:15-17.
- Jakushin S.S., Bojcov S.A., Furmenko G.I., i dr. Vnezapnaja serdechnaja smert’ u bol’nyh ishemicheskoj bolezn’ju serdca po rezul’tatam Rossijskogo mnogocentrovogo jepidemiologicheskogo issledovanija Zabolevaemosti, smertnosti, kachestva diagnostiki i lechenija ostryh form IBS (REZONANS) // Rossijskij kardiologicheskij zhurnal. -2011.-№2.-S. 59-64
- Algra A., Tijssen J.G.P., Roelandt J.R.T.C., et al. Heart rate variability from 24-hour electrocardiography and the 2-year risk for sudden death // Circulation. -1993.- Vol. 88, №1.-P. 180-185.
- Bernardi L., Salvucci F., Suardi R., et al. Evidence for an intrinsic mechanism regulating heart rate variability in the transplanted and the intact heart during submaximal dynamic exercise? // Cardiovasc Res. – 1990.-№24.-P. 969-981.
- Bigger J.T., Fleiss J.L., Rolnitzky L.M., et at. Time course of recovery of heart period variability after myocardial infarction // J Am Coll Cardiol. -1991.- Vol. 18, №7.-P. 1643-1649
- Brown A.M., Malliani A. Spinal sympathetic reflexes initiated by coronary receptors // J Physiol. – 1971-№212.-P. 685-705.
- Cao J.M., Fishblin M.C., Ham J.B. et al. Relationship between regional cardiac hyperinnervation and ventricular arrhythmia // Curculation. – 2000.-Vol. 101, №16.-P. 1960-1969.
- Casolo G.C., Stroder P., Signorini С., et al. Heart rate variability during the acute phase of myocardial infarction // Circulation. – 1992.- Vol. 85, №6.-P. 2073-2079
- Cerati D., Schwartz P.J. Single cardiac vagal fiber activity, acute myocardial ischemia, and risk for sudden death // Circ Res. – 1991.-№69.-P. 1389-1401.
- Chen S.W. A wavelet-based heart rate variability analysis for the study of nonsustained ventricular tachycardia // Trans Biomed Eng. – 2002.- Vol. 49, №7.-P. 736-742
- Ewing D.J. Heart Rate Variability: New Risk Factor in Patients Following Myocardial Infarction // Clin Cardiol. – 1991.-№14.-P. 683-685
- ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST- segment elevation // European Heart Journal. – 2012.- doi:10.1093/eurheartj/ehs 215.
- James T.N. De Subitaneis Mortibus. Apoplexy of the heart // Circulation. – 1978.-P. 385-391.
- James T.N., Zipes D.P., Finegan F.E., Eisele J.W., Carter J.E. Cardiacganglionitis associated with sudden unexpected death // Ann. Intern. Med. – 1979/- №91.- 727-730.
- James T. N. Cholinergic mechanisms in the sinus node with particular reference to the actions of hemicholinium // Circ. Res. – 1966.-. №19.-P. 347-357.
- James T. N., Bear E. S., Lang K. F., Green E W., Winkler H. H. Adrenergic mechanisms in the sinus node // Arch, Intern. Med. – 1970.-№125.-P. 512-547.
- Griffin B.P., Topol E.J. The manual of cardiovascular medicine. – LWW, 2013. – 1192.
- Kaasik A., Ristimae T., Soopold U. The relationship between left ventricular mass and ventricular late potential in patients with myocardial infarction // J Coronary Artery Disease. – 2001.- Vol. 4, №1.-P. 60.
- Kent K.M., Smith E.R., Redwood O.R., et al. Electrical stability of acutely ischemic myocardium. Influences of heart rate and vagal stimulation // Circulation. -1973.- Vol.47, №2.-P. 291-298
- Leenhardt A., Maison-Blancke P., Denjoy I. et al. Mechanism of spontaneous occurence of tachycardia // Arch Mal Coeur Vaiss. – 1999.- Vol. 92, №1.-P. 17-22
- Levy M.N. Sympathetic-parasympathetic interactions in the heart // Circ Res. – 1971.-№29.-P. 437-445.
- Lombardi F., Sandrone G., Pempruner S., et al. Heart rate variability as an index of sympathovagal interaction after acute myocardial infarction // Am J Cardiol. -1987.- Vol. 60, №16.-P. 1239-1245.
- Lown B., Verrier R.L. Neural activity and ventricular fibrillation // N Engl J Med. -1976.- Vol. 294, №21.- 1165-1170.
- Malfatto G., Rosen T.S., Steinberg S.F. et al. Sympathetic neural modulation of cardiac impulse initiation and repolarization in the newborn rat // Circ Res. – 1990.-№66.-P. 427-437.
- Malliani A., Recordati G., Schwarz P.J. Nervous activity of afferent cardiac sympathetic fibers with atrial and ventricular endings // J Physiol. -1973.-№229.-P. 457-469.
- Malliani A. Cardiovascular sympathetic afferent fibers // Rev Physiol Biochem Pharmacol. – 1982.-№94.-P. 11-74
- Malliani A., Schwartz P.J., Zanchetti A. A sympathetic reflex elicited by experimental coronary occlusion // Am J Physiol. – 1969.-№217.-P. 703-709.
- Nannenberg, E. A., Sijbrands, E. J. G., Dijksman, et al. Mortality of Inherited Arrhythmia Syndromes / Clinical Perspective // Circulation: Cardiovascular Genetics. – 2012.- Vol. 5, №2.-P. 183-189.
- Osaka M., Saitoh H., Sasabe N. et al. Changes in autonomic activity preceding onset of nonsustained ventricular tachycardia // Ann Noninvasive Electrocardiol. – 1996.- Vol. 1, №1.- 3-11.
- Schwarz P.J., Pagani M., Lombardi F., Malliani A., Brown A.M. A cardio-cardiac sympatho-vagal reflex in the cat // Circ Res. – 1973.-№32.-P. 215-220.
- Saul J.P., Berger R.D., Albrecht P., Stein S.P., Chen M.N., Cohen R.J. Transfer function analysis of the circulation: Unique insights into cardiovascular regulation // Am J Physiol. – 1991.-№261.-P. 1231-1245.
- Schwartz P.J., Vanoli E., Stramba-Badiale M., De Ferrari G.M., Billman G.E., Foreman R.D. Autonomic mechanisms and sudden death. New insights from analysis of baroreceptor relexes in conscious dogs with and without a myocardial infarction // Circulation. – 1988.- Vol. 78, №4.-P. 969-979.
- Schwarz P.J., Pagani M., Lombardi F., Malliani A., Brown A.M. A cardio-cardiac sympatho-vagal reflex in the cat // Circ Res. – 1973.-№32.-P. 215-220.
- Shvalev V.N., Stropus R.A., Abraytis R.I., et al. Ultrastructural and histochemical studies of the cardiac nervous system and the hypothalamohypophyseal-adrenal system in sudden cardiac death. In: Sudden Cardiac Death, Proceedings of the Third USA-USSR Joint Symposium, Kaunas, USSR, June 29-July 1, 1982. Washington, D.C.: U.S. Department of Health & Human Services, Public Health Service, National Institutes of Health. // DNEW Рublication no. (NIH) 84–1604.- 1984.-P. 115-139.
- Third Universal Definition of Myocardial Infarction // European Heart Journal. – 2012.- doi:10.1093/eurheartj/ehs 184.
- Urthaler F., Katholi С.R., Macy J., James T.N. Electrophysiological and mathematical characteristics of the escape rhythm during complete AV block // Cardiovasc. Res. – 1974.-№8.-P. 173-186.
The Parasympathetic Nervous System | Protocol (Translated to Russian)
18.2: Парасимпатическая нервная система
Обзор
Парасимпатическая нервная система является одним из двух основных отделов вегетативной нервной системы. Эта парасимпатическая система отвечает за регулирование многих бессознательных функций, таких как пульс и пищеварение. Он состоит из нейронов, расположенных как в головном мозге, так и в периферической нервной системе, которые отправляют свои аксоны на мышцы, органы и железы.
Система «Отдых и дайджест»
Активация парасимпатической системы, как правило, имеет расслабляющее воздействие на организм, способствуя функции, которые пополняют ресурсы и восстановить гомеостаз. Поэтому иногда его называют системой «отдых и переваривание». Парасимпатическая система преобладает в спокойные времена, когда безопасно выделять ресурсы на основные функции «домашнего хозяйства» без угрозы нападения или вреда.
Управление парасимпатической системой
Парасимпатическая нервная система может быть активирована различными частями мозга, включая гипоталамус. Преганглионные нейроны в стволе мозга и сакральной части спинного мозга сначала отправляют свои аксоны в ганглии -кластеры нейрональных клеток органов — в периферической нервной системе. Эти ганглии содержат связи между до- и постганглионные нейроны и расположены рядом с органами или железами, которые они контролируют. Отсюда постганглионные нейроны отправляют свои аксоны на ткани-мишени – как правило, гладкие мышцы, сердечную мышцу или железы. Как правило, нейромедиатор ацетилхолин используется для регулирования деятельности этих целей.
Активация парасимпатической системы вызывает различные эффекты на организм. Это снижает сердечный ритм и заставляет зрачков сжиматься, восстанавливая тело в более расслабленном состоянии. Он также стимулирует пищеварение и выделение, например, путем содействия слюноотделение, перистальтические сокращения в желудке и кишечнике, и сокращение мочевого пузыря, чтобы изгнать мочу. Это помогает восстановить запасы энергии, вызывая поджелудочной железы, чтобы выделяет больше инсулина. Наконец, он даже способствует размножению за счет увеличения притока крови к половым органам.
Стоя
Контроль сердечного ритма и кровотока имеет важное значение даже для повседневных задач, таких как стоя. Люди, которые страдают от ортостатической непереносимости (ОИ) могут испытывать хроническую легкомысленность и обморок от простого акта попасть в вертикальной позе, называется ортостаз. Вегетативная нервная система контролирует необходимые изменения сосудов и сердечного ритма, когда мы участвуем в ортостазе. В частности, парасимпатическая система отвечает за сигналы, которые позволяют вазодилатации-расслабление мышц вокруг слизистой оболочки кровеносных сосудов- из мозговых артерий. Неправильная сигнализация парасимпатической нервной системы может привести к потере сознания из-за недостаточного притока крови к мозгу.
Литература для дополнительного чтения
Stewart, Julian M. “Common Syndromes of Orthostatic Intolerance.” Pediatrics 131, no. 5 (May 2013): 968–80. [Source]
Определение вегето-сосудистой дистонии | Поликлиника «Медицинский комплекс – Елец»
Вегето-сосудистая дистония является мультифакториальным заболеванием, возникающим, как правило, на фоне стрессовых ситуаций, ведущих к нарушению нейро-гуморальной и эндокринной регуляции различных систем организма, что проявляется нарушением адаптации к меняющимся условиям внешней и внутренней среды и множеством разнообразных клинических симптомов и синдромов.
В клинической практике используют разные синонимы: нейроциркуляторная дистония, вегето-сосудистая дистония, вегетативный невроз, вегетоз, синдром вегетативной дисфункции, нейроциркуляторная астения.
Под синдромом вегетативной дисфункции (СВД) понимают полисистемные расстройства, возникающие в результате нарушения деятельности надсегментарных вегетативных структур. Клинические проявления СВД (рубрика G.90 по МКБ-10) характеризуются полиморфизмом, при котором в патологический процесс вовлекаются многие системы организма, при этом ведущее место отводится расстройствам сердечно-сосудистой системы.
В нашей стране это заболевание традиционно рассматривается как психовегетативный синдром, или СВД. Данный термин подчеркивает облигатность и приоритет психических нарушений и не ограничивает вегетативные расстройства рамками одной системы. При этом необходимо учитывать механизм соматизации, который определяется, как склонность переживать психологический стресс на физиологическом уровне. Одним из важнейших отличительных признаков рассматриваемого расстройства считается соматосенсорное усиление (англ.: somatosensory amplification), под которым понимают стабильное свойство личности усиленно фокусировать внимание на соматических симптомах, из-за чего усиливается субъективное восприятие физических ощущений и повышается опасность того, что эти неприятные физические ощущения будут ошибочно расценены как болезненные.
В современной психиатрии СВД чаще всего рассматривается как «Соматоформная дисфункция вегетативной нервной системы» (по МКБ-10 рубрика F45.3). Данное понятие объединяет группу психогенных заболеваний, характеризующихся патологическими симптомами, напоминающими соматическое заболевание, но при которых по результатам обследования не обнаруживается никаких морфологических проявлений, хотя при этом часто имеются неспецифические функциональные нарушения.
Вегетативная нервная система представлена в коре головного мозга, гипоталамической области, мозговом стволе, спинном мозге, имеются также периферические отделы вегетативной нервной системы. Наличие патологического процесса в любой из этих структур, а также функциональное нарушение связи между ними могут обусловить появление вегетативных расстройств.
Функции, выполняемые вегетативной нервной системой (ВНС) в организме, чрезвычайно важны: ею осуществляется контроль и регуляция деятельности внутренних органов, обеспечивающие поддержание гомеостаза – постоянного баланса внутренней среды. По своему функционированию ВНС является автономной, т.е. не подчиняющейся сознательному, волевому контролю и другим отделам нервной системы. Вегетативной нервной системой обеспечивается регуляция множества физиологических и биохимических процессов: поддержание терморегуляции, оптимального уровня АД, процессов метаболизма, мочеобразования и пищеварения, эндокринной, сердечно-сосудистой, иммунной реакций и т.д.
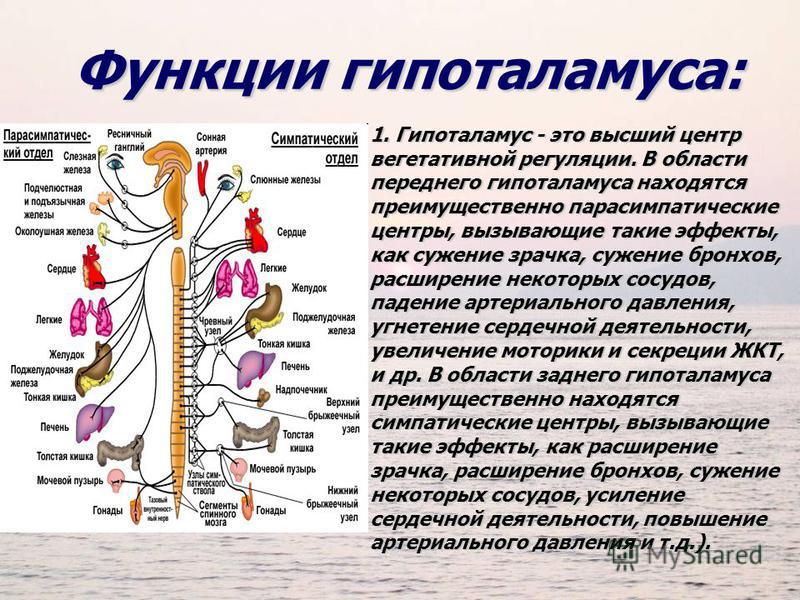
ВНС состоит из симпатического и парасимпатического отделов, оказывающих противоположные эффекты на регуляцию различных функций. К симпатическим эффектам влияния ВНС относятся расширение зрачка, усиление обменных процессов, повышение АД, уменьшение тонуса гладкой мускулатуры, увеличение ЧСС, учащение дыхания. К парасимпатическим – сужение зрачка, снижение АД, повышение тонуса гладкой мускулатуры, уменьшение ЧСС, замедление дыхания, усиление секреторной функции пищеварительных желез и т.д.
|
Орган |
Симпатическая иннервация | Парасимпатическая иннервация |
|
Глаза |
Расширяет зрачок, расширение глазной щели, выстояние глазного яблока. | Суживает зрачок, сужение глазной щели, западение глазного яблока. |
|
Слюнные железы |
Вызывает малообильную густую слюну. | Вызывает усиленное выделение жидкой водянистой слюны. |
|
Сердце |
Вызывает тахикардию (усиленное сердцебиение) повышает кровяное давление. | Вызывает брадикардию (замедленное сердцебиение) понижает кровяное давление. |
|
Бронхи |
Расширяет бронхи, уменьшает выделение слизи. | Суживает бронхи, увеличивает выделение слизи. |
|
Пищевод, желудок, кишечник |
Понижает секрецию (выработку пищеварительного сока), ослабляет перистальтику (сокращение мышц стенки ЖКТ, продвигающее пищу). | Повышает секрецию, усиливает перистальтику, вызывает спазмы. |
|
Кожа |
Суживает сосуды, вызывает побледнение, «гусиную кожу», уменьшает потоотделение. | Расширяет сосуды, вызывает покраснение, усиливает потоотделение. |
Нормальная деятельность ВНС обеспечивается согласованностью функционирования симпатического и парасимпатического отделов и их адекватной реакцией на изменения внутренних и внешних факторов. Нарушение баланса между симпатическими и парасимпатическими эффектами ВНС вызывает развитие вегето-сосудистой дистонии.
Причины и развитие вегето-сосудистой дистонии
В последние годы отмечается неуклонный и повсеместный рост заболеваемости нейроциркуляторной дистонией (НЦД), что обусловлено нарастанием темпов социальных, экономических, технологических и даже климатических изменений, и требует от человека всё возрастающей адекватной адаптации к условиям жизнедеятельности.
По мнению В.Н. Мясищева, причина патологии — результат взаимодействия психогенных факторов с аномалиями (врожденными или приобретенными) вегетативной иннервации. Причинами расстройств вегетативной регуляции являются наследственная предрасположенность, патология перинатального периода, последствия черепно-мозговых травм и нейроинфекций, очаги хронического воспаления и соматические заболевания, чрезмерные физические нагрузки, неблагоприятные условия внешней среды и др.
Развитие вегето-сосудистой дистонии у детей младшего возраста может быть обусловлено патологией перинатального периода (внутриутробной гипоксией плода), родовыми травмами, заболеваниями периода новорожденности. Эти факторы отрицательно сказываются на формировании соматической и вегетативной нервной системы, полноценности выполняемых ими функций. Вегетативная дисфункция у таких детей проявляется пищеварительными нарушениями (частые срыгивания, метеоризм, неустойчивый характер стула, плохой аппетит), эмоциональным дисбалансом (повышенной конфликтностью, капризностью), склонностью к простудным заболеваниям.
В период полового созревания развитие внутренних органов и рост организма в целом опережает становление нейроэндокринной регуляции, что ведет к усугублению вегетативной дисфункции. В этом возрасте вегето-сосудистая дистония проявляется болями в области сердца, перебоями и сердцебиением, лабильностью артериального давления, психоневрологическими расстройствами (повышенной утомляемостью, снижением памяти и внимания, вспыльчивостью, высокой тревожностью, раздражительностью).
У взрослых пациентов возникновение вегето-сосудистой дистонии может провоцироваться и усугубляться в связи с влиянием хронических заболеваний, депрессий, стрессов, неврозов, черепно-мозговых травм и травм шейного отдела позвоночника, эндокринных болезней, патологий ЖКТ, гормональных перестроек (беременности, климакса). В любом возрасте фактором риска развития вегето-сосудистой дистонии является конституциональная наследственность.
Симптомы вегето-сосудистой дистонии
Для вегетососудистой дистонии, являющейся следствием нарушения тонуса сосудов, характерны следующие симптомы:
- психоневрологические симптомы ВСД – слабость, вялость, повышенная утомляемость, низкая работоспособность, головокружения, головные боли, расстройства сна, обмороки, снижение сексуальной активности, панические атаки;
- терморегуляторные симптомы ВСД, связанные с нарушением потоотделения и терморегуляции – самопроизвольное повышение температуры тела, чрезмерная потливость, жар, приливы, либо, наоборот, зябкость конечностей
- сосудистые — нестабильность артериального и венозного давления;
- кардиологические – перебои в работе сердца, учащенные сердечные сокращения и боли в области сердца;
- дыхательные (респираторные) в рамках гипервентиляционного синдрома– одышка, тяжесть в груди и ощущение нехватки воздуха;
- диспепсические симптомы ВСД – расстройство работы желудочно – кишечного тракта (запор, понос), нарушения мочеиспускания и дискинезия желчного пузыря.
Скачать и читать статью полностью…
Текущий обзор регулирования покоя вегетативных почек в JSTOR
AbstractВ этом обзоре мы исследуем современные методы и недавние достижения, направленные на понимание клеточных механизмов, участвующих в контроле покоя вегетативных отростков. Вегетативные отростки (включая стебли, корневища, клубни, луковицы, столоны, ползучие корни и т. Д.) Содержат пазушные и придаточные почки, способные давать новые стебли / ветви в благоприятных условиях.Пазушные и придаточные почки отличаются тем, что пазушные почки образуются в пазухах листьев и отвечают за образование боковых побегов (ветвей). Придаточные почки относятся к почкам, которые возникают на растении в местах (стеблях, корнях или листьях), кроме пазух листьев. Как пазушные, так и придаточные почки обычно находятся в состоянии покоя. Покой описывается как временная приостановка видимого роста любой структуры растения, содержащей меристему (Lang et al. 1987). Состояние покоя можно разделить на три категории: (1) остановка экодормальности находится под контролем внешних факторов окружающей среды; (2) остановка парадормальности находится под контролем внешних физиологических факторов внутри растения; и (3) остановка эндодормальности находится под контролем внутренних физиологических факторов.Общей чертой всех этих процессов является предотвращение роста в условиях, когда в противном случае рост должен продолжаться. Появляется все больше доказательств того, что отсутствие роста происходит из-за блокировки клеточного деления в результате взаимодействий между сигнальными путями, контролирующими покой, и теми, которые контролируют клеточный цикл.
Информация журналаWeed Science публикует оригинальные исследования и стипендии, направленные на понимание того, «почему» происходят явления. Таким образом, он сосредоточен на фундаментальных исследованиях, непосредственно связанных со всеми аспектами науки о сорняках.
Информация для издателейCambridge University Press (www.cambridge.org) — издательское подразделение Кембриджского университета, одного из ведущих исследовательских институтов мира и лауреата 81 Нобелевской премии. В соответствии со своим уставом издательство Cambridge University Press стремится максимально широко распространять знания по всему миру. Он издает более 2500 книг в год для распространения в более чем 200 странах. Cambridge Journals издает более 250 рецензируемых академических журналов по широкому кругу предметных областей в печатном виде и в Интернете.Многие из этих журналов являются ведущими научными публикациями в своих областях, и вместе они составляют одну из наиболее ценных и всеобъемлющих областей исследований, доступных сегодня. Для получения дополнительной информации посетите http://journals.cambridge.org.
miR156 включает изменение вегетативной фазы при регуляции окислительно-восстановительных сигналов в проростках яблони.
Poethig, R. S. Изменение фазы и регуляция времени развития у растений. Наука 301 , 334–346 (2003).
ADS CAS Статья PubMed Google Scholar
Поэтиг, Р. С. Вегетативное фазовое изменение и созревание побегов у растений. Curr Top Dev Biol 105 , 125–152 (2013).
CAS Статья PubMed PubMed Central Google Scholar
Zhang, X. Z. et al . Возможные полифенольные маркеры фазового перехода яблони ( Malus domestica ). J Физиол растений 164 , 574–580 (2007).
CAS Статья PubMed Google Scholar
Хакетт У. П. Молодость, созревание и омоложение древесных растений. Horti Rev p: 109–56 (1985).
Циммерман Р. Х., Хакетт В. П. и Фарис Р. П. Гормональные аспекты фазового перехода и преждевременного цветения в гормональной регуляции развития III. Springer 11 , 79–115 (1985).
Google Scholar
Xiao, Z. F. et al. . Потеря молодости вызывает у подвоев яблони придаточную сопротивляемость укоренению. Культ органа ткани клеток растений 119 , 51–63 (2014).
Артикул Google Scholar
Мейлан, Р. Цветочная индукция у древесных покрытосеменных растений. Новые леса 14 , 179–202 (1997).
Артикул Google Scholar
Wu, G. & Poethig, R. S. Временная регуляция развития побегов у Arabidopsis thaliana с помощью miR156 и его мишени SPL3 . Развитие 133 , 3539–3547 (2006).
CAS Статья PubMed PubMed Central Google Scholar
Ву, Г. и др. . Последовательное действие miR156 и miR172 регулирует время развития у Arabidopsis . Ячейка 138 , 750–759 (2009).
CAS Статья PubMed PubMed Central Google Scholar
Du, Z. et al . Редокс-гомеостаз и поглотители активных форм кислорода смещаются во время онтогенетических фазовых изменений у яблони. Завод Sci 236 , 283–294 (2015).
CAS Статья PubMed Google Scholar
Джонс-Роудс, М. В., Бартель, Д. П. и Бартель, Б. МикроРНК и их регуляторные роли в растениях. Анну Рев Завод Биол 57 , 19–53 (2006).
CAS Статья PubMed Google Scholar
Schwab, R. et al. . Специфические эффекты микроРНК на транскриптом растений. Dev Cell 8 , 517–27 (2005).
CAS Статья PubMed Google Scholar
Ким, Дж. и др. . Направленное микроРНК расщепление мРНК ATHB15 регулирует развитие сосудов в стеблях соцветий Arabidopsis . Завод J 42 , 84–94 (2005).
CAS Статья PubMed PubMed Central Google Scholar
Мэллори, А.С. и др. . Контроль микроРНК PHABULOSA в развитии листа: важность спаривания с 5′-областью микроРНК. EMBO J 23 , 3356–64 (2004).
CAS Статья PubMed PubMed Central Google Scholar
Франко-Соррилья, Дж. М. и др. . Мимикрия мишеней обеспечивает новый механизм регуляции активности микроРНК. Нат Генет 39 , 1033–1037 (2007).
CAS Статья PubMed Google Scholar
Ван, Дж. У. и др. . MiRNA контроль изменения вегетативной фазы у деревьев. PLoS Genet 7 , e1002012 (2011).
CAS Статья PubMed PubMed Central Google Scholar
Xu, X. Z. et al . Высокая экспрессия miR156 необходима для индуцированного ауксином образования придаточных корней посредством MxSPL26 независимо от PINs и ARFs у Malus xiaojinensis . Фасадный завод Sci 8 , 1059 (2017).
Артикул PubMed PubMed Central Google Scholar
Хирагури, А. и др. . Специфические взаимодействия между Dicer-подобными белками и дцРНК-связывающими белками семейства HYL1 / DRB в Arabidopsis thaliana . Завод Мол Биол 57 , 173–188 (2005).
CAS Статья PubMed Google Scholar
Kurihara, Y., Takashi, Y. & Watanabe, Y. Взаимодействие между DCL1 и HYL1 важно для эффективного и точного процессинга pri-miRNA в биогенезе микроРНК растений. РНК 12 , 206–212 (2006).
CAS Статья PubMed PubMed Central Google Scholar
Лоббс, Д., Раллапалли, Г., Шмидт, Д. Д., Мартин, К. и Кларк, Дж. СЕРРАЙТ: новый игрок на сцене микроРНК растений. Эмбо Реп 7 , 1052–1058 (2006).
CAS Статья PubMed PubMed Central Google Scholar
Донг, З., Хан, М. Х. и Федорофф, Н. РНК-связывающие белки HYL1 и SE способствуют точному процессингу in vitro pri-miRNA с помощью DCL1. P Natl Acad Sci USA 105 , 9970–9975 (2008).
ADS CAS Статья Google Scholar
Янг, С. В. и др. . Структура Arabidopsis HYPONASTIC LEAVES1 и его молекулярное значение для процессинга miRNA. Строение 18 , 594–605 (2010).
CAS Статья PubMed PubMed Central Google Scholar
Мачида, С., Чен, Х. Ю. и Юань, Ю. А. Молекулярное понимание процессинга миРНК с помощью Arabidopsis thaliana SERRATE. Нуклеиновые кислоты Res 39 , 7828–7836 (2011).
CAS Статья PubMed PubMed Central Google Scholar
МакКоннелл, Дж. Р. и Бартон, М. К. Влияние мутаций в гене PINHEAD из Arabidopsis на формирование апикальных меристем побегов. Dev Genet 16 , 358–366 (1995).
Артикул Google Scholar
Clarke, JH, Tack, DT, Findlay, K., Van Montagu, M. & Van Lijsebettens, M. Локус SERRATE контролирует формирование ранних молодых листьев и длину фазы у арабидопсиса Arabidopsis . . Завод J 20 , 493–501 (1999).
CAS Статья PubMed Google Scholar
Liu, Q. et al . Ген ARGONAUTE10 модулирует поддержание апикальной меристемы побегов и установление полярности листьев путем репрессии miR165 / 166 в Arabidopsis . Завод J 58 , 27–40 (2009).
CAS Статья PubMed Google Scholar
Джи, Л. и др. . ARGONAUTE10 и ARGONAUTE1 регулируют терминацию стволовых клеток цветка через две микроРНК в Arabidopsis . PLoS Genet 7 , e1001358 (2011).
CAS Статья PubMed PubMed Central Google Scholar
Li, S. X., Yang, X., Wu, F. J. и He, Y. K. HYL1 контролирует miR156-опосредованную ювенильную фазу вегетативного роста. J Exp Bot 63 , 2787–2798 (2012).
CAS Статья PubMed PubMed Central Google Scholar
Эшби, А. Исследования морфогенеза листьев I. Очерк формы листа. Новый Фитол 47 , 153–176 (1948).
Артикул Google Scholar
Янг, Л., Конвей, С. Р. и Поэтиг, Р. С. Изменение вегетативной фазы опосредуется сигналом, полученным из листьев, который репрессирует транскрипцию miR156. Развитие 138 , 2011, 245–249.
CAS Статья PubMed PubMed Central Google Scholar
Ю., С., Лиан, Х. и Ван, Дж. У. Переходы в развитии растений: роль микроРНК и сахаров. Curr Opin Завод Биол 27 , 1–7 (2015).
Артикул PubMed CAS Google Scholar
Wahl, V. et al. . Регулирование цветения с помощью передачи сигналов трегалозо-6-фосфата у Arabi d op sis thaliana . Наука 339 , 704–707 (2013).
ADS CAS Статья PubMed Google Scholar
Ван Дейкен, А.J., Schluepmann, H. & Smeekens, S. C. Arabidopsis трегалозо-6-фосфатсинтаза 1 необходима для нормального вегетативного роста и перехода к цветению. Физиология растений 135 , 969–977 (2004).
Артикул PubMed PubMed Central Google Scholar
Gomez, LD, Gilday, A., Feil, R., Lunn, JE & Graham, IA AtTPS1 -опосредованный синтез трегалозы-6-фосфата необходим для эмбриогенного и вегетативного роста и реакции на АБК при прорастании семена и замыкающие клетки устьиц. Завод J 64 , 1–13 (2010).
CAS PubMed Google Scholar
Yang, L., Xu, M. L., Koo, Y., He, Y. J. & Poethig, R. S. Сахар способствует изменению вегетативной фазы у Arabidopsis thaliana путем подавления экспрессии MIR156A и MIR156C. Элиф 2 , e00260 (2013).
PubMed PubMed Central Google Scholar
Ю., С. и др. . Сахар — это эндогенный сигнал для фазового перехода от молоди к взрослой особи у растений. Элиф 2 , e00269 (2013).
PubMed PubMed Central Google Scholar
Консидайн, М. Дж. И Фойер, К. Х. Редокс-регуляция развития растений. Знак антиоксид-окислительно-восстановительного потенциала 21 , 1305–1326 (2014).
CAS Статья Google Scholar
Фойер, К. Х. и Ноктор, Г. Редокс-гомеостаз и передача антиоксидантных сигналов: метаболический интерфейс между восприятием стресса и физиологическими реакциями. Растительная клетка 17 , 1866–1875 (2005).
CAS Статья PubMed PubMed Central Google Scholar
Петров В. Д., Ван Брейзегем Ф. Перекись водорода — центральный узел информационного потока в клетках растений. AoB Plants , pls014 (2012).
Миттлер Р., Вандераувера С., Голлери М. и Брейсегем Ф. В. Сеть генов реактивного кислорода растений. Тенденции Завод Sci 9 , 490–498 (2004).
CAS Статья PubMed Google Scholar
Li, S. W., Xue, L., Xu, S., Feng, H. & An, A. Перекись водорода действует как сигнальная молекула в формировании придаточных корней проростков маша. Environ Exp Bot 65 , 63–71 (2009).
CAS Статья Google Scholar
Sewelam, N. et al. . Пространственная H 2 O 2 сигнальная специфичность: H 2 O 2 из хлоропластов и пероксисом дифференцированно модулирует транскриптом растения. Завод Мол 7 , 1191–1210 (2014).
CAS Статья PubMed Google Scholar
Де Туллио, М. К., Цзян, К. и Фельдман, Л. Дж. Редокс-регуляция организации апикальной меристемы корня: связь развития корня с окружающей средой. Биохимия растений 48 , 328–336 (2010).
Артикул CAS Google Scholar
Шнаубельт, Д. и др. . Низкий уровень глутатиона регулирует экспрессию генов и окислительно-восстановительные потенциалы ядра и цитозоля у Arabidopsis thaliana . Среда растительных клеток 38 , 266–279 (2015).
CAS Статья PubMed Google Scholar
Баллатори, Н., Кранс, С. М., Марчан, Р. и Хаммонд, К. Л. Плазматические мембранные переносчики глутатиона и их роль в физиологии и патофизиологии клетки. Мол Аспект Мед 30 , 13–28 (2009).
CAS Статья PubMed Google Scholar
Noctor, G. et al. . Глутатион в растениях: комплексный обзор. Среда растительных клеток 35 , 454–484 (2012).
CAS Статья PubMed Google Scholar
Коффлер Б. Э., Блум Э., Зелльниг Г. и Зехманна Б. Визуализация с высоким разрешением субклеточных концентраций глутатиона с помощью количественной иммуноэлектронной микроскопии в различных областях листьев Arabidopsis . микрон 45 (2013). С. 119–128.
CAS Статья PubMed PubMed Central Google Scholar
Cochemé, H. M. et al. . Измерение H 2 O 2 в живых Drosophila во время старения с использованием логометрического масс-спектрометрического зонда, нацеленного на митохондриальный матрикс. Cell Metab 13 , 340–350 (2011).
Артикул PubMed PubMed Central CAS Google Scholar
Сохал Р. С. и Орр В. С. Гипотеза окислительно-восстановительного стресса старения. Свободные радикалы Biol Med 52 , 539–555 (2012).
CAS Статья Google Scholar
Ji, N., Wang, Y., Wu, T., Zhang, X. Z. & Han, Z.H. Корреляционный анализ экспрессии miR156 и окислительно-восстановительного статуса во время фазового изменения проростков Malus xiaojinensis . J China Agri Uni 21 , 59–64 (2016).
Google Scholar
Мёллер, И. М. Митохондрии растений и окислительный стресс: перенос электронов, оборот НАДФН и метаболизм активных форм кислорода. Annu Rev Plant Physiol Mol Biol. 52 , 561–591 (2001).
Артикул Google Scholar
Ferguson, M., Mockett, R.J., Shen, Y. & Orr, W.C. Возрастное снижение митохондриального дыхания и транспорта электронов у Drosophila melanogaster . Biochem J 390 , 501–511 (2005).
CAS Статья PubMed PubMed Central Google Scholar
Саги, М. и Флур, Р. Производство активных форм кислорода растительными НАДФН-оксидазами. Физиология растений 14 , 336–340 (2006).
Артикул CAS Google Scholar
Дитц, К. Дж., Тюркан, И. и Кригер-Лишкай, А. Редокс- и активные формы кислорода, зависящие от передачи сигналов в фотосинтезирующий хлоропласт и из него. Физиология растений 171 , 1541–1550 (2016).
CAS Статья PubMed PubMed Central Google Scholar
Xia, R., Zhu, H., An, Y.Q., Eric, P. B. & Liu, Z. R. Apple miRNA и tasiRNA с новыми регуляторными сетями. Геном биол 13 , R47 (2012).
CAS Статья PubMed PubMed Central Google Scholar
Вс, К., Zhao, Q., Liu, D. D., You, C. X. & Hao, Y. J. Эктопическая экспрессия гена APP e Md-miRNA 156 h регулирует развитие цветков и плодов в Arabidopsi s. Культ органа ткани клеток растений 112 , 343–351 (2013).
CAS Статья Google Scholar
Yu, N., Niu, Q. W., Ng, K. H. & Chua, N.H. Роль модулей miR156 / SPLs в развитии боковых корней Arabidopsis . Завод J 83 , 673–685 (2015).
CAS Статья PubMed Google Scholar
Martin, M. N., Saladores, P.H., Lambert, E., Hudson, A.O. & Leustek, T. Локализация членов семейства γ-глутамилтранспептидазы позволяет идентифицировать участки гидролиза глутатиона и S-конъюгата глутатиона. Физиология растений 144 , 1715–1732 (2007).
CAS Статья PubMed PubMed Central Google Scholar
Zhang, H.Q. & Forman, H.J. Редокс-регуляция γ-глутамилтранспептидазы. Am J Resp Cell Mol 41 , 509–515 (2009).
CAS Статья Google Scholar
Noctor, G., Mhamdi, A., Queval, G. & Foyer, C.H. Регулирование «привратника» окислительно-восстановительного потенциала: вакуумная секвестрация заменяет дисульфид глутатиона. Физиология растений 163 , 665–671 (2013).
CAS Статья PubMed PubMed Central Google Scholar
Гуо, К. К. и др. . Репрессия miR156 с помощью miR159 регулирует время перехода от молоди к взрослой у Arabidopsis . Растительная клетка 29 , 1293–1304 (2017).
PubMed Google Scholar
Петров В. Д. и Бриусегем Ф. В. Перекись водорода — центральный узел информационного потока в клетках растений. Заводы AoB 1 , pls014 (2012).
Google Scholar
Noctor, G. & Foyer, C.H. Внутриклеточная редокс-компартментация и связанная с ROS коммуникация в регуляции и передаче сигналов. Физиология растений 171 , 1581–1592 (2016).
CAS Статья PubMed PubMed Central Google Scholar
Miret, J. A. & Munne-Bosch, S. Передача сигналов окислительно-восстановительного потенциала и устойчивость растений к стрессу: в центре внимания витамин E. Ann NY Acad Sci 1340 , 29–38 (2015).
ADS CAS Статья PubMed Google Scholar
Чен, Дж., Чжу, К., Ли, Л. П., Сан, З. Ю. и Пан, X. Б. Влияние экзогенной салициловой кислоты на рост и H 2 O 2 -метаболизирующие ферменты в проростках риса в условиях свинцового стресса. J Environ Sci 19 , 44–49 (2007).
CAS Статья Google Scholar
Zhang, X. L., Jia, X. F., Yu, B., Gao, Y. & Bai, J. G. Экзогенная перекись водорода влияет на активность антиоксидантных ферментов и перекисное окисление липидов в листьях огурцов при слабом освещении. Sci Hortic 129 , 656–662 (2011).
CAS Статья Google Scholar
Amini, SA, Shabani, L., Афгани, L., Jalalpour, Z. & Sharifi-tehrani, M. Вызванное сквалестатином производство таксола и баккатина в суспензионной культуре клеток тиса ( Taxus baccata Л.). Турок Дж Биол 8 , 2014. С. 528–536.
Артикул CAS Google Scholar
Voehringer, D. W., McConkey, D. J., McDonnell, T. J., Brisbay, S. & Meyn, R. E. Экспрессия Bcl-2 вызывает распределение глутатиона в ядре. P Natl Acad Sci USA 95 , 2956–2960 (1998).
ADS CAS Статья Google Scholar
Виванкос, П. Д. и др. . Рекрутирование глутатиона в ядро во время пролиферации клеток регулирует окислительно-восстановительный гомеостаз всей клетки у Arabidopsis thaliana и снижает защиту от окисления. Завод J 64 , 825–838 (2010).
CAS Статья PubMed Google Scholar
Гарсия-Хименес, Дж. Л. и др. . Ядерный глутатион. Biochim Biophys Acta 1830 , 3304–3316 (2013).
CAS Статья PubMed Google Scholar
Огава, К. Глутатион-связанная регуляция роста растений и реакции на стресс. Сигнал Antiox Rad. 7 , 973–981 (2005).
CAS Статья Google Scholar
Пастернак Т., Асард Х., Поттерс Г. и Янсен М. А. К. Тиоловые соединения глутатион и гомоглутатион по-разному влияют на развитие клеток люцерны ( Medicago sativa L .). Физиол растений Биохимия 74 , 16–23 (2014).
CAS Статья PubMed Google Scholar
Gao, Y. et al. . Различия в экспрессии и регуляции генов при изменении онтогенетической фазы у проростков яблони. Завод Мол Биол Реп 32 , 357–371 (2013).
Артикул CAS Google Scholar
Zhu, L. et al. .Зеараленон вызывает апоптоз и некроз в клетках гранулезы свиней через каспазу-3- и каспазу-9-зависимый митохондриальный сигнальный путь. J Cell Physiol 227 , 1814–1820 (2012).
CAS Статья PubMed Google Scholar
Дуда, М., Кнет, М., Табаровски, З. и Сломчинска, М. Кондиционированная среда с лютеиновыми макрофагами влияет на стероидогенез в клетках гранулезы свиней. Репрод Биол 11 , 117–34 (2011).
Артикул PubMed Google Scholar
Zhang, Y. G., Chen, J. H., Han, Z. H., Xu, X. F. и Li, T. Z. Сравнение методов выделения тотальной РНК из Malus xiaojinensis и амплификации кДНК LD-PCR. Биотех Бык 4 , 50–53 (2004).
Google Scholar
Гриффитс-Джонс, С., Грокок, Р. Дж., Ван Донген, С., Бейтман, А. и Энрайт, А. J. miRBase: последовательности микроРНК, мишени и номенклатура генов. Нуклеиновые кислоты Res 34 , D140 – D144 (2006).
CAS Google Scholar
Гриффитс-Джонс, С., Сайни, Х. К., ван Донген, С. и Энрайт, А. J. miRBase: инструменты для геномики микроРНК. Нуклеиновые кислоты Res 36 , Д154–158 (2008).
CAS Google Scholar
Xing, L. B. et al. . Полногеномная идентификация микроРНК, связанных с вегетативным фазовым переходом, и прогнозирование мишеней с использованием секвенирования деградома в Malus hupehensis . BMC Genomics 15 , 1125 (2014).
Артикул PubMed PubMed Central CAS Google Scholar
Мукерджи, С. П. и Чоудхури, М. А. Влияние системы перекись водорода-аскорбат на проницаемость мембран подвергшихся воздействию воды проростков vigna . Новый Фитол 99 , 355–360 (1985).
CAS Статья Google Scholar
Yang, W. et al. . Перекись водорода является вторым посредником в побочном процессе укоренения проростков маша, вызванном салициловой кислотой. PLoS ONE 8 , e84580 (2013).
ADS Статья PubMed PubMed Central CAS Google Scholar
Darius, C. et al. . Характеристика генов НАДФН-оксидазы яблока и их экспрессии, связанной с окислительным стрессом, в культуре побегов in vitro . Культ органа ткани клеток растений 124 , 621–633 (2016).
Артикул CAS Google Scholar
Liang, Z. C. et al. . Наследование содержания сахара и кислоты в спелых ягодах тетраплоидной × диплоидной популяции винограда. Euphytica 182 , 251–259 (2011).
CAS Статья Google Scholar
Garg, N. & Pandey, R. Высокая эффективность экзотических арбускулярных микоризных грибов отражается в улучшении ризобиального симбиоза и оборота трегалозы у генотипов Cajanus cajan , выращенных в условиях солевого стресса. Экология грибов 21 , 57–67 (2016).
Артикул Google Scholar
Характеристика вегетативной регуляции ЧСС и центральной гемодинамики в тренировках юных гимнасток
Л.Н. Ботова, аспирант
Т.Г. Кириллова, доцент, к.э.н.
Приволжская государственная академия физической культуры, спорта и туризма, г. Казань
Ключевые слова: типологическая характеристика вегетативной регуляции, вариабельность сердечного ритма, центральная гемодинамика, юные гимнастки.
Введение. В современном детско-юношеском спорте отмечается рост нагрузки. Нагрузка у юных спортсменов часто выше, чем у спортсменов старшего возраста. Поэтому в детских спортшколах необходимо ввести жесткий контроль за здоровьем и спортивной формой юных спортсменов, а также за их переносимостью не только тренировочных, но и соревновательных нагрузок.
Учитывая эти особенности, можно сделать раннюю диагностику утомления и проследить адаптационные процессы организма молодого спортсмена для предотвращения состояний утомления и перетренированности [1-6].
Западные писатели часто анализируют в своих работах влияние пола, возраста и многоцелевых тренировок спортсменов на значение показателей ВСР [7-10]. Однако в проанализированных работах западных и большинства отечественных авторов мы не обнаружили каких-либо описаний индивидуально-типологических особенностей вегетативной регуляции сердечного ритма у спортсменов.
Целью исследования было определение индивидуально-типологических характеристик вегетативной регуляции сердечного ритма и показателей центральной гемодинамики (ЦГД) у гимнасток 8–11 лет, определение взаимосвязи характеристик вариабельности сердечного ритма. центральная гемодинамика у гимнасток с разными типами вегетативной регуляции и особенности реакции вариабельности сердечного ритма и характеристики центральной гемодинамики по типу вегетативной регуляции на тренировочную нагрузку.
Методы исследования . Зарегистрировано и проанализировано 264 записи временных и спектральных показателей ВСР, а также показателей ИБС у гимнасток 8-11 лет в недельном тренировочном цикле (предсезонный микроцикл) до и после тренировок. В качестве устройства ввода данных использовали диагностический прибор «Варикард 2,51» (г. Рязань, 2008 г.) и программу «Иским-6» для анализа полученной информации. ЧСС регистрировали в положении лежа на спине, в стандартном отведении II, в течение пяти минут. .Типы вегетативной регуляции определяли по классификации профессора Н.И. Шлык (1992). Все гимнастки были допущены к занятиям спортом по заключению медицинского обследования, проведенного в диспансере лечебной физкультуры.
Состояние сердечно-сосудистой системы исследовали на диагностическом оборудовании «Валента» (Санкт-Петербург, 2007) реографическим методом. Показатели центральной гемодинамики регистрировали с помощью интегральной реографии (ИР) Тищенко (1968).Систолическое и диастолическое артериальное давление (АД и АД в сутки) измеряли с помощью механического тонометра.
Результаты и обсуждение. В результате исследования установлен широкий межиндивидуальный разброс показателей ВСР у гимнасток 8-11 лет. Используя индивидуальный подход к оценке типа вегетативной регуляции сердечного ритма, мы опирались на идею двухэтапной модели управления сердечным ритмом и на классификацию, предложенную профессором Н.И. Шлык (1992).Как следствие, выявлены гимнастки с разным состоянием регуляторных систем: с умеренно преобладающей центральной регуляцией — тип I (n = 22), с выраженным преобладанием центральной регуляции — тип II (n = 47), с умеренно преобладающей автономной регуляцией. — III (n = 51) и выраженное преобладание автономного регулирования — IV тип (n = 12). Определенные различия выявлены при сравнении показателей ВСР у юных гимнасток с преобладанием автономной и центральной регуляции сердечного ритма.В таблице 1 представлены результаты анализа ВСР юных гимнасток 8-11 лет в зависимости от преобладающего типа вегетативной регуляции
Таблица 1. Вариабельность сердечного ритма Показатели гимнасток 8-11 лет с разными типами вегетативной регуляции в покое (M ± m)
Автономное регулирование | |||||||
Тип вегетативной регуляции (TVR) | ЧСС (уд / мин) | MxDMn (MC) | RMSSD (мк) | SI (ок.ед.) | HF, (%) | LF, (%) | |
Тип I n = 22 (16,67%) | 91,73 ± 1,27 ** | 226,45 ± 7,54 ** | 41,64 ± 2,47 ** | 182,09 ± 13,80 ** | 40,07 ± 2,46 ** | 27,23 ± 1,21 | |
Тип II = 47 (35,6%) | 93,45 ± 1,69 ** | 188,94 ± 6,75 ** | 36,36 ± 2,10 ** | 304,06 ± 32,72 ** | 47,88 ± 2,47 ** | 26,21 ± 1,92 | |
Тип III n = 51 (38,64%) | 80,65 ± 0,85 | 369,29 ± 8,61 | 80,76 ± 2,96 | 56,98 ± 2,73 | 53,22 ± 2,14 | 25,29 ± 1,45 | |
Тип IV n = 12 (9,09%) | 69,00 ± 1,39 ** | 534,00 ± 14,88 ** | 138,42 ± 6,78 ** | 19,08 ± 1,25 ** | 52,77 ± 3,06 | 23,03 ± 2,03 | |
Центральное регулирование | |||||||
TVR | TP (MC 2 ) | ВЧ (MC 2 ) | LF (MC 2 ) | VLF (MC 2 ) | ULF (MC 2 ) | ||
Тип I n = 22 (16,67%) | 2160,16 ± 159,65 ** | 902,65 ± 93,44 ** | 592,42 ± 56,23 ** | 329,75 ± 10,23 ** | 335,34 ± 47,18 | ||
Тип II = 47 (35,6%) | 1328,69 ± 94,13 ** | 677,52 ± 61,97 ** | 335,86 ± 26,93 ** | 147,96 ± 7,91 ** | 165,22 ± 17,54 ** | ||
Тип III n = 51 (38,64%) | 5171,63 ± 275,14 | 2878,00 ± 231,70 | 1238,32 ± 87,66 | 551,55 ± 36,95 | 503,77 ± 58,86 | ||
Тип IV n = 12 (9,09%) | 12357,01 ± 899,99 ** | 6508,89 ± 550,69 ** | 2824,46 ± 306,57 ** | 1258,19 ± 243,53 ** | 1765,47 ± 419,96 ** | ||
* достоверность отличий относительно 3-го типа равна (Р ≤ 0,05)
** достоверность отличий относительно 3-го типа равна (Р ≤ 0,01)
Полученные данные ВСР показали, что гимнастки с I, II и IV типами вегетативной регуляции достоверно различались по показателям ВСР гимнасток с III типом вегетативной регуляции (P ≤ 0,01).При усреднении показателей ВСР гимнасток получены значения, близкие к значениям показателей ВСР гимнасток с III типом вегетативной регуляции. Таким образом, усреднение показателей ВСР без учета типа вегетативной регуляции недопустимо, поскольку искажает истинное состояние регуляторных систем и, в свою очередь, дискредитирует этот метод.
Мы также разделили данные ИБС на 4 группы в зависимости от типов вегетативной регуляции (табл. 2). Установлено, что гимнастки с разными типами вегетативной регуляции различаются (относительно гимнасток с III типом вегетативной регуляции) и по показателям ИБС, т.е.е. MBV у гимнасток с разными типами регуляции поддерживается за счет включения разных механизмов.
Таблица 2. Показатели центральной гемодинамики юных гимнасток (8-11 лет) с разными типами вегетативной регуляции в покое (M ± m)
ТВР | ЧСС (уд / мин) | SV (мл) | МБВ (л) | РУ (%) | SVI (мл / с) | СИ (мл / с) | SPR (мм рт. Ст. / Мл / с) |
Тип I n = 22 (16,67%) | 88,59 ± 1,74 ** | 60,18 ± 2,52 * | 5,27 ± 0,16 | 252,50 ± 8,60 | 62,17 ± 2,25 | 5,48 ± 0,18 | 12,41 ± 0,64 |
Тип II = 47 (35,6%) | 91,19 ± 1,61 ** | 62,11 ± 1,65 * | 5,58 ± 0,12 | 259,57 ± 5,36 | 65,27 ± 1,32 * | 5,88 ± 0,11 | 10,96 ± 0,30 |
Тип III n = 51 (38,64%) | 77,78 ± 1,31 | 69,72 ± 2,50 | 5,34 ± 0,16 | 256,16 ± 8,03 | 72,59 ± 2,43 | 5,59 ± 0,18 | 11,82 ± 0,43 |
Тип IV n = 12 (9,09%) | 66,17 ± 1,46 ** | 73,62 ± 2,83 | 4,85 ± 0,18 | 205,17 ± 8,24 * | 70,98 ± 3,51 | 4,69 ± 0,25 * | 13,75 ± 0,66 * |
* достоверность различий относительно G 3 равна (Р ≤ 0,05),
** достоверность различий относительно G 3 равна (Р ≤ 0,01)
По данным таблицы 2 установлено, что самые низкие значения индексов MBV и SVI, а также самые высокие значения индексов HR и MBV наблюдаются у гимнасток с преобладанием центральной регуляции (типы I и II).Высокие значения индекса MBV у этих гимнасток подтверждаются высокими значениями индекса HR, т.е. наблюдается снижение функциональных возможностей организма, так как поддержание уровня MBV за счет HR неэкономично. Наименьшие значения ЧСС, МБВ и максимальные значения показателей СК и СПР наблюдаются у гимнасток с IV типом вегетативной регуляции. Оптимальные параметры ИБС отмечены у гимнасток с умеренной активностью автономной регуляции (тип III), эти спортсменки обладают высокими функциональными возможностями организма и могут достигать высоких спортивных результатов.При усреднении значений показателей ИБС установлено, что средние значения этих показателей не отличаются, за исключением индекса ЧСС, от показателей ИБС у гимнасток с III типом вегетативной регуляции.
Анализ реакции организма на тренировочную нагрузку — еще один критерий оценки физической подготовленности спортсменов к высоким физическим нагрузкам, включая соревнования (таблицы 3 и 4). Гимнастки с I типом вегетативной регуляции сердечного ритма в большинстве случаев отреагировали на тренировочную нагрузку повышенным индексом SI (p ≤ 0,05).Посттренировочная тенденция к увеличению значения индекса SV без повышения ЧСС у этих гимнасток обусловлена повышенным напряжением механизмов ИБС и регуляторных систем.
Гимнастки со II типом вегетативной регуляции отличаются гипореактивностью регуляторных систем к тренировочной нагрузке и несоответствием автономной и центральной регуляции. Следовательно, у этих гимнасток, скорее всего, будут низкие спортивные результаты. Посттренировочный индекс МБВ в большей степени увеличивается за счет увеличения индекса ЧСС.Индексы SPR и PR при этом существенно не изменились. Этот ответ характеризует нарушения регуляции из-за снижения функциональных возможностей организма.
Гимнастки с III типом вегетативной регуляции сердечного ритма отличаются адекватной реакцией сердечно-сосудистой системы на тренировочную нагрузку. За тренировками следует значительное увеличение ЧСС, SI (p ≤ 0,01). Повышение индекса MBV связано с увеличением показателей ЧСС и SV, вызванным повышенной симпатической активностью.Индекс периферического сопротивления также снижается (p ≤ 0,05) в результате адекватного ответа на повышение сердечной эффективности по снижению нагрузки на миокард.
Гимнастки с IV типом вегетативной регуляции сердечного ритма характеризуются повышением индекса напряжения регуляторных систем (SI) (p ≤ 0,01), снижением MxDMn, SDNN и индекса ULF (p ≤ 0, 05). Индекс MBV после тренировки также имеет тенденцию к увеличению из-за повышения в большей степени индекса HR.В этом случае отсутствие значимых изменений реакции на тренировочную нагрузку можно рассматривать как реакцию организма на чрезмерную или вынужденную нагрузку для данного возраста.
Таблица 3. Показатели вариабельности сердечного ритма у гимнасток (8-11 лет) с разными типами вегетативной регуляции в покое и после нагрузки (M ± m)
ЧСС (уд / мин) | MxDMn (MC) | RMSSD (мк) | SDNN (MC) | SI (ок.ед.) | ||||||||||||||
Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | |||||||||
Тип I n = 22 (16,67%) | 91,7 ± 1,3 | 94,4 ± 2,2 | 226,4 ± 7,5 | 212,3 ± 22,2 | 41,6 ± 2,5 | 39,9 ± 5,8 | 46,6 ± 1,7 | 41,8 ± 4,2 | 182,1 ± 13,8 | 392,4 ± 75,2 * | ||||||||
Тип II = 47 (35,6%) | 93,4 ± 1,7 | 94,7 ± 1,5 | 188,9 ± 6,7 | 195,5 ± 10,8 | 36,4 ± 2,1 | 46,6 ± 3,8 * | 37,5 ± 1,5 | 44 ± 3,1 | 304,1 ± 32,7 | 320,2 ± 33 | ||||||||
Тип III n = 51 (38,64%) | 80,6 ± 0,8 | 83,2 ± 1,3 | 369,3 ± 8,6 | 358,2 ± 16,8 | 80,8 ± 3 | 80,4 ± 5,1 | 78,6 ± 2,2 | 77,6 ± 4,2 | 57 ± 2,7 | 106,8 ± 18,4 ** | ||||||||
Тип IV n = 12 (9,09%) | 69,0 ± 1,4 ** | 79, 7 ± 2,9 | 534,0 ± 14,9 | 434,4 ± 43,9 ** | 138,4 ± 6, 8 | 107,1 ± 17,6 | 124,8 ± 4,6 | 92,1 ± 12,1 * | 19,1 ± 1,2 | 78,1 ± 21,2 ** | ||||||||
Центральное регулирование | ||||||||||||||||||
М ± м | TP (MC 2 ) | ВЧ (мк 2 ) | LF (MC 2 ) | VLF (MC 2 ) | ULF (MC 2 ) | |||||||||||||
Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | |||||||||
Тип I n = 22 (16,67%) | 2160,2 ± 159,6 | 1989,1 ± 370,7 | 902,65 ± 93,44 | 994,66 ± 243,54 | 592,4 ± 56,2 | 572,3 ± 104,4 | 329,7 ± 10,2 | 279,7 ± 41,3 | 335,3 ± 47,2 | 142,5 ± 25,6 ** | ||||||||
Тип II = 47 (35,6%) | 1328,7 ± 94,1 | 1703,5 ± 192,6 | 677,52 ± 61,97 | 898,43 ± 125,59 | 335,7 ± 26,9 | 454,4 ± 57,0 | 147,7 ± 7,9 | 197,1 ± 20,4 * | 165,2 ± 17,5 | 227,3 ± 29,9 | ||||||||
Тип III n = 51 (38,64%) | 5171,6 ± 275,1 | 5016,3 ± 439,5 | 2878,00 ± 231,70 | 2714,54 ± 277,55 | 1238,3 ± 87,7 | 1365,9 ± 124,3 | 551,5 ± 36,9 | 548,5 ± 70,6 | 503,8 ± 58,9 | 365,8 ± 42,6 | ||||||||
Тип IV n = 12 (9,09%) | 12357 ± 900 | 8323,7 ± 1757 | 6508,89 ± 550,69 | 4327,87 ± 987,55 | 2824,4 ± 306,6 | 2433 ± 531,3 | 1377,5 ± 239,7 | 943,7 ± 187,4 | 1765,5 ± 420 | 620,0 ± 165,6 * | ||||||||
* достоверность различий равна (Р ≤ 0,05)
** достоверность различий равна (Р ≤ 0,01)
Таблица 4 . Показатели центральной гемодинамики у гимнасток (8-11 лет) с разными типами вегетативной регуляции в покое и после нагрузки (M ± m)
ЧСС (уд / мин) | SV (мл) | МБВ (л) | Давление наполнения LV | ||||||
Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | ||
Тип I n = 22 (16,67%) | 88,59 ± 1,74 | 87,64 ± 2,24 | 60,18 ± 2,52 | 64,90 ± 2,63 | 5,27 ± 0,16 | 5,59 ± 0,16 | 27,91 ± 0,78 | 25,27 ± 1,43 | |
Тип II = 47 (35,6%) | 91,19 ± 1,61 | 93,45 ± 1,22 | 62,11 ± 1,65 | 65,31 ± 1,91 | 5,58 ± 0,12 | 6,04 ± 0,15 * | 26,94 ± 0,64 | 26,28 ± 0,72 | |
Тип III n = 51 (38,64%) | 77,78 ± 1,31 | 81,00 ± 1,23 | 69,72 ± 2,50 | 74,48 ± 2,60 | 5,34 ± 0,16 | 5,95 ± 0,17 * | 25,37 ± 0,85 | 22,24 ± 1,45 | |
Тип IV n = 12 (9,09%) | 66,17 ± 1,46 | 77,67 ± 3,06 ** | 73,62 ± 2,83 | 74,49 ± 4,86 | 4,85 ± 0,18 | 5,79 ± 0,44 | 28,83 ± 1,03 | 24,67 ± 1,93 | |
САД (мм рт. Ст.) | САД (мм рт. Ст.) | SPR (мм рт. Ст. / Мл / с) | PR (дин * сек / см 5 ) | ||||||
Перед тренировкой | Перед тренировкой | После тренировки | После тренировки | Перед тренировкой | После тренировки | Перед тренировкой | После тренировки | ||
Тип I n = 22 (16,67%) | 94,23 ± 1,90 | 100,45 ± 1,84 * | 53,86 ± 1,16 | 50,73 ± 1,80 | 12,41 ± 0,64 | 11,73 ± 0,47 | 1080,33 ± 37,96 | 1006,00 ± 34,66 | |
Тип II = 47 (35,6%) | 91,62 ± 1,09 | 97,15 ± 1,51 ** | 50,36 ± 0,75 | 49,15 ± 0,91 | 10,96 ± 0,30 | 10,85 ± 0,37 | 971,19 ± 22,67 | 941,72 ± 29,52 | |
Тип III n = 51 (38,64%) | 93,24 ± 1,46 | 98,25 ± 1,33 | 52,84 ± 1,05 | 52,94 ± 1,17 | 11,82 ± 0,43 | 11,06 ± 0,29 | 1045,98 ± 30,41 | 938,85 ± 17,17 * | |
Тип IV n = 12 (9,09%) | 93,92 ± 2,56 | 99,25 ± 2,31 | 51,42 ± 0,96 | 51,67 ± 1,27 | 13,75 ± 0,66 | 12,08 ± 0,96 | 1098,44 ± 22,99 | 971,78 ± 71,97 | |
* достоверность различий равна (Р ≤ 0,05)
** достоверность различий равна (Р ≤ 0,01)
В результате проведенных исследований выделены типологические характеристики вегетативной регуляции сердечного ритма и показателей центральной гемодинамики у гимнасток 8-11 лет, выявлена взаимозависимость характеристик вариабельности сердечного ритма и центральной гемодинамики у гимнасток с разным возрастом. типы вегетативной регуляции и определение особенностей реакции организма, которые помогут спортивным специалистам в будущем контролировать здоровье юных спортсменов, оптимизировать тренировочную нагрузку в соответствии с типом вегетативной регуляции сердечного ритма и контролировать переносимость спортсменами не только тренировок , но и конкурентоспособные нагрузки.
Список литературы
- Баевский, Р. Изучение механизмов вегетативной регуляции кровообращения на основе ортостатической пробы с использованием математического анализа частоты сердечных сокращений / Р.М. Баевский, В. Лаубе, А.П. Берсенева // Вестник Удмуртского университета. — Ижевск: опубл. г-гэ «Удмуртский университет», 1995. — № 3. — С. 13–20. (На русском языке)
- Гаврилова, Е.А. Спортивное сердце. Стресс-кардиомиопатия: монография / Е.А. Гаврилова. — М .: Советский спорт, 2007.- 200 с .: ил. (На русском языке)
- Иванова Н.В. Влияние специфики двигательной активности на вариабельность сердечного ритма спортсменов: по данным амбулаторного наблюдения: автореф. диссертация / Н.В. Иванова. — Ярославль, 2003. — 21 с.
- Кардиогемодинамика и физическая работоспособность спортсменов: Собрание сочинений / сост. автор R.A. Меркулова. — М .: Советский спорт, 2012. — 186 с .: ил. (На русском языке)
- Шлык, Н.Частота сердечных сокращений и тип регуляции у детей, подростков и спортсменов: монография / Н.И. Шлык. — Ижевск: опубл. г-гп «Удмуртский университет», 2009. — 255 с.
- Яблучанский, Н. Изменчивость частоты сердечных сокращений. Гид терапевта. Для профессиональных врачей / Н.И. Яблучанский, А. Мартыненко. — Харьков, 2010. — 131 с.
- Портье, Х., Луизи, Ф., Лауд, Д. и др. Интенсивные тренировки на выносливость в отношении вариабельности сердечного ритма и артериального давления у бегунов. Med Sci Sports Exerc 2001; 33 (7): 1120-5.
- Мальпас, С.С. Нейронные влияния на сердечно-сосудистую изменчивость: возможности и подводные камни. Am J Physiol Heart Circ Phys 2002; 282: H6-h30.
- Вариабельность сердечного ритма у спортсменов / Андре Э. Обер, Берт Сепс и Фрэнк Бекерс Лаборатория экспериментальной кардиологии, Медицинский факультет, K.U. Левен, Левен, Бельгия / Sports Med 2003. — 33 (12), с. 889-919.
- Иелламо, Ф. Нейронные механизмы сердечно-сосудистой регуляции во время физических упражнений. Auton Neurosci 2001; 20: 66-75.
Контакты автора: L.Botova@sportacadem.ru
Оптимальное распределение ресурсов семенам и вегетативным побегам при регулировании в зависимости от плотности у Syneilesis palmata (Compositae)
Американская генетическая ассоциация 1993. Проблема симпозиума AGA: эволюция пола. J. Hered. 84: 321–424.
Google Scholar
Белл, Г. 1985. Две теории пола и вариации. Experientia 41: 1235–1245.
Google Scholar
Бежичудек, П. 1982. Демография лесного многолетника, меняющего пол. Ecol. Monogr. 52: 335–351.
Google Scholar
Кальво Р. Н. 1993. Эволюционная демография орхидей: интенсивность и частота опыления и стоимость плодоношения. Экология 74: 1033–1042.
Google Scholar
Касуэлл, Х.1982. Стабильная популяционная структура и репродуктивная ценность популяций со сложными жизненными циклами. Экология 63: 1223–1231.
Google Scholar
Касуэлл, Х. 1985. Эволюционная демография клонального воспроизводства. Стр. 187–224. В: Джексон, Дж. Б. С., Басс, Л. В. и Кук, Р. Е. (ред.), Популяционная биология и эволюция клональных организмов. Издательство Йельского университета, Нью-Хейвен.
Google Scholar
Касуэлл, Х.1988. Приближение размера и возраста в матричных моделях населения. Стр. 85–105. В: Эбенман, Б. и Перссон, Л. (ред.), Популяции, структурированные по размеру. Springer-Verlag, Берлин.
Google Scholar
Касуэлл, Х. 1989. Матричные модели населения. Sinauer Associates, Массачусетс.
Google Scholar
Чарльзуорт, Б. 1994. Эволюция возрастных структур населения, 2-е издание.Издательство Кембриджского университета, Кембридж.
Google Scholar
Чиполлини М. Л., Уигхэм Д. Ф. и О’Нил Дж. 1993. Рост, структура и распространение семян в подлеске травы Cynoglossum virginianum : динамическая модель популяции и участка. Виды растений Biol. 8: 117–129.
Google Scholar
Фаулер, Н. 1988. Влияние неоднородности окружающей среды в пространстве и времени на регулирование популяций и сообществ.Стр. 249–269. В: Дэви, А. Дж., Хатчингс, М. Дж. И Уоткинсон, А. Р. (ред.), Экология популяций растений. Научные публикации Блэквелла, Оксфорд.
Google Scholar
Гиллман, М., Баллок, Дж. М., Сильвертаун, Дж. И Хилл, Б. С. 1993. Модель динамики популяции Cirsium vulgare , зависящая от плотности, с использованием значений параметров, оцененных на местах. Oecologia 96: 282–289.
Google Scholar
Ганский, И., Кууссаари М. и Ниеминен М. 1994. Структура метапопуляции и миграция у бабочки Melitaea cinxia . Экология 75: 747–762.
Google Scholar
Харада, Ю. и Иваса, Ю. 1994. Динамика популяции решеток для растений с рассеянием семян и вегетативным размножением. Res. Popul. Ecol. 36: 237–249.
Google Scholar
Хорвиц, К.C. & Schemske, D.W. 1986. Распространение семян и неоднородность окружающей среды в неотропической траве: модель популяции и динамика участков. Стр. 169–186. В: Эстрада, А. и Флеминг, Т. Х. (ред.), Плодоядные и распространение семян. DrW. Junk Publishers, Дордрехт.
Google Scholar
Херст, Л. Д. и Пек, Дж. Л. 1996. Последние достижения в понимании эволюции и сохранения пола. Trends Ecol. Evol. 11: 46–52.
Google Scholar
Иваса, Ю.1990. Эволюция самооценки и моделей распределения ресурсов. Виды растений Biol. 5: 19–30.
Google Scholar
Какехаши, М. и Харада, Ю. 1987. Теория репродуктивного распределения, основанная на демографии, зависящей от размера. Виды растений Biol. 2: 1–13.
Google Scholar
Кояма, Х. 1965. Цитотаксономические исследования композитов 1. Проблема видов на Syneilesis tagawae .Acta Phytotax. Геобот. 21: 129–132.
Google Scholar
Кояма, Х. 1967. Таксономические исследования трибы Senecioneae в Восточной Азии I. Общая часть. Mem. Coll. Sci. Univ. Киото, сер. В 33: 181–209.
Google Scholar
Лефкович, Л. П. 1965. Исследование роста популяции организмов, сгруппированных по стадиям. Биометрия 21: 1–18.
Google Scholar
Мэйнард Смит, Дж.1989. Эволюционная генетика. Издательство Оксфордского университета, Оксфорд.
Google Scholar
Мишод, Р. Э. и Левин, Б. Р. 1988. Эволюция пола: исследование современных идей. Sinauer Associates Inc., Массачусетс.
Google Scholar
Нишитани С. 1993. Репродуктивная экология Syneilesis palmata (Compositae). Диссертация, Токийский столичный университет.
Нишитани С. и Кимура М. 1993. Распределение ресурсов для полового и вегетативного размножения у лесной травы Syneilesis palmata (Compositae). Ecol. Res. 8: 173–183.
Google Scholar
Нишитани С. и Кимура М. 1995. Противопоставление демографических характеристик семян и вегетативных побегов в подлеске травы Syneilesis palmata (Compositae). Виды растений Biol.10: 1–10.
Google Scholar
Нишитани С., Такада Т. и Качи Н. 1995. Оптимальное распределение ресурсов для семян и вегетативных побегов в подлеске травы Syneilesis palmata (Compositae). Виды растений Biol. 10: 127–135.
Google Scholar
Нанни, Л. 1989. Поддержание секса путем группового отбора. Evolution 43: 245–257.
Google Scholar
Перри, Дж.Н. и Гонсалес-Андухар, Дж. Л. 1993. Рассеивание в модели района метапопуляции однолетнего растения с семенным фондом. J. Ecol. 81: 453–463.
Google Scholar
Рофф, Д. А. 1992. Эволюция историй жизни — теория и анализ. Чепмен и Холл, Нью-Йорк.
Google Scholar
Рафгарден, Дж. 1979. Теория популяционной генетики и эволюционной экологии: введение.Macmillan Publishing Co., Inc., Нью-Йорк.
Google Scholar
Сарухан Дж. 1974. Исследования по демографии растений: Ranunculus repens L., R. Bulbosus L. и R. acris L. II. Репродуктивные стратегии и динамика популяции семян. J. Ecol. 62: 151–177.
Google Scholar
Schellner, R.A., Newell, S.J. & Solbrig, O.Т. 1982. Исследования по популяционной биологии рода Viola . IV. Распространение раметок и проростков трех столонных видов. J. Ecol. 70: 273–290.
Google Scholar
Шни Б. К. и Уоллер Д. М. 1986. Репродуктивное поведение Amphicarpaea bracteata (Leguminosae), амфиккарпического однолетника. Амер. J. Bot. 73: 376–386.
Google Scholar
Стернс, С.C. 1985. Эволюция пола и роль пола в эволюции. Experientia 41: 1231–1235.
Google Scholar
Такада Т. 1995. Эволюция семелепестковых и итероплодных многолетних растений: сравнение динамики, не зависящей от плотности, и динамики, зависящей от плотности. J. theor. Биол. 173: 51–60.
Google Scholar
Такада Т. и Накадзима Х. 1992. Анализ эволюции жизненного цикла в терминах матричной модели Лефковича, зависящей от плотности.Математика. Biosci. 112: 155–176.
Google Scholar
Такада Т. и Накадзима Х. 1996. Оптимальное распределение для семенного и вегетативного размножения у многолетних растений: приложение к матричной модели перехода, зависящей от плотности. J. Theor. Биол. 182: 179–191.
Google Scholar
Такада Т. и Накадзима Х. 1998. Теоремы о процессе вторжения в популяциях с поэтапной структурой и динамикой, зависящей от плотности.J. Math. Биол. 36 (в печати).
Такада Т. и Накашизука Т. 1996. Демография в зависимости от плотности населения в умеренном широколиственном лесу Японии. Vegetatio 124: 211–221.
Google Scholar
ван Грюнендал, Дж., Де Крун, Х. и Касуэлл, Х. 1988. Матрицы проекций в популяционной биологии. Trends Ecol. Evol. 3: 264–269.
Google Scholar
Венейбл, Д.Л. и Браун, Дж. С. 1988. Избирательное взаимодействие рассеивания, покоя и размера семян как приспособления для снижения риска в изменчивой среде. Амер. Natur. 131: 360–384.
Google Scholar
Вернер П. А. и Касвелл Х. 1977. Темпы роста популяции и возраст в зависимости от моделей распределения по стадиям для тизели ( Dipsacus sylvestris Huds.). Экология 58: 1103–1111.
Google Scholar
Зейде, Б.1977. Репродуктивное поведение растений во времени. Амер. Natur. 112: 636–639.
Google Scholar
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Особенности вегетативной регуляции у больных сахарным диабетом 2 типа в зависимости от структуры сопутствующей патологии
20 -я Азиатско-Тихоокеанская конференция по диабету
16-17 июля 2018 г. Сидней, Австралия
Ирина Курникова
РУДН, Россия
Основной доклад : J Diabetes Metab
Аннотация :Диабет 2 типа (DT2) в сочетании с другими заболеваниями часто влияет на особенности прогрессирования диабета.Механизм, реализация этого влияния — вопрос для изучения. Проведен сравнительный анализ спектральных характеристик. сердечного ритма у больных ДТ2 с сопутствующими заболеваниями: 28 пациентов (группа-1) с артериальной гипертензией (АГ) и 17 пациентов (группа 2) с заболеваниями желудочно-кишечного тракта (ЖКТ) в сравнении (группа 3) 25 пациентов с CD2 без сопутствующие заболевания. Мощность частотного спектра оценивалась для HF, LF, VLF, ULF, индекса централизации IC. В в группе-1 наблюдались повышение НЧ и снижение ВЧ, что характерно для хронического стресса.УНЧ был выше в группа-3 (p = 0,0002; r = 0,48), что характерно для нарушения вегетативной регуляции. У двух пациентов из этой группы значения УНЧ занимали практически весь спектр (76% и 91%). В обоих случаях пациенты умирали от сосудистых катастроф в течение полутора лет. год, несмотря на активную терапию гипотензивными препаратами. Во 2-й группе активация центрального эрготропного и гуморального метаболизма. процессов, превышающих значение в группе-3 (p = 0,001; r = 0,37). Этот индикатор демонстрирует практически прямое влияние АГ на метаболический гомеостаз объясняет причины, по которым трудно добиться компенсации DT2 с помощью сопутствующая артериальная гипертензия и объясняет один из механизмов прогрессирования осложнений диабета.Увеличение IC в группе-2 подтверждена высокая активность центрального контура регуляции по отношению к вегетативному. Более того, это в прогностические условия, свидетельствовавшие об истощении регуляторных механизмов и высоком риске развития сосудистых нарушений (OR = 2,7; р = 0,001). Во второй группе направленность регуляторных процессов также свидетельствовала о хроническом стрессе.
Биография :Ирина Курникова — профессор медицины РУДН, Москва, Россия.Имеет опыт решения проблем эндокринологии более 20 лет. В настоящее время куратор научного направления эндокринология РУДН. Она опубликовала более 30 статей в известных журналах; Автор 25 книг и учебных пособий на русском языке и 10 патентов на изобретения. Область научных интересов — оптимизация системного подхода к лечению и реабилитации больных сахарным диабетом и заболеваниями щитовидной железы.
Электронная почта: [электронная почта защищена]
Вегетативное состояние
У пациентов в вегетативном состоянии цель с когнитивной точки зрения состоит в достижении «адекватного взаимодействия с окружающей средой» посредством добровольных и последовательных реакций на стимулы окружающей среды .Для этого врачи должны использовать общий подход через стимуляцию. Такой подход призван вселить в пациентов уверенность и позволить им снова разобраться в окружающем мире. Для этого мы используем две техники: базальную стимуляцию и мультисенсорную стимуляцию.
В отделении нейрореабилитации больниц vithas мы специализируемся на обследовании и лечении пациентов с измененным состоянием сознания ( кома, вегетативное состояние или синдром неотзывчивого бодрствования и состояние минимального сознания ).Фактически, наша исследовательская группа проводит постоянные исследования, направленные на улучшение нашего понимания нейронных изменений, которые происходят между состоянием комы и восстановлением сознания.
Поэтому мы разработали специальную программу терапии для пациентов с измененным состоянием сознания. Программа включает клиническое и неврологическое обследование, а также фармакологическое лечение, мультисенсорную и базальную стимуляцию, физиотерапию и осанку, а также стимуляцию орофациальных мышц и другие виды лечения. .
Наша медицинская помощь для пациентов с измененным состоянием сознания была недавно разработана с одобрения Федерации ассоциаций по повреждению головного мозга (FEDACE). Это сотрудничество привело к созданию первого в Испании справочника по синдрому безответного бодрствования и состояния минимального сознания. Справочник предоставляет новейшие знания по диагностике, оценке и лечению пациентов с измененным состоянием сознания. Он также предлагает практические отчеты наших специалистов в области здравоохранения, основанные на их обширном опыте лечения пациентов с измененным состоянием сознания (http: // fedace.