§ 5. Учение Чарлза Дарвина о естественном отборе
1. Выберите и подчеркните правильный ответ.
Какой закономерности подчиняется процесс размножения живых организмов при условии полного отсутствия ограничивающих факторов?
Ответ: арифметической прогрессии, геометрической прогрессии, логарифмической прогрессии.
2. Допишите определение.
Борьба за существование – это совокупность многообразных и сложных взаимоотношений, существующих между организмами и условиями внешней среды.
3. Назовите три формы борьбы за существование, которые выделил Ч. Дарвин.
1. Межвидовая.
2. Внутривидовая.
3. Борьба с неблагоприятными условиями внешней среды.
4. Объясните, почему внутривидовая борьба является наиболее напряженной.
У особей одного и того же вида потребности в пище, территории и других условиях существования одинаковы.
Кроме того, особи-самцы некоторых видов конкурируют между собой за самку. Поэтому данный вид борьбы является наиболее острым.

5. Охарактеризуйте организмы, выигрывающие в борьбе за существование.
В борьбе за существование выживают и оставляют потомство индивидуумы, которые обладают наследственным комплексом признаков и свойств, позволяющих наиболее успешно конкурировать с другими.
6. Объясните, что понимал Ч. Дарвин под естественным отбором.
Под естественным отбором Дарвин понимал происходящие в природе процессы избирательного уничтожения одних особей и преимущественного размножения других особей.
7. Закончите утверждение.
Материалом для естественного отбора является наследственная (индивидуальная, мутационная) изменчивость.
8. Объясните, почему изменчивость, обусловленная прямым воздействием внешней среды, не имеет значения для эволюции.
Изменчивость, обусловленная прямым воздействием внешней среды, не имеет значения для эволюции, так как не передается по наследству.
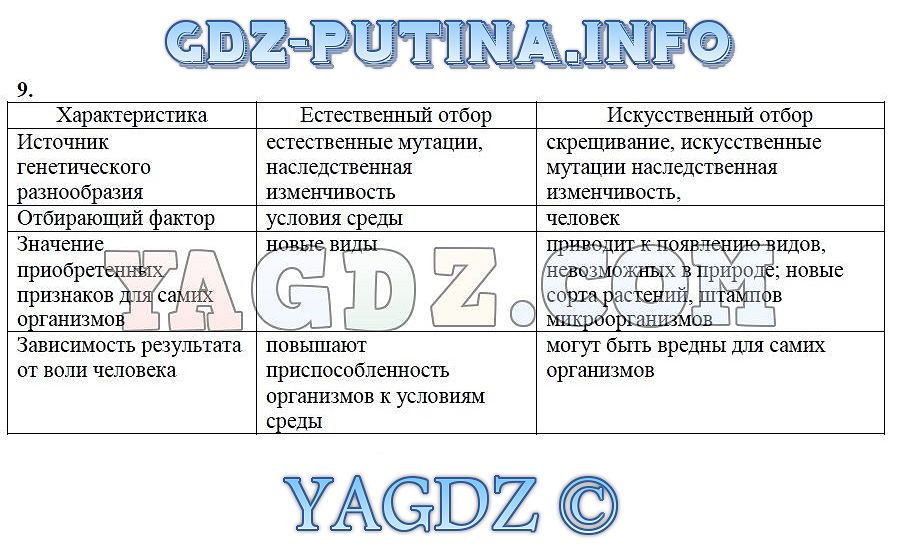
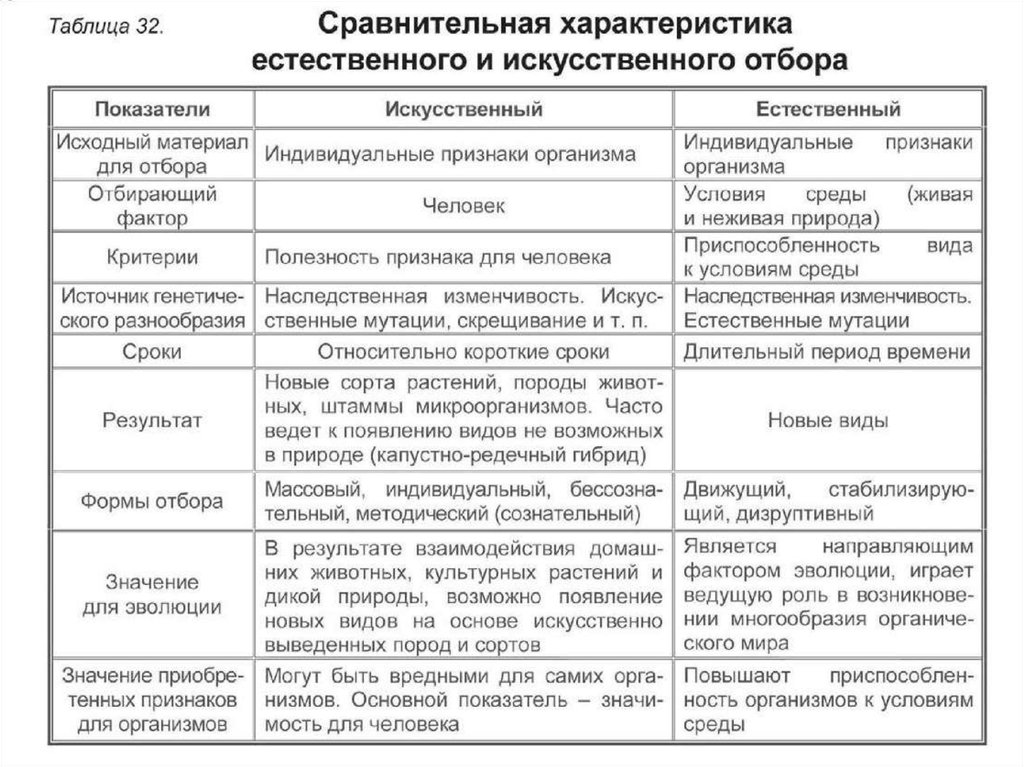
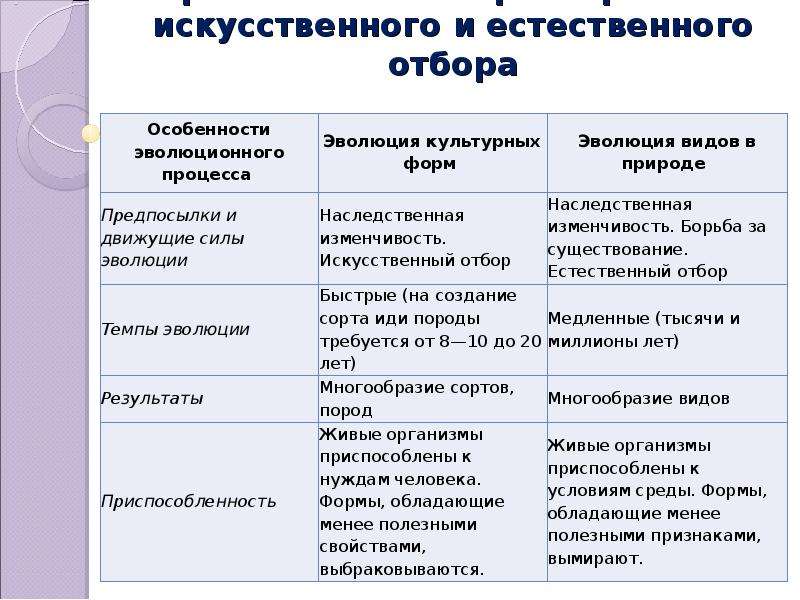
9. Дайте сравнительную характеристику естественного и искусственного отбора. Заполните таблицу.
Характеристика Естественный отбор Искусственный отбор Источник генетического разнообразия наследственная изменчивость, естественные мутации наследственная изменчивость, искусственные мутации, скрещивание Отбирающий фактор условия среды человек Значение приобретенных признаков для самих организмов новые виды новые сорта растений, штампов микроорганизмов. Приводит к появлению видов, невозможных в природе Зависимость результата от воли человека поышают приспособленность организмов к условиям среды могут быть вредны для самих организмов
АмГПГУНовая страница
В вашем браузере отключен JavaScript, поэтому некоторые возможности сайта будут недоступны.
Как включить JavaScript ?
Результаты 1 — 10 из 183
Начало | Пред. | 1 2 3 4 5 | След. | Конец | Все
| Изменено | Курс | Школа | Класс | ФИО | Обратный контакт | |||
|---|---|---|---|---|---|---|---|---|
| 03.11.2022 12:13:48 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ16 с УИОП | 10А | Куберская Полина Степановна | 79242251090 | |||
| 02.11.2022 19:17:56 | ИПиП Тренинг «Как преодолеть страх перед ОГЭ и ЕГЭ» | гимназия №1 | 11 | Какаджанов Артур Илмуратович | 89141617834 | |||
01. 12:18:54 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ 37 | 9В | Молодцева Алиса Андреевна | +79141968033 | |||
| 01.11.2022 10:30:00 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ №42 | 8А | Глушакова Алина Евгеньевна | 89143169842 | |||
| 01.11.2022 09:21:26 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ с УИОП №16 | 10Б | Сосина Виталина Витальевна | 89147792000 | |||
01. 11.2022 11.202200:15:42 | ИПиП Тренинг «Как преодолеть страх перед ОГЭ и ЕГЭ» | инженерная школа | 9а | Иманрммти мамин иоиит | 89095586522 | |||
| 31.10.2022 23:59:06 | ФФиМК Лингвистическая игра «Тайны фразеологии» | МОУ СОШ С УИОП 16 | 10А | Куберская Полина Степановна | 79242251090 | |||
| 31.10.2022 10:19:05 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МБОУ СОШ 2 с. п. «Село Хурба» п. «Село Хурба» | 11 | Шульга Варвара Викторовна | 89638280255 | |||
| 30.10.2022 20:26:54 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МОУ «Инженерная школа» | 11 А | Болдырев Семен | 89990876239 | |||
| 30.10.2022 20:24:22 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МОУ «Инженерная школа» | 11 А | Рыжкина Александра Викторовна | 89141633647 | |||
| Всего: 183 | ||||||||
Результаты 1 — 10 из 183
Начало | Пред. | 1 2 3 4 5 | След. | Конец | Все
| 1 2 3 4 5 | След. | Конец | Все
Естественный отбор, генетический дрейф и поток генов не действуют изолированно в природных популяциях
Естественный отбор, генетический дрейф и поток генов — это механизмы, вызывающие изменения частот аллелей с течением времени. Когда одна или несколько из этих сил действуют в популяции, популяция нарушает предположения Харди-Вайнберга, и происходит эволюция. Таким образом, теорема Харди-Вайнберга обеспечивает нулевую модель для изучения эволюции, и основное внимание популяционной генетики уделяется пониманию последствий нарушения этих предположений.
Естественный отбор имеет место, когда особи с определенными генотипами имеют больше шансов, чем особи с другими генотипами, выжить и размножиться и, таким образом, передать свои аллели следующему поколению. Как утверждал Чарльз Дарвин (1859 г.) в книге «О происхождении видов» , естественный отбор должен происходить при соблюдении следующих условий:
- Среди особей внутри популяции существуют различия по некоторым признакам.

- Эта вариация является наследственной (т. е. существует генетическая основа вариации, так что потомство имеет тенденцию напоминать своих родителей по этому признаку).
- Изменчивость по этому признаку связана с изменчивостью приспособленности (среднее чистое воспроизводство особей с данным генотипом по отношению к особям с другими генотипами).
Направленный отбор со временем приводит к увеличению частоты предпочтительного аллеля. Рассмотрим три генотипа ( АА , Аа и аа ), которые различаются по приспособленности, так что особи АА производят в среднем больше потомства, чем особи других генотипов. В этом случае, если предположить, что селективный режим остается постоянным и что действие отбора является единственным нарушением предположений Харди-Вайнберга, Аллель становился все более распространенным с каждым поколением и в конечном итоге закреплялся в популяции. Скорость, с которой полезный аллель приближается к фиксации, частично зависит от доминантных отношений между аллелями в рассматриваемом локусе (Рис.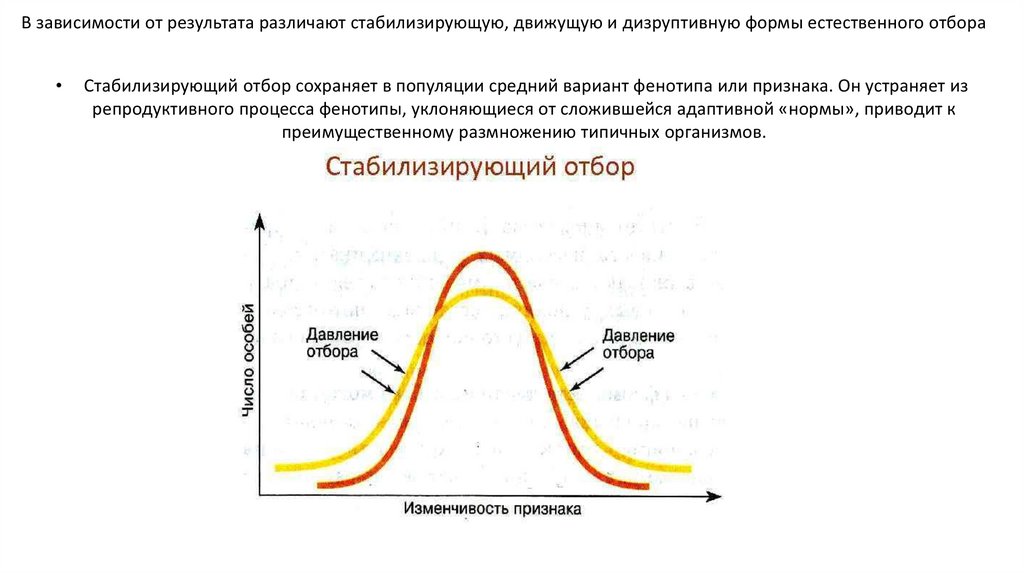

Рисунок 1: Изменение частоты аллелей при направленном отборе в пользу (а) доминантного полезного аллеля и (б) рецессивного благоприятного аллеля
Балансирующий отбор, в отличие от направленного отбора, поддерживает генетический полиморфизм в популяциях. Например, если гетерозиготы в локусе имеют более высокую приспособленность, чем гомозиготы (сценарий, известный как преимущество или сверхдоминирование гетерозигот), естественный отбор будет поддерживать множественные аллели на стабильных равновесных частотах. Стабильный полиморфизм также может сохраняться в популяции, если приспособленность, связанная с генотипом, снижается по мере увеличения частоты этого генотипа (т. е. если существует отрицательный частотно-зависимый отбор). Важно отметить, что недостаток гетерозигот (недостаточное доминирование) и положительный частотно-зависимый отбор также могут действовать на локус, но ни один из них не поддерживает множественные аллели в популяции, и, таким образом, ни один из них не является формой уравновешивающего отбора.
Генетический дрейф возникает из-за ошибки выборки, присущей передаче гамет особями в ограниченной популяции. Пул гамет популяции в поколении t — это общий пул яйцеклеток и сперматозоидов, произведенных особями в этом поколении. Если бы пул гамет был бесконечен по размеру и если бы не было отбора или мутации, действующих в локусе с двумя аллелями ( А и А ), мы могли бы ожидать, что доля гамет, содержащих аллель А, точно равна частоте из A , а доля гамет, содержащих a , равна частоте a . Сравните эту ситуацию с подбрасыванием правильной монеты. Если бы вы подбрасывали монету бесконечное количество раз, доля выпадения орла была бы 0,50, а доля решки — 0,50. Однако если вы подбросите монету всего 10 раз, не стоит слишком удивляться, если выпадет 7 орлов и 3 решки. Это отклонение от ожидаемых частот начала и окончания связано с ошибкой выборки. Чем больше раз вы подбрасываете монету, тем ближе эти частоты должны приближаться к 0,50, поскольку ошибка выборки уменьшается по мере увеличения размера выборки.
В конечной популяции взрослые особи в поколении t передадут конечное число гамет, чтобы произвести потомство в поколении t + 1. Частоты аллелей в этом пуле гамет обычно отклоняются от частот популяции в поколение t из-за ошибки выборки (опять же, если предположить, что в локусе нет выбора). Таким образом, частоты аллелей в этой популяции со временем будут меняться из-за случайных событий, то есть популяция будет подвергаться генетическому дрейфу. Чем меньше численность населения ( N ), тем важнее эффект генетического дрейфа. На практике при моделировании эффектов дрейфа мы должны учитывать эффективный размер популяции ( N e ), который по существу представляет собой количество размножающихся особей и может отличаться от учетного размера, N , при различных сценариях. включая неравное соотношение полов, определенные структуры спаривания и временные колебания численности популяции.
В локусе с множественными нейтральными аллелями (аллелями, идентичными по своему влиянию на приспособленность) генетический дрейф приводит к фиксации одного из аллелей в популяции и, таким образом, к потере других аллелей, так что гетерозиготность в популяции спадает до нуля. В любой момент времени вероятность того, что один из этих нейтральных аллелей в конечном итоге закрепится, равна частоте этого аллеля в популяции. Мы можем рассматривать эту проблему с точки зрения множества повторяющихся популяций, каждая из которых представляет дему (субпопуляцию) внутри метапопуляции (набора дем). Учитывая 10 конечных демов, равных N e , каждая из которых имеет начальную частоту аллеля A , равную 0,5, мы ожидаем возможную фиксацию A в 5 демах и возможную потерю A в 5 демах. Наши наблюдения, вероятно, в некоторой степени отклоняются от этих ожиданий, поскольку мы рассматриваем конечное число демов (рис. 2). Таким образом, дрейф генов устраняет генетическую изменчивость внутри демов, но приводит к дифференциации между демами полностью за счет случайных изменений частот аллелей.
В любой момент времени вероятность того, что один из этих нейтральных аллелей в конечном итоге закрепится, равна частоте этого аллеля в популяции. Мы можем рассматривать эту проблему с точки зрения множества повторяющихся популяций, каждая из которых представляет дему (субпопуляцию) внутри метапопуляции (набора дем). Учитывая 10 конечных демов, равных N e , каждая из которых имеет начальную частоту аллеля A , равную 0,5, мы ожидаем возможную фиксацию A в 5 демах и возможную потерю A в 5 демах. Наши наблюдения, вероятно, в некоторой степени отклоняются от этих ожиданий, поскольку мы рассматриваем конечное число демов (рис. 2). Таким образом, дрейф генов устраняет генетическую изменчивость внутри демов, но приводит к дифференциации между демами полностью за счет случайных изменений частот аллелей.
Рисунок 2: Моделирование изменения частоты аллеля в 10 повторных популяциях (N = 20)
Поскольку начальная частота аллеля A = 0,5, мы ожидаем, что A будет зафиксирован в 5 популяциях и потерян в 5 популяциях, но наши наблюдения отклоняются от ожиданий из-за конечного числа популяций.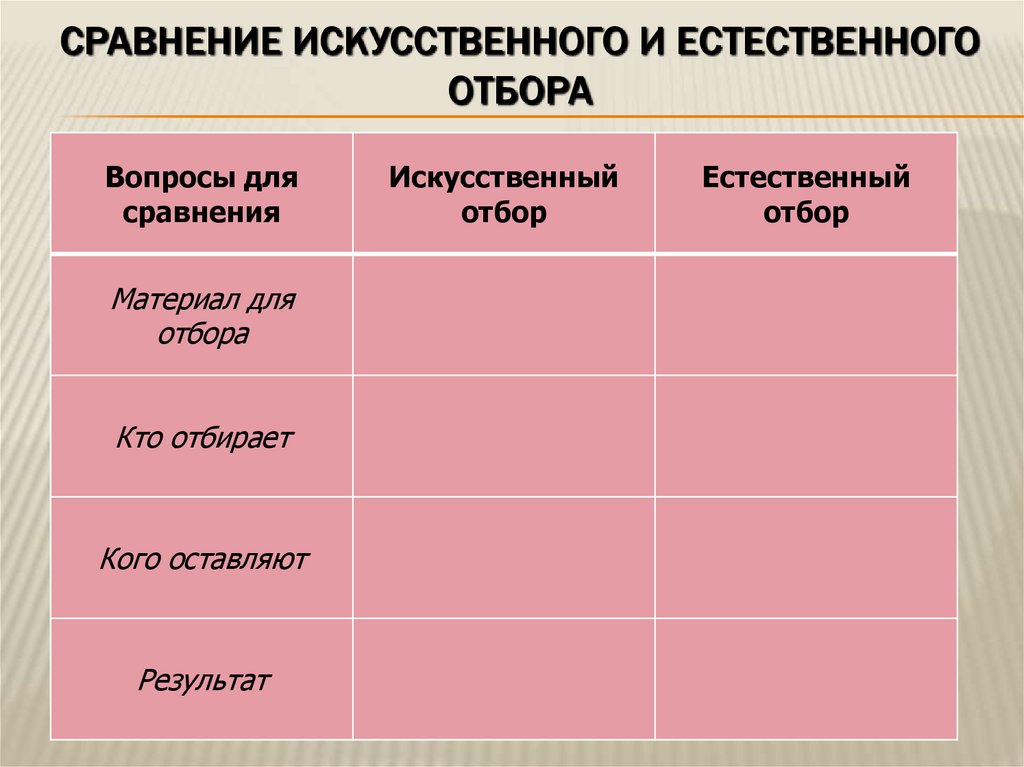 В этом цикле симуляций мы видим 7 случаев фиксации (p = 1), 2 случая потери (p = 0) и один случай, в котором остаются два аллеля после 100 поколений. В этой последней популяции A в конечном итоге достигнет фиксации или потери.
В этом цикле симуляций мы видим 7 случаев фиксации (p = 1), 2 случая потери (p = 0) и один случай, в котором остаются два аллеля после 100 поколений. В этой последней популяции A в конечном итоге достигнет фиксации или потери.
Поток генов — это перемещение генов в популяцию или из нее. Такое перемещение может быть связано с миграцией отдельных организмов, размножающихся в своих новых популяциях, или с перемещением гамет (например, вследствие переноса пыльцы между растениями). В отсутствие естественного отбора и генетического дрейфа поток генов приводит к генетической однородности среди демов в метапопуляции, так что для данного локуса частоты аллелей достигают равновесных значений, равных средним частотам в метапопуляции. Напротив, ограниченный поток генов способствует дивергенции популяций за счет отбора и дрейфа, которые, если они сохраняются, могут привести к видообразованию.
Естественный отбор, генетический дрейф и поток генов не действуют изолированно, поэтому мы должны рассмотреть, как взаимодействие этих механизмов влияет на эволюционные траектории в природных популяциях.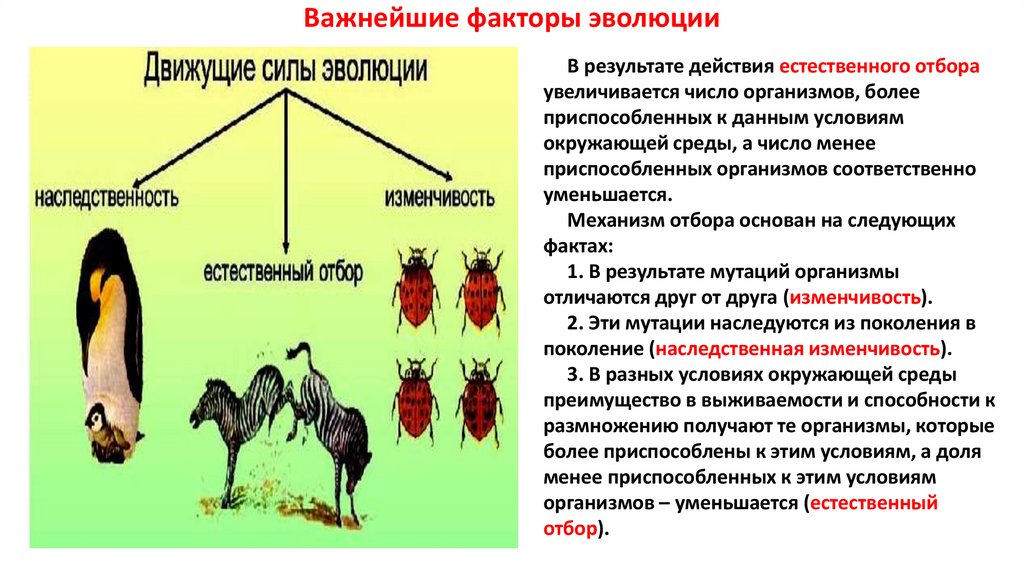 Этот вопрос имеет решающее значение для специалистов по природоохранной генетике, которые борются с последствиями этих эволюционных процессов, разрабатывая заповедники и моделируя динамику популяций находящихся под угрозой исчезновения видов в фрагментированных средах обитания. Все реальные популяции конечны и, следовательно, подвержены влиянию генетического дрейфа. Мы ожидаем, что в бесконечной популяции направленный отбор в конце концов зафиксирует выгодный аллель, но это не обязательно произойдет в конечной популяции, потому что эффекты дрейфа могут преодолеть эффекты отбора, если отбор слаб и/или популяция мала. Потеря генетической изменчивости из-за дрейфа вызывает особую озабоченность в небольших популяциях, находящихся под угрозой исчезновения, в которых фиксация вредных аллелей может снизить жизнеспособность популяции и повысить риск исчезновения. Даже если усилия по сохранению увеличат рост популяции, низкая гетерозиготность, вероятно, сохранится, поскольку узкие места (периоды сокращения численности популяции) оказывают более выраженное влияние на Ne, чем периоды большей численности популяции.
Этот вопрос имеет решающее значение для специалистов по природоохранной генетике, которые борются с последствиями этих эволюционных процессов, разрабатывая заповедники и моделируя динамику популяций находящихся под угрозой исчезновения видов в фрагментированных средах обитания. Все реальные популяции конечны и, следовательно, подвержены влиянию генетического дрейфа. Мы ожидаем, что в бесконечной популяции направленный отбор в конце концов зафиксирует выгодный аллель, но это не обязательно произойдет в конечной популяции, потому что эффекты дрейфа могут преодолеть эффекты отбора, если отбор слаб и/или популяция мала. Потеря генетической изменчивости из-за дрейфа вызывает особую озабоченность в небольших популяциях, находящихся под угрозой исчезновения, в которых фиксация вредных аллелей может снизить жизнеспособность популяции и повысить риск исчезновения. Даже если усилия по сохранению увеличат рост популяции, низкая гетерозиготность, вероятно, сохранится, поскольку узкие места (периоды сокращения численности популяции) оказывают более выраженное влияние на Ne, чем периоды большей численности популяции.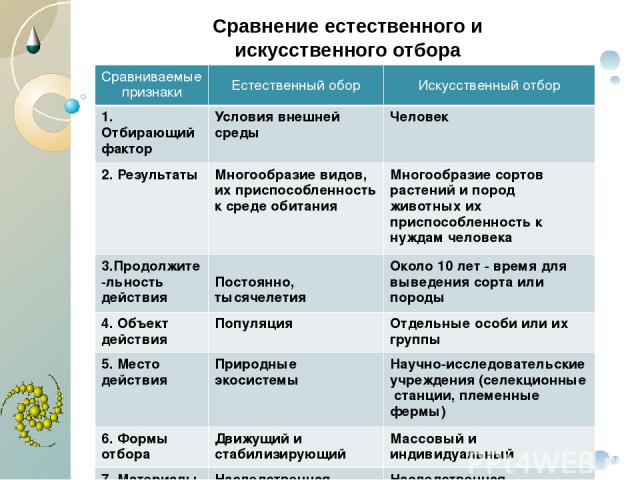
Мы уже видели, что генетический дрейф ведет к дифференциации демов внутри метапопуляции. Если принять простую модель, в которой индивидуумы имеют равные вероятности расселения среди всех демов (каждый из которых имеет эффективный размер N и ) внутри метапопуляции, то скорость миграции ( м ) представляет собой долю копий гена в пределах дема, введенная через иммиграцию в поколение. Согласно общепринятому приближению, интродукция только одного мигранта на поколение ( N e m = 1) составляет достаточный поток генов, чтобы противодействовать диверсифицирующим эффектам генетического дрейфа в метапопуляции.
Естественный отбор может производить генетическую изменчивость среди демов в пределах метапопуляции, если в разных демах преобладает различное давление отбора. Если N e достаточно велико, чтобы не учитывать влияние генетического дрейфа, то мы ожидаем, что направленный отбор закрепит предпочтительный аллель в пределах данной фокальной демы. Однако постоянное введение посредством генного потока аллелей, полезных в других демах, но вредных в фокальном деме, может противодействовать эффектам отбора. В этом сценарии вредоносный аллель останется на промежуточной равновесной частоте, которая отражает баланс между потоком генов и естественным отбором.
Однако постоянное введение посредством генного потока аллелей, полезных в других демах, но вредных в фокальном деме, может противодействовать эффектам отбора. В этом сценарии вредоносный аллель останется на промежуточной равновесной частоте, которая отражает баланс между потоком генов и естественным отбором.
Аргумент Дарвина
Аргумент ДарвинаТеория Чарльза Дарвина эволюции по естественному отбору
Объяснение очевидного дизайн встречается в природе.
- Организмы различаются, и эти случайные, но небольшие вариации наследуются ,
по крайней мере частично, их потомками (которые напоминают своих родителей в этих
характеристики).
- Организмы производят больше потомства, чем могут выжить , поскольку
ресурсы жизнеобеспечения ограничены. В этой борьбе за существование многие
организмы погибают, но некоторые выживают и размножаются. (Мальтузианский
принцип)
- Потомство, отличающееся тем, что дает ему преимущество в
среда выживет и размножится более успешно , в среднем,
чем потомство, которое менее приспособлено. Выжившие обычно лучше подходят
чем невыжившие.
- Таким образом, в популяциях накапливаются благоприятные вариации . (Такой
вариации повышают ПРИГОДНОСТЬ* или вероятность того, что организм выживет
и размножаются в данной среде.) И наоборот, неблагоприятные вариации
ухудшит физическую форму и, скорее всего, приведет к гибели их носителей.
Назовите этот неслучайный процесс дифференцированного выживания и размножения, основанный на
на условные экологические обстоятельства ЕСТЕСТВЕННЫЙ ОТБОР .
- Таким образом, организмов и их комплекс признаков (т. е. АДАПТАЦИИ =
наследственные вариации, повышающие приспособленность) будет отображаться как разработанный или
хорошо приспособлены к местным условиям и образу жизни. Этот результат, а именно
очевидный замысел в природе, не требует сверхъестественного вмешательства, но также
не исключает этого.
 (Мальтузианский
принцип)
(Мальтузианский
принцип) 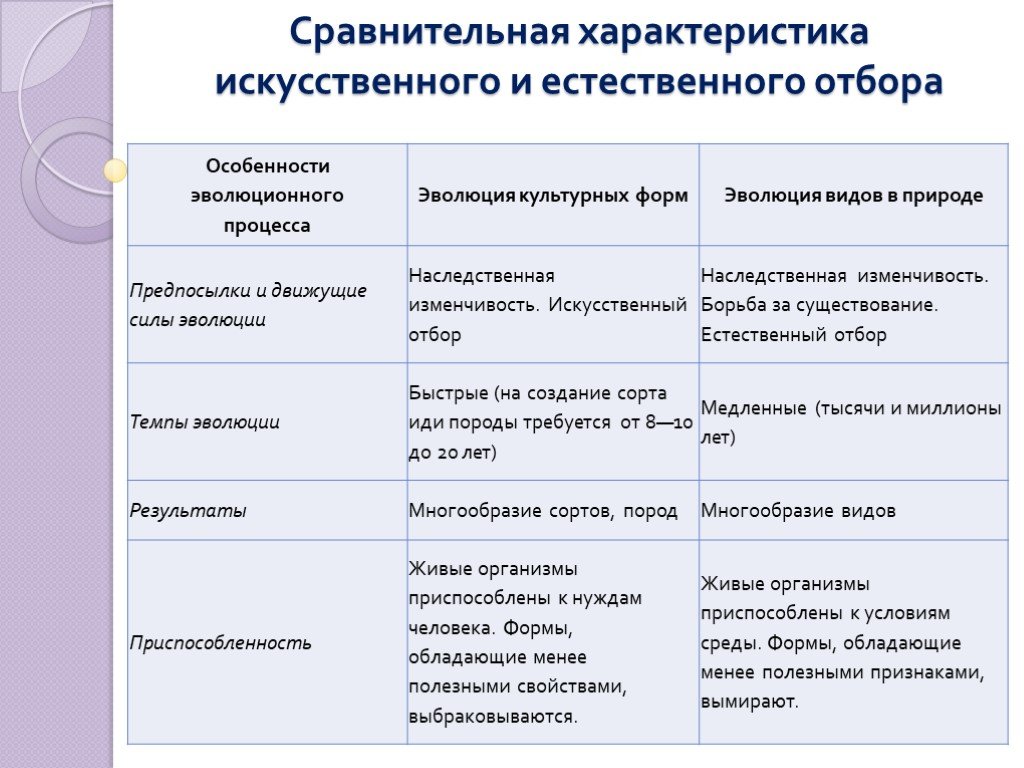 Этот результат, а именно
очевидный замысел в природе, не требует сверхъестественного вмешательства, но также
не исключает этого.
Этот результат, а именно
очевидный замысел в природе, не требует сверхъестественного вмешательства, но также
не исключает этого.Дарвин описал его взгляд в 1859 г.как теория происхождения с модификацией через изменение и естественный отбор . Обратите внимание, что теория Дарвина (и современная теория органической эволюции) не предполагает и не подразумевает, что естественный отбор должен происходить. Этот процесс случайного изменения и неслучайного отбора (который наблюдалось спонтанно) — это только один из возможных естественных механизмов эволюционных изменений, но есть и другие (например, мутация, миграция, генетическая дрейф**). Однако дарвинисты считают, что этот процесс является первичным механизмом эволюции, хотя многие биологи-эволюционисты с этим не согласны. Естественный отбор является альтернативой сверхъестественному [вызванному богом] или искусственному [вызванному человеком] выбор.
«Это интересно созерцать запутанный берег, покрытый множеством растений разных видов, с пением птиц на кустах, с порхающими разными насекомыми вокруг, и с червями, ползающими по сырой земле, и отражать, что эти искусно построенные формы, столь отличные друг от друга и зависимые друг на друга таким сложным образом, все они были произведены законами, действующими вокруг нас.
Чарльз Дарвин ( О происхождении видов путем естественного отбора или сохранения избранных рас в борьбе за жизнь, 1859, XIV:489)
- Весь последняя глава «Происхождения», в которой Дарвин повторяет свой основной аргумент и заключение
- Дарвина Метафора Древа Жизни (Калифорнийский университет в Беркли)
- Некоторые из опубликованных работ Дарвина
- Дарвин об органах предельного совершенства и сложности
- Введение к эволюционной биологии (Крис Колби)
- Эволюция и философия (Джон Уилкинс)
- 29 Доказательства макроэволюции (Дуглас Теобальд)
- Кольцо эволюции
- Эволюция Обновлять
- Как близки ли люди к человекообразным обезьянам и другим животным?
- Эволюция и происхождение болезни
- Наблюдается Примеры видообразования
*»Дано в конкретной среде каждый генетический тип в популяции имеет некоторую вероятность выживания и размножения, то, что мы называем приспособленностью типа .
** Генетический дрейф — это случайное колебание частоты примера гена в популяции, независимо от наследственности или приспособленности. Возникает дрейф, например, когда гены случайно удаляются из популяции в результате природной катастрофы. Когда случайные события вызывают последовательные изменения в частоты двух или более генов в небольших предковых популяциях производят значительные кумулятивные различия в частотах генов у последующих потомков популяции, распределение генов, таким образом, случайным образом дрейфует от поколения к поколению.
 Эти законы, взятые в самом широком смысле, суть рост с воспроизводством;
Наследование, которое почти подразумевает воспроизводство; Изменчивость от
косвенное и прямое действие условий жизни и от использования и неиспользования:
Коэффициент увеличения настолько высок, что приводит к борьбе за жизнь и, как следствие,
к естественному отбору, влекущему за собой дивергенцию признаков и вымирание
менее совершенных форм. Так от войны природы, от голода и смерти,
самый возвышенный объект, который мы способны себе представить, а именно производство
из высших животных следует непосредственно. В этом взгляде есть величие
жизнь с ее различными силами, первоначально вдохнутая в несколько
формы или в одну; и что, в то время как эта планета вращалась в соответствии с
к неизменному закону тяготения, от такого простого начала бесконечные формы наиболее
прекрасные и самые замечательные были и находятся в процессе эволюции» 9.0121
Эти законы, взятые в самом широком смысле, суть рост с воспроизводством;
Наследование, которое почти подразумевает воспроизводство; Изменчивость от
косвенное и прямое действие условий жизни и от использования и неиспользования:
Коэффициент увеличения настолько высок, что приводит к борьбе за жизнь и, как следствие,
к естественному отбору, влекущему за собой дивергенцию признаков и вымирание
менее совершенных форм. Так от войны природы, от голода и смерти,
самый возвышенный объект, который мы способны себе представить, а именно производство
из высших животных следует непосредственно. В этом взгляде есть величие
жизнь с ее различными силами, первоначально вдохнутая в несколько
формы или в одну; и что, в то время как эта планета вращалась в соответствии с
к неизменному закону тяготения, от такого простого начала бесконечные формы наиболее
прекрасные и самые замечательные были и находятся в процессе эволюции» 9.0121 Эти приспособленности можно усреднить, взвесив каждую по частоте генетических изменений.
находится в популяции, чтобы получить число, называемое средней приспособленностью
население. Эволюция – это изменение частот различных
типов от поколения к поколению, в результате чего средняя пригодность
населения меняется от поколения к поколению» (Ричард Левонтин,
1998).
Эти приспособленности можно усреднить, взвесив каждую по частоте генетических изменений.
находится в популяции, чтобы получить число, называемое средней приспособленностью
население. Эволюция – это изменение частот различных
типов от поколения к поколению, в результате чего средняя пригодность
населения меняется от поколения к поколению» (Ричард Левонтин,
1998).