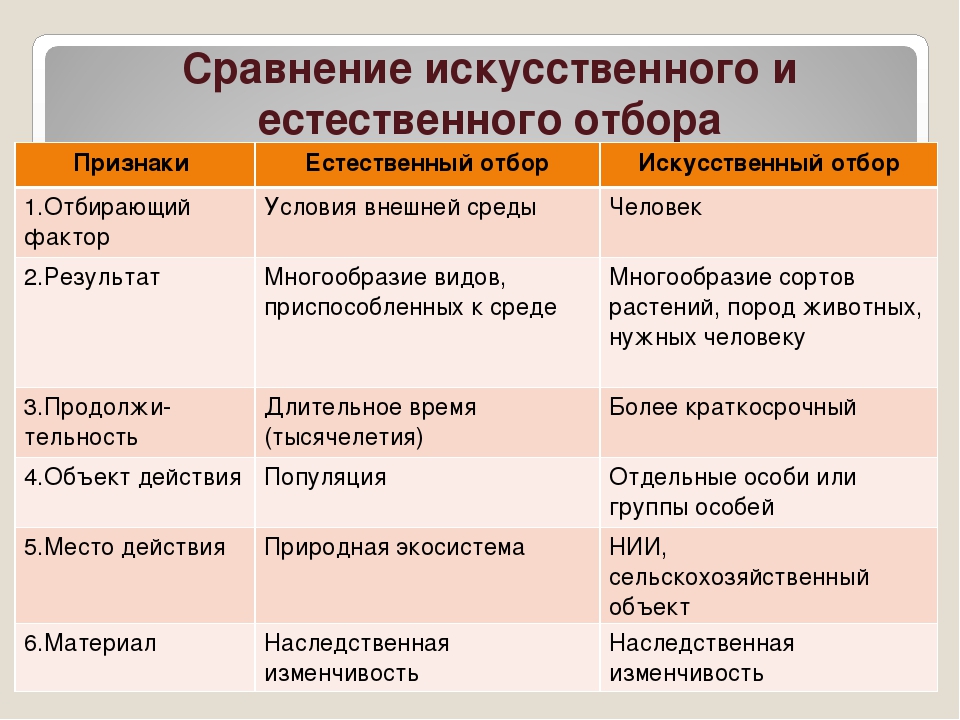
Естественный отбор. Большая российская энциклопедия
Есте́ственный отбо́р, процесс избирательного выживания и дифференциального размножения организмов, основной движущий фактор их эволюции. Естественный отбор носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции.
Историческая справка
Идеи о существовании естественного отбора высказывались с начала 19 в. разными английскими натуралистами (в том числе А. Уоллесом). Но только Ч. Дарвин (1842, 1859) оценил его как главный фактор эволюции. По Дарвину, естественный отбор является результатом борьбы за существование; даже незначительные наследуемые различия между особями одного вида могут дать преимущества в этой борьбе, которая обусловлена тенденцией организмов к высокой интенсивности размножения (в геометрической прогрессии) и невозможности сохранения всего потомства вследствие ограниченности природных ресурсов. Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к естественному отбору – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение многих поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии естественного отбора Дарвин строил преимущественно на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции естественный отбор определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к естественному отбору – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение многих поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии естественного отбора Дарвин строил преимущественно на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции естественный отбор определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Систематическое исследование естественного отбора, расширение и совершенствование методик его изучения началось с конца 19 в. Использование методов биометрии позволило установить статистически значимые различия между выжившими и погибшими организмами при изменении условий среды. Благодаря разработкам Р. Фишера, Дж. Холдейна, С. Райта и С. С. Четверикова, осуществивших синтез классического дарвинизма и генетики, появилась возможность приступить к экспериментальному изучению генетических основ естественного отбора. Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с другими мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетическую разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность естественного отбора. Кроме того, стало очевидным, что естественный отбор имеет дело не с единичными признаками, а с целыми организмами и что генетическая сущность естественного отбора заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям.
Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с другими мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетическую разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность естественного отбора. Кроме того, стало очевидным, что естественный отбор имеет дело не с единичными признаками, а с целыми организмами и что генетическая сущность естественного отбора заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям.
Роль естественного отбора в эволюционном процессе
Естественный отбор носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции. Под контролем естественного отбора находятся не только разнообразные признаки, но и сами факторы эволюции, например интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же естественного отбора происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетического груза, в том числе в популяциях современного человека.
Под контролем естественного отбора находятся не только разнообразные признаки, но и сами факторы эволюции, например интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же естественного отбора происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетического груза, в том числе в популяциях современного человека.
Формы естественного отбора
Выделяют более 30 форм естественного отбора; ни одна из них не существует в чистом виде, а скорее характеризует тенденцию действия отбора в конкретной экологической ситуации. Так, движущий отбор способствует сохранению определённого отклонения от прежней нормы и приводит к выработке новых приспособлений через направленную перестройку всего генофонда популяций, а также генотипов и фенотипов особей. Он может вести к доминированию одной (или нескольких) ранее существовавшей формы над другими. Классическим примером его действия стало преобладание в промышленных районах темноокрашенных форм бабочки берёзовая пяденица, незаметных для птиц на загрязнённых копотью стволах деревьев (до середины 19 в. встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание различных видов насекомых и грызунов к ядам, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Например, длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание различных видов насекомых и грызунов к ядам, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Например, длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
Результатом действия движущего отбора в филогенезе крупных таксонов является ортоселекция, примером которой служит установленная В. О. Ковалевским направленная эволюция конечности предков лошади (от пятипалости к однопалости), протекавшая миллионы лет и обеспечившая увеличение скорости и экономичности бега.
Дизруптивный, или разрывающий, отбор благоприятствует сохранению крайних отклонений и ведёт к увеличению полиморфизма. Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в начале 20 в. российский ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю – низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской.
Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в начале 20 в. российский ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю – низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской. В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классический пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классический пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
Если указанные формы естественного отбора приводят к изменению и фенотипической, и генетической структуры популяций, то впервые описанный И. И. Шмальгаузеном (1938) стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение геномы особей, наиболее отклоняющихся от этой нормы. Он направлен на поддержание и повышение устойчивости в популяции среднего, ранее сложившегося фенотипа. Известно, например, что во время снежных бурь выживают птицы, которые по многим признакам (длина крыла, клюва, масса тела и т. д.) приближаются к средней норме, а отклонившиеся от этой нормы особи погибают.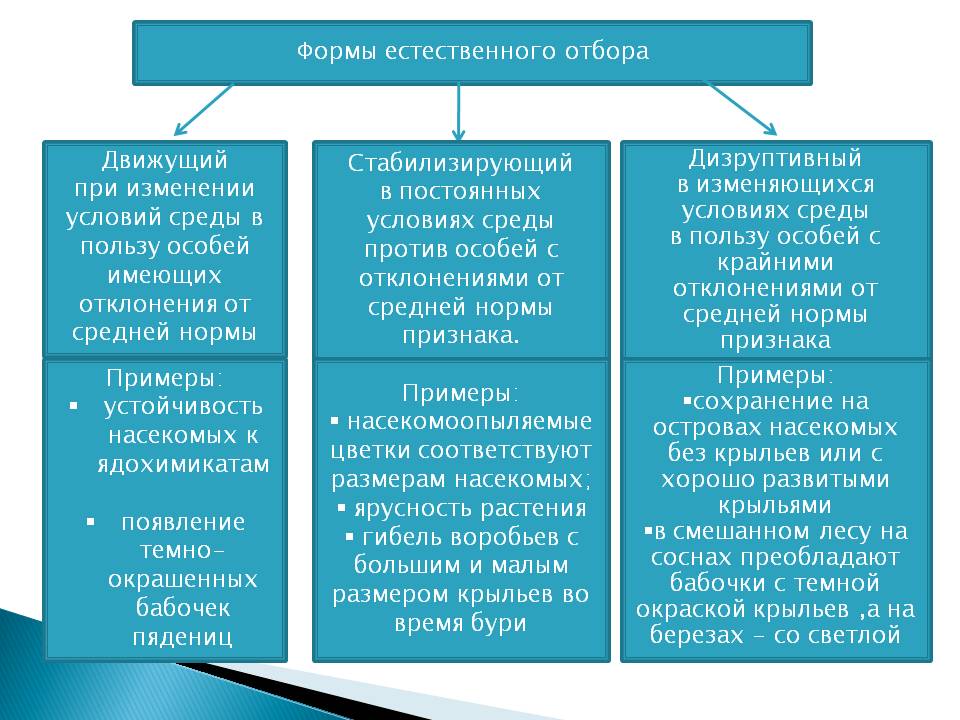 Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (например, шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком). Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетические изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (например, шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком). Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетические изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Дестабилизирующий отбор (название предложено Д. К. Беляевым, 1970) ведёт к резкому нарушению систем регуляции онтогенеза, вскрытию мобилизационного резерва и росту фенотипической изменчивости при интенсивном отборе в каком-либо определённом направлении. Например, отбор на снижение агрессивности хищных зверей в неволе через перестройку нейрогуморальной системы ведёт к дестабилизации цикла размножения, сдвигам в сроках линьки, изменениям в положении хвоста, ушей, в окраске и др.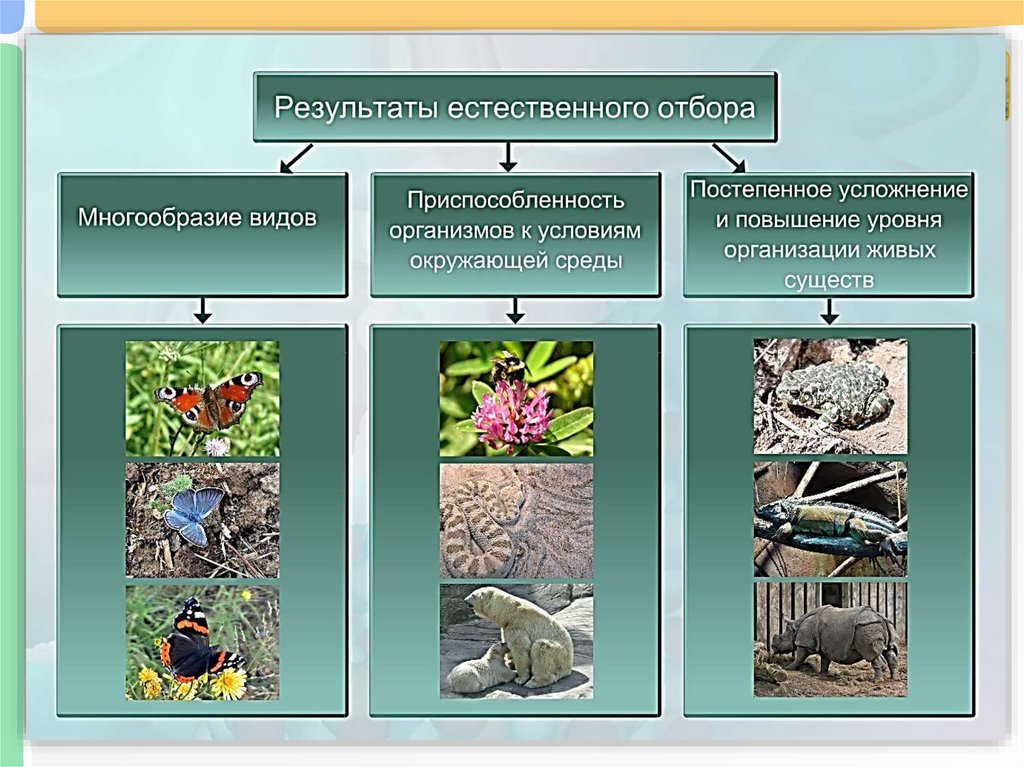
Обнаружены гены, которые могут быть летальными или снижать жизнеспособность организмов в гомозиготном состоянии, а в гетерозиготном, напротив, повышать экологическую пластичность и другие показатели. В этом случае можно говорить о так называемом сбалансированном отборе, обеспечивающем поддержание генетического разнообразия с определённым соотношением частот аллелей. Примером его действия может служить повышение устойчивости у больных серповидно-клеточной анемией (гетерозиготных по гену гемоглобина S) к заражению различными штаммами малярийного плазмодия.
Важным шагом в преодолении стремления все признаки организмов объяснять действием естественного отбора стала концепция нейтральной эволюции, согласно которой часть изменений на уровне белков и нуклеиновых кислот происходит путём фиксации адаптивно нейтральных или почти нейтральных мутаций. Возможен отбор видов, возникающих в периферийных популяциях «внезапно» с геохронологической точки зрения.
Ещё раньше было доказано, что катастрофический отбор, при котором в период резких изменений среды выживает небольшое число особей и даже единственный организм, может стать основой формирования нового вида за счёт хромосомной перестройки и смены экологической ниши. Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофический характер в периферийных популяциях.
Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофический характер в периферийных популяциях.
Естественный отбор, затрагивающий вторичные половые признаки особей, называют половым (например, яркая брачная окраска самцов у многих видов рыб и птиц, зазывающие крики, специфические запахи, сильно развитые орудия для турнирного боя у млекопитающих). Эти признаки полезны, так как повышают возможность участия их носителей в воспроизведении потомства. В половом отборе наибольшую активность проявляют самцы, что выгодно для вида в целом, так как самки остаются в большей безопасности в период размножения.
Выделяют также групповой отбор, способствующий сохранению признаков, полезных семье, стае, колонии. Его частным случаем у колониальных насекомых является отбор сородичей, при котором стерильные касты (рабочие, солдаты и др.) обеспечивают (нередко ценой собственной жизни) выживание плодовитых особей (маток) и личинок и тем самым сохранение всей колонии. Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Критика существующих концепций
Хотя представления о ведущей роли естественного отбора в эволюции получили подтверждение во множестве экспериментов, до сих пор они подвергаются критике, исходящей из представления о невозможности образования организмов в результате случайной комбинации мутаций. При этом игнорируется тот факт, что каждый акт естественного отбора совершается на базе предыдущих результатов его же действия, которые, в свою очередь, предопределяют формы, интенсивность и направления естественного отбора, а значит, пути и закономерности эволюции.
Колчинский Эдуард ИзраилевичДата публикации: 10 июня 2022 г. в 11:17 (GMT+3)
Естественный отбор начинается на уровне генов
В последнее десятилетие среди ученых все больше распространяется недоверие к некоторым положениям эволюционной теории, сложившейся в середине XX века.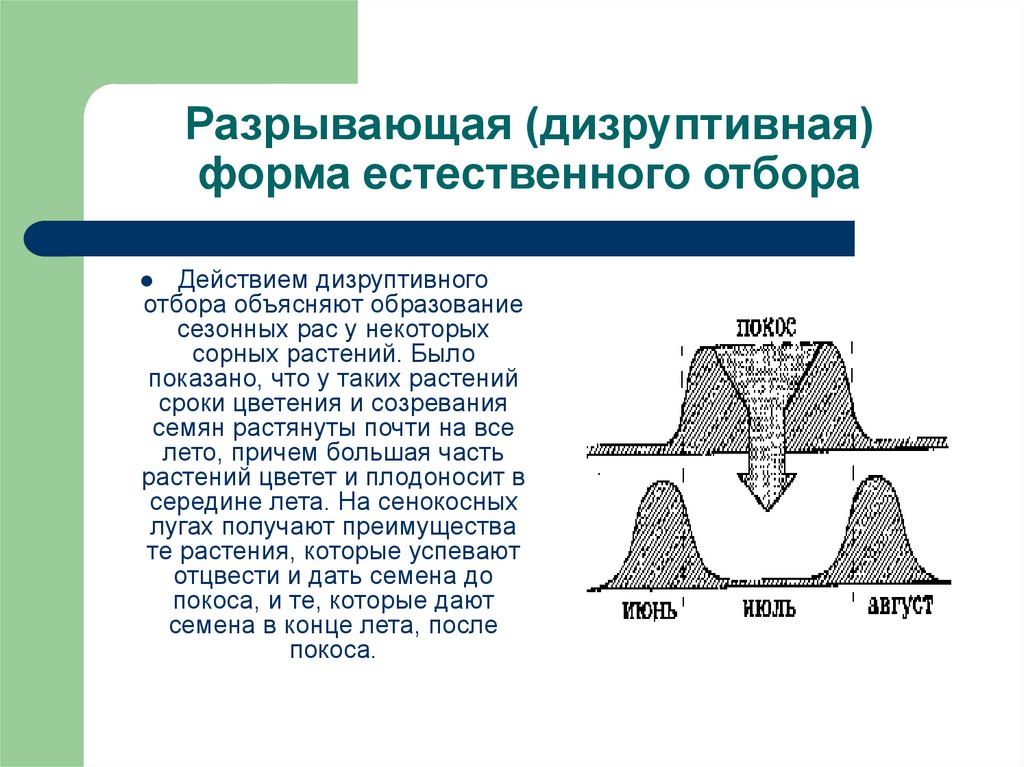 Особенно много сомнений вызывает тезис о полной случайности всех наследственных изменений. Возможно, мутации совсем не случайны.
Особенно много сомнений вызывает тезис о полной случайности всех наследственных изменений. Возможно, мутации совсем не случайны.
В 1950-1960-ые годы XX века, когда сложилась так называемая синтетическая теория эволюции, произошло своего рода объединение классического дарвинизма XIX века с достижениями генетики и молекулярной биологии. В синтетической теории эволюции принималось, что основными факторами эволюции являются мутационный процесс, то есть возникновение случайных изменений в ДНК и естественный отбор. При этом считалось, что наследственные изменения имеют случайный характер. Но в дальнейшем накопилось довольно много фактов, которые говорят о том что, по-видимому, далеко не всегда изменения генома имеют случайный характер. И поэтому давний спор о том, как же идет эволюция – на основе случайностей или на основе закономерностей, продолжается и по сей день.
Член-корреспондент Российской Академии медицинских наук, сотрудник института микробиологии и эпидемиологии имени Гамалеи Георгий Смирнов рассказывает о новом видении классической триады дарвинизма — наследственности, изменчивости и естественного отбора.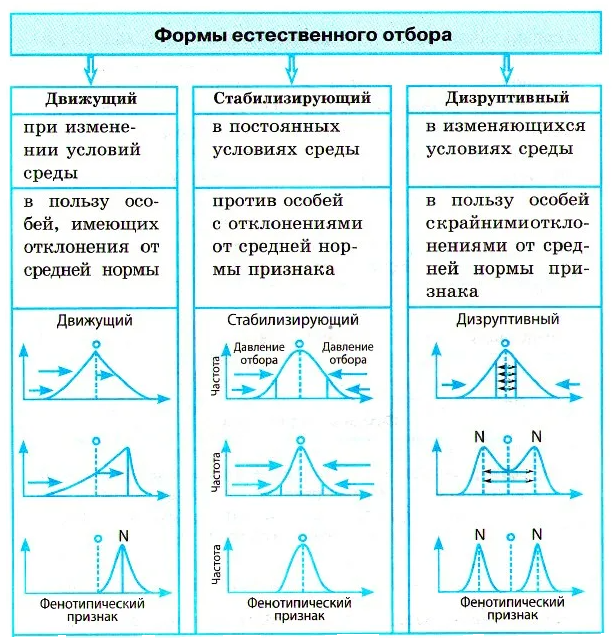
Наследственность, изменчивость и естественный отбор
Мутация (изменчивость) и естественный отбор, безусловно, всегда считались и до сих пор признаются важнейшим фактором эволюции, но есть еще наследственность. То есть эволюция – это наследственность, изменчивость и естественный отбор. Чтобы происходили мутации, нужно, чтобы было где им происходить. И представлена эта наследственность полимерной молекулой ДНК, в которой записаны свойства любого организма и записаны вполне определенным образом с помощью единого генетического кода. Вся совокупность генов, которые определяют свойства организмов, называется геномом, и эволюция любого вида, любого организма начинается с изменения его собственного генома. До сих пор считается, что тот или иной ген может закрепиться и распространиться в популяциях в тех случаях, когда этот ген кодирует какой-то полезный для того или иного организма признак. И собственно естественный отбор призван отбирать те признаки, которые в данных условиях являются полезными.
Но прежде чем в естественном отборе победит тот или иной вид, необходимо чтобы победил ген, чтобы он закрепился в ДНК особи этого вида и распространился. Все начинается с гена. Дело в том, что гены и наследуются, и отбрасываются не сами по себе. Они отбрасываются и наследуются в составе неких сегментов ДНК. Когда ген отбрасывается, выпадает из хромосомы, это происходит в два этапа. Сначала ген активируется за счет мутации, а потом сегмент ДНК, содержащий поврежденный ген, вырезается. То есть удаляется сегмент, содержащий поврежденный ген, который не поддерживается естественным отбором.
ДНК – это текст
Теперь я хочу перейти к тому, почему это происходит. Причем здесь уместно рассматривать оба процесса, и процесс внедрения информации в ДНК, и процесс выбрасывания информации из ДНК. Эти противоположно направленные процессы, в общем-то имеют очень близкую сущность. Информация извне не может попасть в любой отрезок ДНК.
ДНК – это текст. Мы с вами прекрасно понимаем, что если взять осмысленный текст и взять фрагмент другого текста, то мы не можем произвольно вставить этот чужеродный фрагмент в любое место исходного текста. Это приведет к нарушению смысла и, конечно, будет заметно. Правда, в словесных текстах можно допустить внедрение «сорных» слов, например, «так сказать», это словосочетание можно вставить практически в любое место. Так же точно в ДНК можно вставить некую информацию, которая похожа на сорные слова и эта информация называется инсерционным (insertion) элементом, их можно вставить почти в любое место ДНК.
Это приведет к нарушению смысла и, конечно, будет заметно. Правда, в словесных текстах можно допустить внедрение «сорных» слов, например, «так сказать», это словосочетание можно вставить практически в любое место. Так же точно в ДНК можно вставить некую информацию, которая похожа на сорные слова и эта информация называется инсерционным (insertion) элементом, их можно вставить почти в любое место ДНК.
Как только инсерционные элементы оказались в ДНК, они имеют тенденцию себя копировать и встраиваться в новые места той же ДНК, а иногда, если есть такая возможность, то переместиться и в другой организм – это тоже бывает. Как только они оказались в достаточном количестве в ДНК, такая ДНК теряет стабильность. И отрезки ДНК, заключенные между инсерционными элементами, могут либо выбрасываться, либо менять свое местоположение, перемещаться по геному.
Такой элемент ничего не кодирует, кроме собственной способности к перемещению. Это –молекулярный паразит. Свойствами таких элементов обладают многие вирусы, которые внедряются в ДНК или их разнообразные остатки, которые присутствуют в огромном количестве в человеческом геноме. Именно инсерционные элементы, повторяясь, создают нестабильность генома.
Именно инсерционные элементы, повторяясь, создают нестабильность генома.
Естественный отбор на уровне ДНК
Именно инсерционные элементы создают мишени в ДНК, куда может встроиться внешний фрагмент. Второй участник событий – это сам фрагмент, который встраивается, и который может содержать работающий ген. И третий участник событий – это фермент, который узнает и мишень, и встраивающийся фрагмент. Фермент должен распознать и первую и второго и осуществить их объединение, то есть встраивание. Таким образом, есть три компонента, которые необходимы для того, чтобы какой-то сегмент ДНК встроился в геном.
С помощью этого белкового фермента полинуклеотиды в мишени, в месте встраивания выбирают те полинуклеотиды, которые могут в это место встроиться. Я назвал это полинуклеотидным выбором.
Процесс встраивания, в отличие от общепринятого естественного отбора, никак не зависит от внешней среды, все происходит внутри клетки и все определяется свойствами самих полинуклеотидов и фермента, который их распознает.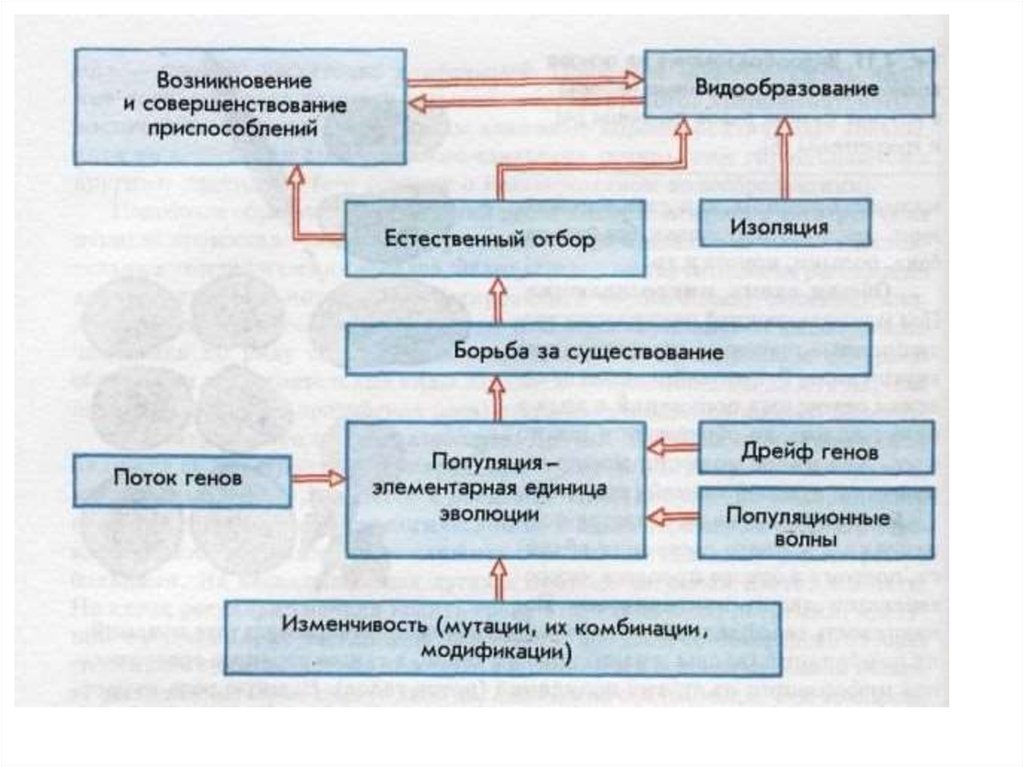
Для того чтобы сегмент ДНК встроился в геном, классический естественный отбор совершенно не нужен. Поэтому я бы разделил естественный отбор на два этапа. Первый этап – это полинуклеотидный выбор, о котором я только что рассказал, а второй этап – это фенотипический отбор, то есть то, что раньше называлось естественным отбором.
Мы видим, что первый акт этой драмы происходит независимо от внешней среды и разыгрывается на уровне общения между собой реципиентного (примимающего) полинуклеотида и донорского (отдающего) полинуклеотида, который входит в геном. А второй акт – это уже закрепление того признака, который был унаследован, и его сохранение и распространение в популяциях.
Полезная информация может быть отброшена
Признак может быть не только закреплен, но и отсеян. Причем отсеивание происходит вовсе не обязательно тогда, когда во внешней среде нет потребности в этом гене, в этом признаке. Ведь смотрите сами: огромное количество генетической информации, присутствующей в геноме, не используется.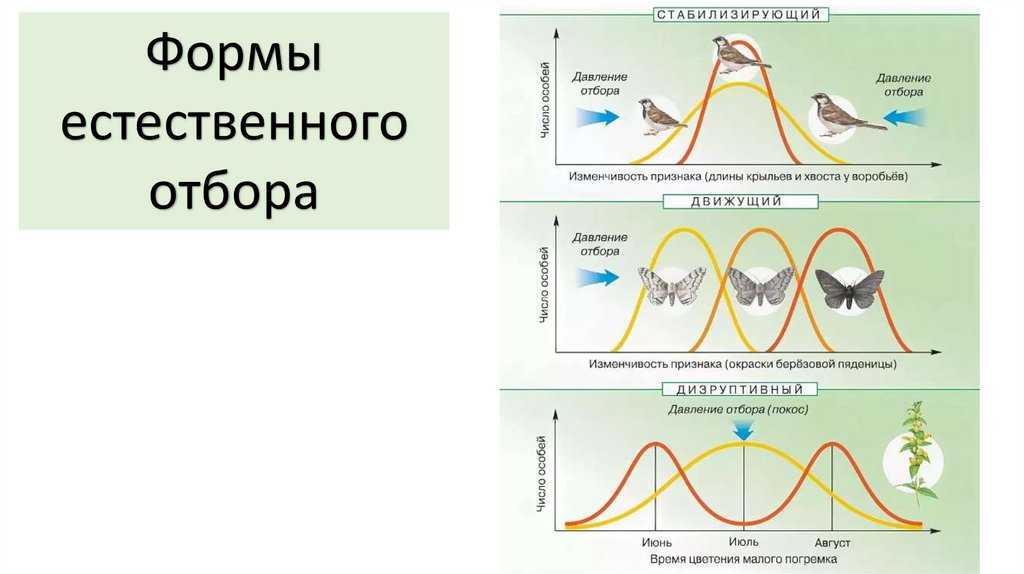 Вы прекрасно знаете, что у высших организмов только небольшая часть ДНК кодирует что-то полезное. У человека это, по-моему, 5%. Вся остальная генетическая информация никак не выражается в признаках и никак не подвержена естественному отбору. У бактерий гораздо компактнее геном, и у них очень мало ненужной информации, но она есть, и она сохраняется миллионы лет. Почему она сохраняется миллионы лет? Потому что с точки зрения полинуклеотидной селекции нет условий для ее отбрасывания. Для того чтобы что-то выбросить, нужно, чтобы для этого были условия в геноме. Вот такими условиями являются, например, прямые повторы, то есть однонаправленные повторы по краям потенциально выбрасываемого фрагмента. Если они есть, такой фрагмент с высокой вероятностью будет выброшен на самом деле. Если внутри этого сегмента содержится нужный, абсолютно необходимый ген для данных условий существования, то выбрасывая его, особь погибает. Но это не значит, что выбрасывание не происходит, оно происходит, просто эта особь гибнет.
Вы прекрасно знаете, что у высших организмов только небольшая часть ДНК кодирует что-то полезное. У человека это, по-моему, 5%. Вся остальная генетическая информация никак не выражается в признаках и никак не подвержена естественному отбору. У бактерий гораздо компактнее геном, и у них очень мало ненужной информации, но она есть, и она сохраняется миллионы лет. Почему она сохраняется миллионы лет? Потому что с точки зрения полинуклеотидной селекции нет условий для ее отбрасывания. Для того чтобы что-то выбросить, нужно, чтобы для этого были условия в геноме. Вот такими условиями являются, например, прямые повторы, то есть однонаправленные повторы по краям потенциально выбрасываемого фрагмента. Если они есть, такой фрагмент с высокой вероятностью будет выброшен на самом деле. Если внутри этого сегмента содержится нужный, абсолютно необходимый ген для данных условий существования, то выбрасывая его, особь погибает. Но это не значит, что выбрасывание не происходит, оно происходит, просто эта особь гибнет.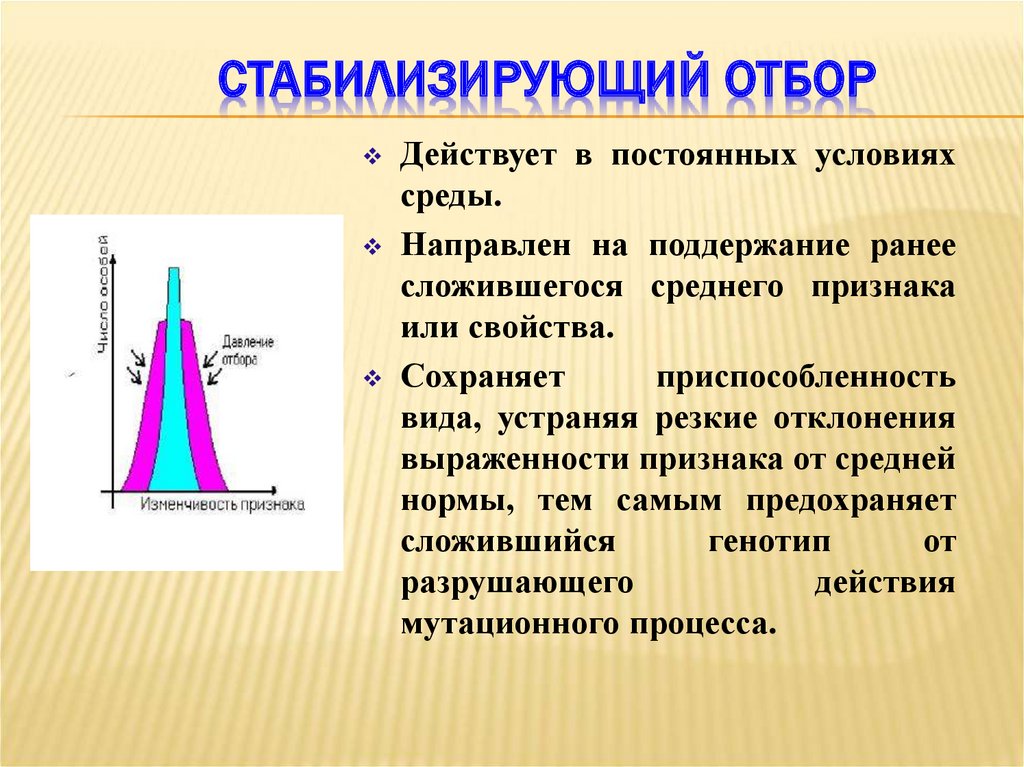 Остается в живых та, которая не выбросит.
Остается в живых та, которая не выбросит.
Случайные мутации – не вполне случайны
Я уверен, что классические эволюционисты до сих пор очень смутно себе представляют мутационный процесс – это очень сложное явление. Более того, я думаю, те мутации, которые считались обычными, о которых всегда говорили, что они возникают случайно в том или ином месте генома, вовсе не так уж случайны. Есть предопределенность и есть некая специфика, о которой мы далеко не все знаем.
Иерархия отбора — Понимание эволюции
Чаще всего мы думаем, что естественный отбор работает на уровне особи, отдавая предпочтение тем, кто лучше оставляет после себя больше индивидуальных потомков. Однако, проявив немного воображения, мы можем увидеть, как естественный отбор может работать и на других уровнях биологической организации. Двигаясь вниз по иерархии, естественный отбор может воздействовать на клетки внутри человека, отдавая предпочтение тем клеточным линиям, которые лучше оставляют после себя клетки-потомки. Двигаясь вверх по иерархии, естественный отбор может воздействовать на виды, благоприятствуя этим видам, которые лучше диверсифицируются в виды-потомки.
Двигаясь вверх по иерархии, естественный отбор может воздействовать на виды, благоприятствуя этим видам, которые лучше диверсифицируются в виды-потомки.
Отбор на индивидуальном уровне
Чтобы понять, как работает отбор на разных уровнях, сначала рассмотрим требования естественного отбора в наиболее знакомой ситуации, на уровне индивидуального организма, на примере популяции жуков:
1. Варьирование признаков. Некоторые жуки зеленые, а некоторые коричневые.
2. Дифференциальное рождение и смерть.
Поскольку окружающая среда не может поддерживать неограниченный рост популяции, не все особи могут размножаться в полной мере. В этом примере зеленые жуки, как правило, поедаются птицами и выживают для размножения реже, чем коричневые жуки.
3. Наследственность.
Поскольку этот признак имеет генетическую основу, у выживших бурых жуков есть коричневые потомки.
В конце концов в популяции становится более распространенным полезный признак – коричневая окраска.
Отбор на клеточном уровне
Для работы этого процесса необходимы всего три основных свойства: изменчивость, дифференцированное рождение и смерть и наследственность. Однако эти черты не уникальны для отдельных популяций. В качестве примера представьте группу клеток в печени одного человека:
1. Различия в признаках.
Большинство клеток в основном одинаковы, но в одной произошла случайная мутация, инактивировавшая ген, контролирующий рост клетки.
2. Дифференциальное рождение и смерть.
Поскольку клетка со случайной мутацией потеряла способность регулировать рост, она делится быстрее остальных.
3. Наследственность.
Нормальные клетки передают нормальную ДНК при делении, а мутантные клетки при делении передают мутантную ДНК.
В конце концов, клетки, несущие мутантную ДНК, становятся все более распространенными в печени и даже могут взять верх, вызывая сбои в работе печени. В этом случае результатом селекции является процесс, более известный как рак. Отбор на клеточном уровне постоянно действует во всех многоклеточных организмах (включая человека), но мы редко замечаем его, за исключением тех случаев, когда он приводит к таким пагубным последствиям. Интересно, что клеточный отбор может работать против отбора на индивидуальном уровне: то, что выгодно для клеточной линии (например, бесконтрольное размножение), может оказаться невыгодным для всего организма (например, вызвать раннюю смерть от рака).
В этом случае результатом селекции является процесс, более известный как рак. Отбор на клеточном уровне постоянно действует во всех многоклеточных организмах (включая человека), но мы редко замечаем его, за исключением тех случаев, когда он приводит к таким пагубным последствиям. Интересно, что клеточный отбор может работать против отбора на индивидуальном уровне: то, что выгодно для клеточной линии (например, бесконтрольное размножение), может оказаться невыгодным для всего организма (например, вызвать раннюю смерть от рака).
Отбор на уровне видов
Двигаясь вверх по иерархии, мы также можем увидеть, как этот же процесс будет работать на целых линиях организмов. В качестве примера представьте себе группу близкородственных видов:
1. Изменчивость признаков.
Некоторые виды имеют очаговое распространение, а другие имеют непрерывное распространение. Неравномерное распространение часто приводит к тому, что изолированные популяции превращаются в новые виды.
2. Дифференциальное рождение и смерть.
Виды с неравномерным распространением чаще других подвергаются видообразованию — аналогу «рождения» вида.
3. Наследственность.
Виды с неоднородным распространением имеют тенденцию видообразования в линии с таким же неоднородным распространением, а виды с непрерывным распространением имеют тенденцию видообразования в линии с непрерывным распространением.
В конце концов, этот процесс благоприятствует видам с неравномерным распространением, и эти виды со временем станут более распространенными, чем виды с непрерывным распространением. Если пятнистая линия окажется пурпурной, а непрерывная линия окажется коричневой, количество пурпурных видов будет увеличиваться быстрее, чем количество коричневых видов.
Отбор на видовом уровне наблюдать гораздо труднее, чем отбор на клеточном или даже индивидуальном уровне, потому что отбор видов происходит на протяжении миллионов лет. Тем не менее, мы знаем, что этот процесс теоретически мог произойти. У нас даже есть некоторые доказательства того, что действительно происходит . Палеонтолог Дэвид Яблонки и его коллеги нашли доказательства того, что географический ареал морских моллюсков может эволюционировать таким образом. Они собрали данные о нахождении более 1000 различных видов моллюсков, окаменевших в позднемеловых породах юго-востока Северной Америки, и обнаружили несколько ключевых закономерностей:
Тем не менее, мы знаем, что этот процесс теоретически мог произойти. У нас даже есть некоторые доказательства того, что действительно происходит . Палеонтолог Дэвид Яблонки и его коллеги нашли доказательства того, что географический ареал морских моллюсков может эволюционировать таким образом. Они собрали данные о нахождении более 1000 различных видов моллюсков, окаменевших в позднемеловых породах юго-востока Северной Америки, и обнаружили несколько ключевых закономерностей:
1. Различия в признаках.
Географический ареал зависит от вида моллюсков. Некоторые имеют широкий диапазон, а некоторые имеют небольшие диапазоны.
2. Дифференциальное рождение и смерть.
Те виды моллюсков с широким ареалом устойчивы к вымиранию (т. е. имеют низкий уровень «смерти»).
3. Наследственность.
Когда моллюски с широким географическим ареалом образуют виды, они имеют тенденцию давать начало другим видам с широким географическим ареалом — другими словами, географический ареал является наследуемым признаком для этих видов.
Морские моллюски, кажется, обладают всеми основными характеристиками, которые привели бы к отбору видов, но насколько важен этот процесс в их эволюционной истории и истории жизни в целом? Ученые до сих пор ищут ответ на этот вопрос. Хотя мы знаем, что отбор может действовать на уровнях выше индивидуального, все еще неясно, насколько важен этот процесс в формировании истории жизни.
Эволюция и естественный отбор
— Ральф Уолдо Эмерсон Я назвал этот принцип, согласно которому каждый
небольшие вариации, если они полезны, сохраняются под термином «естественный отбор».
В этом уроке мы хотим спросить:
(R)Эволюция теории Теория эволюции является одной из
великие интеллектуальные революции в истории человечества, коренным образом изменившие нашу
восприятие мира и своего места в нем. Чарльз Дарвин выдвинул
последовательной теории эволюции и собрал большое количество доказательств в поддержку
этой теории. Во времена Дарвина большинство ученых полностью верили, что каждый
организма и каждое приспособление было делом творца. В прочем слова, все сходства и различия между группами организмов которые являются результатом процесса ветвления, создающего великое древо жизни ( см. рис. 1 ), рассматривались философами начала 19 века и ученых как следствие всемогущего замысла.
Однако, к 19 веку ряд естествоиспытателей начали думать эволюционных изменений как объяснение закономерностей, наблюдаемых в природе. следующие идеи были частью интеллектуального климата времени Дарвина.
Теория Дарвина Теория эволюции Дарвина влечет за собой следующие основные идеи. Первые три идеи уже обсуждались среди естествоиспытателей прошлого и современников, работавших над «проблемой видов», когда Дарвин начал свои исследования. Первоначальным вкладом Дарвина был механизм естественного отбора и огромное количество свидетельств эволюционных изменений из многих источников. Он также дал вдумчивые объяснения последствий эволюции для нашего понимания истории жизни и современного биологического разнообразия.
Основным механизмом изменений с течением времени является естественный отбор, подробно описанный ниже. Этот механизм вызывает изменение свойств (признаков) организмов внутри родословных от поколения к поколению. Процесс естественного отбора Дарвиновский процесс естественного отбора состоит из четырех компонентов.
От поколения к поколению борьба за ресурсы (то, что Дарвин называл «борьбой за существование») будет отдавать предпочтение особям с некоторыми вариациями по сравнению с другими и, таким образом, изменять частоту признаков в популяции. Этот процесс является естественным отбором. Признаки, дающие преимущество тем особям, которые оставляют больше потомства, называются адаптациями. Чтобы естественный отбор воздействовал на признак, он должен обладать наследуемой изменчивостью и давать преимущество в борьбе за ресурсы. Если одно из этих требований не выполняется, то признак не подвергается естественному отбору. Естественный отбор действует на основе сравнительного преимущества, а не абсолютного стандарта дизайна. «… как естественный отбор действует путем конкуренции за ресурсы, он приспосабливает жителей каждой страны только по отношению к степени совершенства их -соратников» (Чарльз Дарвин, О происхождении видов , 1859). В ХХ веке генетика была интегрирована с дарвиновским механизмом, что позволило нам оценить естественный отбор как дифференцированное выживание и воспроизводство генотипов, соответствующих определенным фенотипам. Естественный отбор может воздействовать только на существующую изменчивость внутри популяции. Такие вариации возникают в результате мутации, изменения какой-либо части генетического кода признака. Мутации возникают случайно и без предвидения потенциального преимущества или недостатка мутации. Другими словами, вариации возникают не потому, что они необходимы. Доказательства естественного отбораДавайте посмотрите на пример, чтобы помочь понять естественный отбор. Промышленный меланизм — это явление, поразившее более 70 видов мотыльков в Англии. Лучше всего он изучен у перечной моли Biston betularia . Прежний до 1800 г. типичная бабочка этого вида имела светлый рисунок ( см. рис. 2 ). Мотыльки темного цвета или меланические бабочки были редкостью и поэтому были предметами коллекционирования.
Во время
промышленная революция, сажа и другие промышленные отходы потемнели дерево
стволы и отмершие лишайники. Светлая морфа мотылька стала
редки, а темная морфа стала обильной. В 1819 году была получена первая меланическая морфа.
видимый; к 1886 году она стала гораздо более распространенной, что свидетельствует о быстрой эволюции
изменять. В конце концов легкие морфы были распространены лишь в нескольких местах, вдали от промышленных районов. Считалось, что причиной этого изменения является избирательное хищничество птиц. что способствовало маскировочной окраске бабочки. В В 1950-х годах биолог Кеттлвелл провел эксперименты по освобождению и повторной поимке, используя обе морфы. Краткое изложение его результатов показано ниже. Наблюдая хищничество птиц из жалюзи, он мог подтвердить, что заметность мотылька сильно повлияло на вероятность того, что его съедят. Повторить успех
Локальная адаптация — дополнительные примеры Так
далеко в сегодняшней лекции мы подчеркивали, что естественный отбор является
Краеугольный камень эволюционной теории. Ниже приведены примеры, иллюстрирующие адаптацию популяций к местным условиям. условия.
Рисунок 3: Подвиды крысиной змеи Elaphe obsoleta , которые скрещиваются там, где встречаются их ареалы.
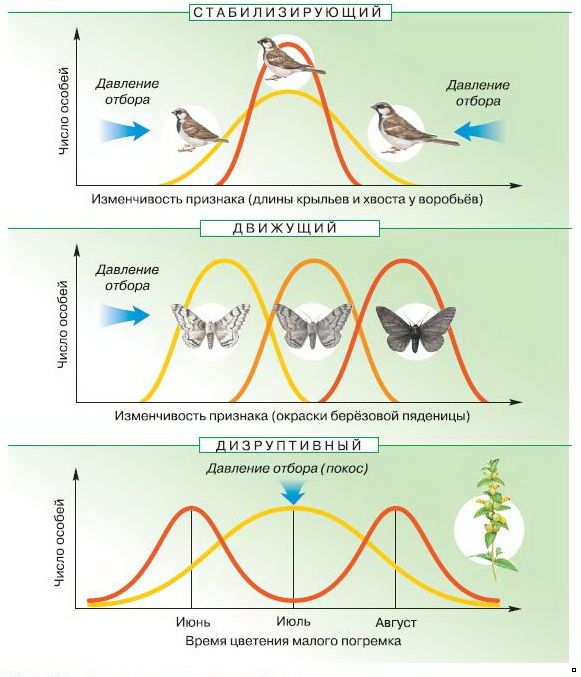
Стабилизирующие, направленные и Разнообразный выборНаконец, мы рассмотрим статистический способ мышления об отборе. Предположим, что каждую популяцию можно изобразить как частотное распределение некоторого признака — размер клюва, например. Еще раз отметим, что вариация признака – это важнейшее сырье для эволюции. Что будет ли частотное распределение выглядеть в следующем поколении?
Первый,
доля особей с каждым значением признака (размер клюва или
масса тела) может быть точно такой же.
Под стабилизирующая селекция , крайние сорта с обоих концов частотное распределение исключено. Распределение частот выглядит точно так же, как это было в предыдущем поколении ( см. рис. 5a ). Вероятно это самая распространенная форма естественного отбора, и мы часто ошибаемся за отсутствие выбора. Примером из реальной жизни является вес при рождении человеческих младенцев. ( см. рис. 6 ). Под направленный отбор , особи на одном конце распределения
размеры клюва особенно хороши, поэтому частотное распределение признака
в последующем поколении смещается с того места, где оно было в родительском
поколения ( см.
ископаемое происхождение лошади представляет собой замечательную демонстрацию направленная преемственность. Полная родословная довольно сложна и не просто простая линия от крошечной рассветной лошади Hyracotherium раннего Эоцен, к сегодняшнему знакомому Equus . Но в целом лошадь произошел от предка с небольшим телом, созданного для передвижения по лесным массивам и зарослей до своего длинноногого потомка, построенного для скорости на открытых пастбищах. Эта эволюция включала хорошо задокументированные изменения в зубах, длине ног, и структура пальцев ног ( см. рис. 7 ). Под диверсифицирующий (подрывной) отбор , обе крайности предпочтительны на
за счет промежуточных сортов ( см. РезюмеДарвина теория эволюции коренным образом изменила направление будущих научных мысль, хотя она и была построена на растущем массиве мыслей, который начал подвергнуть сомнению прежние представления о мире природы. Ядром теории Дарвина является естественный отбор, процесс, который происходит в течение последовательных поколений и определяется как дифференциальное воспроизводство генотипы. Натуральный отбор требует наследственной изменчивости данного признака, а дифференциальная выживание и размножение, связанные с обладанием этим признаком. Примеры естественного отбора хорошо задокументированы как наблюдениями, так и Окаменелости. Выбор
действует на частоту признаков и может принимать форму стабилизирующих,
направленный или диверсифицирующий отбор. About the AuthorRelated Posts |
 Линней
создал систему биологической классификации, которую мы используем сегодня, и
сделал это в духе каталогизации Божьих творений.
Линней
создал систему биологической классификации, которую мы используем сегодня, и
сделал это в духе каталогизации Божьих творений.
 существующие виды.
существующие виды.
 мышление. Изображение в верхней части этой страницы принадлежит Дарвину.
зарисовки вьюрков в его «Журнале исследований».
мышление. Изображение в верхней части этой страницы принадлежит Дарвину.
зарисовки вьюрков в его «Журнале исследований».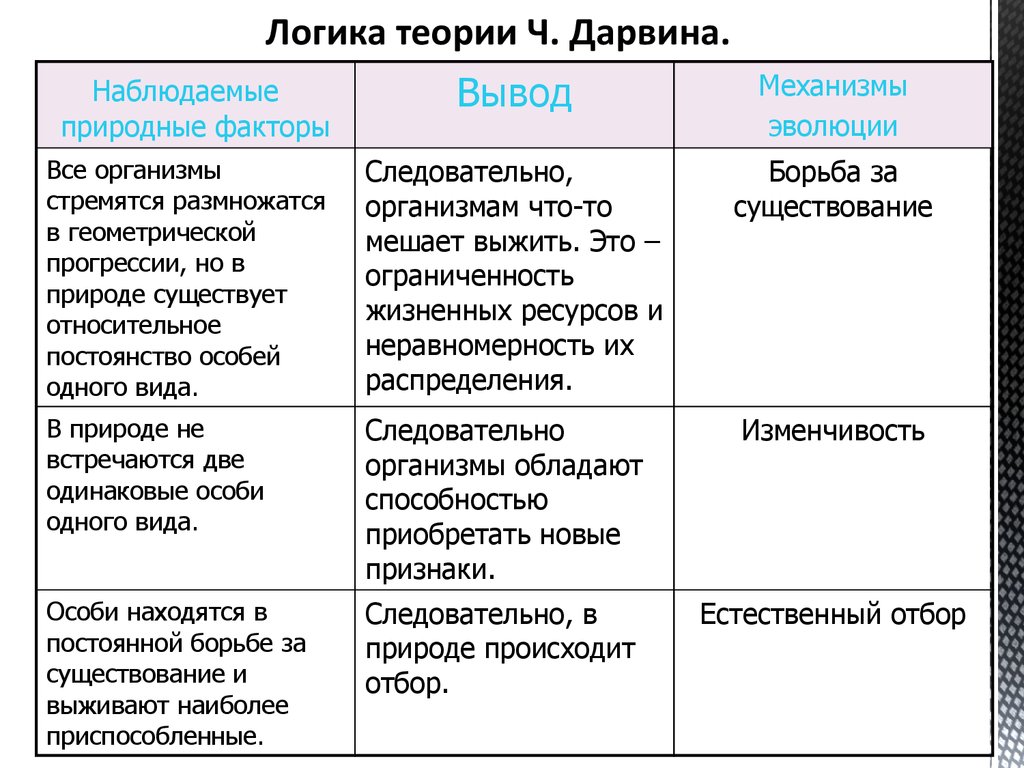 Что означает эта теория
эволюции говорите?
Что означает эта теория
эволюции говорите? 

 У большинства популяций каждый год рождается больше потомства, чем могут обеспечить местные ресурсы, что приводит к борьбе за ресурсы. Каждое поколение переживает значительную смертность.
У большинства популяций каждый год рождается больше потомства, чем могут обеспечить местные ресурсы, что приводит к борьбе за ресурсы. Каждое поколение переживает значительную смертность. (Теперь мы знаем, что такие черты могут изменяться в результате других эволюционных механизмов, открытых со времен Дарвина.)
(Теперь мы знаем, что такие черты могут изменяться в результате других эволюционных механизмов, открытых со времен Дарвина.)

 Он обеспечивает механизм адаптивного
изменять. Любое изменение в окружающей среде (например, изменение фона)
цвет ствола дерева, на котором вы сидите) может привести к
приспособление. Любая широко распространенная популяция, вероятно, будет испытывать различные
экологические условия в разных частях его ареала. Как следствие это
скоро будет состоять из ряда субпопуляций, которые незначительно отличаются друг от друга, или
даже значительно.
Он обеспечивает механизм адаптивного
изменять. Любое изменение в окружающей среде (например, изменение фона)
цвет ствола дерева, на котором вы сидите) может привести к
приспособление. Любая широко распространенная популяция, вероятно, будет испытывать различные
экологические условия в разных частях его ареала. Как следствие это
скоро будет состоять из ряда субпопуляций, которые незначительно отличаются друг от друга, или
даже значительно. лекцию о видообразовании).
лекцию о видообразовании). ) Дарвину казалось, что
что каждый из них был слегка изменен по сравнению с первоначальным колонистом, вероятно,
зяблик на материковой части Южной Америки, примерно в 600 милях к востоку. Это
Вероятно, адаптивная радиация привела к образованию столь многих
видов, потому что других птиц было мало или они отсутствовали, оставляя пустые ниши для
наполнять; и поскольку многочисленные Галапагосские острова давали достаточно
возможность географической изоляции.
) Дарвину казалось, что
что каждый из них был слегка изменен по сравнению с первоначальным колонистом, вероятно,
зяблик на материковой части Южной Америки, примерно в 600 милях к востоку. Это
Вероятно, адаптивная радиация привела к образованию столь многих
видов, потому что других птиц было мало или они отсутствовали, оставляя пустые ниши для
наполнять; и поскольку многочисленные Галапагосские острова давали достаточно
возможность географической изоляции. Во-вторых, могут быть направленные
измениться только в одном направлении. Третий (и с такой редкостью, что его существование
является спорным), могут быть одновременные изменения в обоих направлениях (например,
предпочтительны как большие, так и меньшие клювы за счет клювов
средний размер). На рисунках 5a-c показаны эти три основные категории
естественный отбор.
Во-вторых, могут быть направленные
измениться только в одном направлении. Третий (и с такой редкостью, что его существование
является спорным), могут быть одновременные изменения в обоих направлениях (например,
предпочтительны как большие, так и меньшие клювы за счет клювов
средний размер). На рисунках 5a-c показаны эти три основные категории
естественный отбор. рис. 5b ). Это то, что мы обычно думаем как
естественный отбор. Промышленный меланизм был таким примером.
рис. 5b ). Это то, что мы обычно думаем как
естественный отбор. Промышленный меланизм был таким примером. рис. 5с ). Это редкость,
но представляет теоретический интерес, поскольку предлагает механизм для видов
образование без географической изоляции (см. лекцию о видообразовании).
рис. 5с ). Это редкость,
но представляет теоретический интерес, поскольку предлагает механизм для видов
образование без географической изоляции (см. лекцию о видообразовании).