Естественный отбор — главный фактор эволюции
Главной заслугой Чарлза Дарвина было верное описание механизма эволюции, ее причин, или факторов; то есть того, как на самом деле она протекает. Сама же мысль, что виды меняются, постепенно приспосабливаясь к окружающей среде, высказывалась еще до Дарвина. Так Жан Батист Ламарк внес большой вклад в развитие эволюционного учения, хотя неправильно описал то, как протекает эволюция.
Естественный отбор, которому большое внимание уделил Дарвин, и есть та самая, главная движущая сила эволюции. В отличие от других факторов эволюции (дрейфа генов, мутаций, комбинативной изменчивости) только естественный отбор обладает направленной силой. Его действие не приводит к случайным изменениям, а всегда идет в сторону все большего приспособления организмов к окружающей среде.
Естественный отбор – это выживание наиболее приспособленных к данной среде особей в результате их борьбы за существование в условиях ограниченных ресурсов и на основе мутаций и комбинативной изменчивости, обеспечивающих отличие особей между собой.
Причины естественного отбора
Естественный отбор есть следствие действия совокупности других факторов эволюции. В основном борьбы за существование и наследственной изменчивости.
Организмы способны дать намного больше потомства, чем вместимость среды обитания. В результате между особями возникает борьба за существование – борьба за жизнь, пищу, территорию, право на размножение. Также индивиды и популяции борются не только между собой, но и с неблагоприятными условиями среды. В общей массе выживают наиболее успешные в этой борьбе. Они производят следующее поколение, обладающее их признаками.
Организмы в пределах одной популяции не одинаковы. Они так или иначе отличаются между собой. Другими словами, организмы обладают изменчивостью – способностью приобретать отличительные или новые признаки. Часть из таких признаков передается по наследству, формируя так называемую
наследственную изменчивость.Особенности конкретной среды обитания, ее малая вместимость способствуют выживанию организмов с такой наследственной изменчивостью, которая позволяет им выигрывать борьбу за существование.
Следствие естественного отбора – адаптация, или приспособленность
В процессе естественного отбора происходит приспособление особей и популяций к их среде обитания, как абиогенной – к неживой природе, так и биогенной – к живой природе – биоценозу. На конкретном отрезке времени популяция обладает определенной степенью приспособленности к среде, то есть различными адаптациями к ней.
Адаптацию можно рассматривать и как процесс, увеличивающий приспособленность, и как результат, выражающийся в наличии определенных приспособлений.
Приспособления могут быть различными. Они зависят от среды обитания и исходных характеристик вида. Разные виды могут адаптироваться к одной и той же среде обитания по-разному.
Адаптация, отталкиваясь от имеющейся морфологии и физиологии вида, меняет их так, чтобы популяция как можно быстрее и надежнее заняла определенную экологическую нишу в биоценозе.
Можно сказать, любой признак организма есть следствие адаптации к тем или иным условиям, возникшим в процессе эволюции данного вида.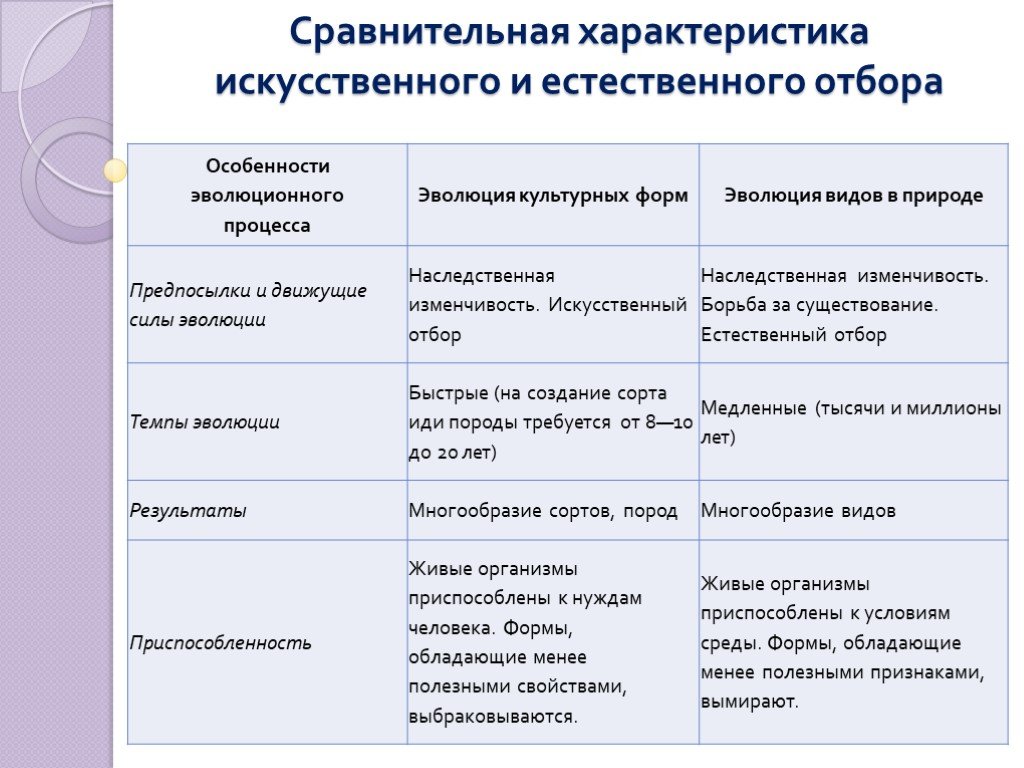 Другими словами, есть признаки, которые были адаптивными к условиям среды предков. У потомков они могут не исчезать, а претерпевать модификацию.
Другими словами, есть признаки, которые были адаптивными к условиям среды предков. У потомков они могут не исчезать, а претерпевать модификацию.
Среди ярких и простых примеров адаптаций часто выделяют мимикрию, покровительственную окраску. Большинство же адаптаций имеют сложный комплексный характер, затрагивающий несколько систем органов.
Адаптации имеют относительный характер. При смене условий обитания они могут утратить свое полезное значение. И требуется время, смена поколений для выработки новых.
Формы естественного отбора
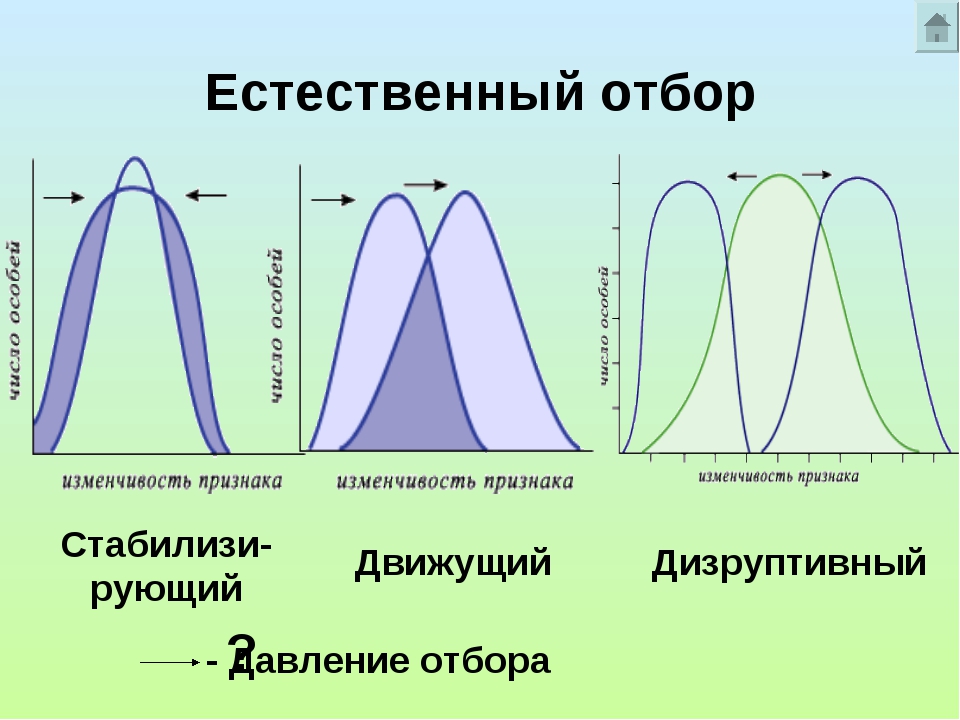
В зависимости от сложившихся условий среды естественный отбор может отбирать особей по-разному. То есть в одних условиях лучше будут выживать одни, в других – вполне может быть, что другие. На этом основании выделяют три основные формы естественного отбора – стабилизирующий, движущий и дизруптивный.
Желтые стрелки – направление действия естественного отбора. Красный график – прежнее состояние популяции.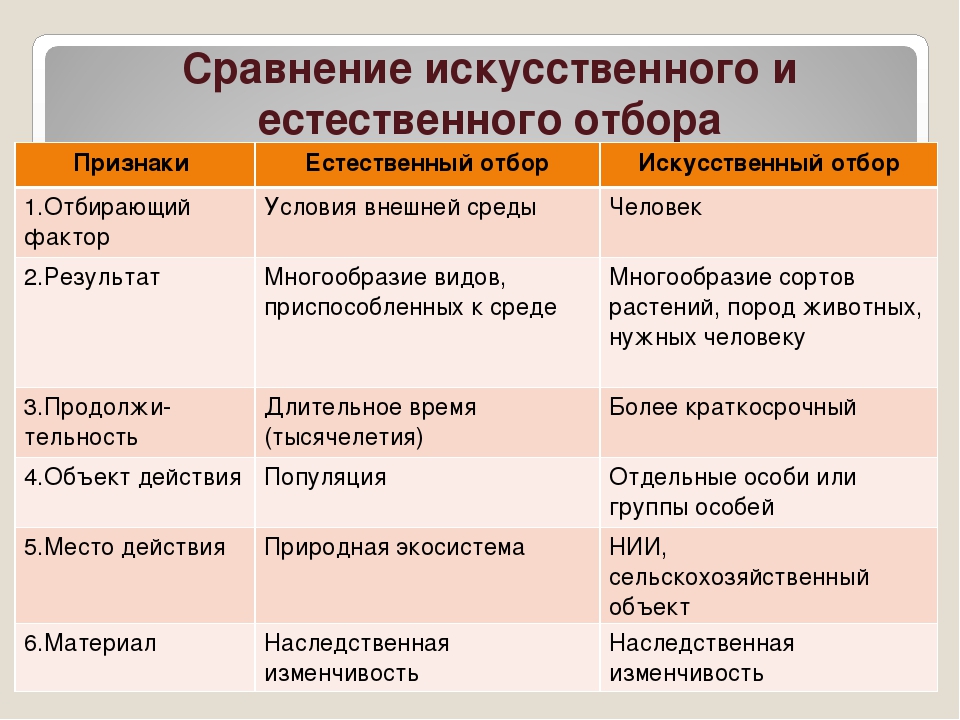 Синий график – состояние популяции после действия естественного отбора. Ось X – значение признака. Ось Y – количество особей.
Синий график – состояние популяции после действия естественного отбора. Ось X – значение признака. Ось Y – количество особей.В случае стабилизирующей формы отбор благоприятствует выживанию особей со средним значением признака и устраняет особей с любыми крайними значениями. Стабилизирующий отбор действует в неизменных условиях среды на популяции, которые давно здесь обитают. Он поддерживает неизменность, то есть стабильность, популяций.
Если условия среды меняются, то на смену стабилизирующему приходит движущий естественный отбор. Обычно при смене условий в популяции могут найтись особи с каким-либо более подходящим значением признака. Естественный отбор будет давить на популяцию так, чтобы количество таких особей увеличивалось.
Движущий естественный отбор всегда действует в одном направлении, сдвигая среднее значение признака в какую-либо одну сторону. Модель ниже можно описать так. Условия среды изменились и погибли все особи, кроме тех, которые обладали определенным значением признака.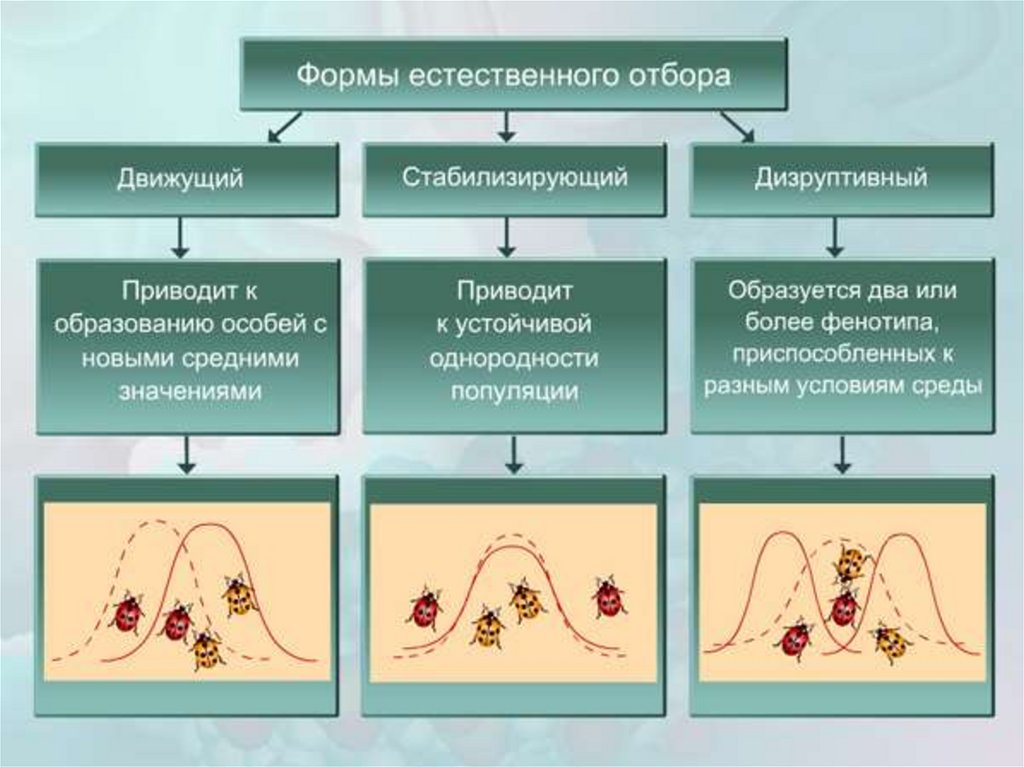 После они размножились, и признак из редкого превратился в распространенный.
После они размножились, и признак из редкого превратился в распространенный.
Дизруптивный, или разрывающий, отбор можно представить как частный случай движущего, так как он тоже приводит к исчезновению старой нормы. Однако, в отличие от движущего, при дизруптивном отборе формируется не одна, а две новые нормы из ранее крайних значений признака. Другими словами, при изменении условий часть популяции начинает приспосабливаться не так, как другая ее часть.
естественный отбор | это… Что такое естественный отбор?
есте́ственный отбо́рпроцесс выживания и воспроизведения организмов, наиболее приспособленных к условиям среды, и гибели в ходе эволюции неприспособленных. Естественный отбор — следствие борьбы за существование; обусловливает относительную целесообразность строения и функций организмов; творческая роль естественного отбора выражается в преобразовании популяций, приводящем к появлению новых видов. Понятие о естественном отборе как основном движущем факторе исторического развития живой природы введено Ч. Дарвином.
Дарвином.
* * *
ЕСТЕСТВЕННЫЙ ОТБОРЕСТЕ́СТВЕННЫЙ ОТБО́Р, основной фактор эволюции организмов, значение и механизм действия которого были вскрыты Ч. Дарвином (
Логика рассуждений Дарвина была безупречна: если среди организмов данного вида или разновидности каждая особь чем-то отличается от всех других особей, то есть существует индивидуальная изменчивость, то среди этих организмов всегда существуют и более приспособленные, и менее приспособленные к условиям окружающей среды.
 В борьбе за существование (см. БОРЬБА ЗА СУЩЕСТВОВАНИЕ) более приспособленные чаще выживают — подвергаются естественному отбору, а менее приспособленные чаще гибнут. Это происходит в каждом поколении, а в чреде поколений полезные изменения накапливаются, организмы постепенно становятся непохожими на своих предков. В конечном итоге, благодаря естественному отбору возникают новые виды.
В борьбе за существование (см. БОРЬБА ЗА СУЩЕСТВОВАНИЕ) более приспособленные чаще выживают — подвергаются естественному отбору, а менее приспособленные чаще гибнут. Это происходит в каждом поколении, а в чреде поколений полезные изменения накапливаются, организмы постепенно становятся непохожими на своих предков. В конечном итоге, благодаря естественному отбору возникают новые виды. Сформулированная Дарвином теория, объяснившая действием естественного отбора и приспособленность организмов к условиям их жизни, и многообразие видов, составляющих биосферу, полностью сохранила свое значение. Все попытки опровергнуть ее, а они были многочисленными и с позиций отрицания самого явления эволюции, и с позиций отрицания значения естественного отбора в этом процессе, до сих пор не увенчались успехом.
 Теперь известно, как возникает наследственная индивидуальная изменчивость, как происходит борьба за существование, в каких условиях и в каком направлении действует естественный отбор. Правда, эволюция — процесс медленный. Формирование нового вида занимает десятки — сотни тысяч лет. Поэтому наблюдать естественный отбор почти невозможно. В биологической литературе мало исследований, в которых непосредственно прослежен этот процесс. Тем не менее результаты экспериментов, анализ причин дифференциальных смертности и выживания организмов в природе, реконструкции, в которых об отборе судят по его результатам, постепенно сформировали современную теорию естественного отбора.
Теперь известно, как возникает наследственная индивидуальная изменчивость, как происходит борьба за существование, в каких условиях и в каком направлении действует естественный отбор. Правда, эволюция — процесс медленный. Формирование нового вида занимает десятки — сотни тысяч лет. Поэтому наблюдать естественный отбор почти невозможно. В биологической литературе мало исследований, в которых непосредственно прослежен этот процесс. Тем не менее результаты экспериментов, анализ причин дифференциальных смертности и выживания организмов в природе, реконструкции, в которых об отборе судят по его результатам, постепенно сформировали современную теорию естественного отбора. Наследственная изменчивость — материал для отбора
Материалом для естественного отбора служит наследственная изменчивость, а источником наследственной изменчивости являются мутации — наследуемые изменения генетического материала. Для эволюции имеют значение только те мутации, которые возникают в половых клетках, так как только они передаются следующему поколению.
 Большинство вновь возникающих мутаций репарируются — исправляются ферментными системами клетки. Однако часть из них остается — ферменты не «узнают» их. Такие мутации относительно редки. В среднем они возникают с частотой одна мутация на 1000000 гомологичных генов (1,10-6). Однако геном, например, человека содержит не менее 30000 структурных генов, то есть генов, кодирующих синтез белков. Количество регуляторных генов, по-видимому, не меньше. Поэтому в пересчете на геном мутации не так уж редки. Главное же в том, что мутации накапливаются. Большинство вновь возникающих мутаций рецессивны, т. е. их действие подавляют доминантные, «нормальные» гены. Поэтому рецессивные мутации сохраняются в генофонде популяции, не проявляясь в фенотипе и не влияя на приспособленность организмов. Поскольку мутационный процесс идет непрерывно, число рецессивных мутаций в генофонде увеличивается от поколения к поколению.
Большинство вновь возникающих мутаций репарируются — исправляются ферментными системами клетки. Однако часть из них остается — ферменты не «узнают» их. Такие мутации относительно редки. В среднем они возникают с частотой одна мутация на 1000000 гомологичных генов (1,10-6). Однако геном, например, человека содержит не менее 30000 структурных генов, то есть генов, кодирующих синтез белков. Количество регуляторных генов, по-видимому, не меньше. Поэтому в пересчете на геном мутации не так уж редки. Главное же в том, что мутации накапливаются. Большинство вновь возникающих мутаций рецессивны, т. е. их действие подавляют доминантные, «нормальные» гены. Поэтому рецессивные мутации сохраняются в генофонде популяции, не проявляясь в фенотипе и не влияя на приспособленность организмов. Поскольку мутационный процесс идет непрерывно, число рецессивных мутаций в генофонде увеличивается от поколения к поколению. 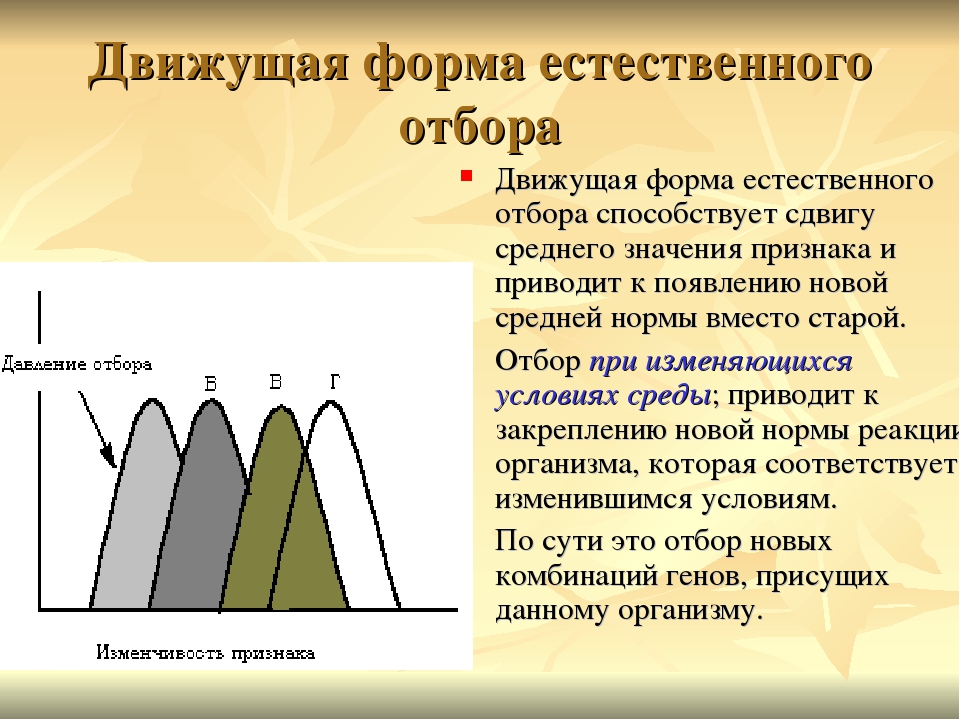 Они вызывают различные нарушения обмена веществ или морфологические изменения — уродства. Однако часть фенотипических проявлений мутаций может оказываться полезной при изменении условий существования популяции. Например, у комнатной мухи известна мутация, снижающая скорость проведения нервных импульсов. В гомозиготном (см. ГОМОЗИГОТА)состоянии такая мутация летальна. Гетерозиготные мутантные мухи проигрывают по приспособленности нормальным, но жизнеспособны, так как нормальный аллель частично компенсирует вредный эффект мутации. Однако, если на популяцию мух, в генофонде которой есть эта мутация, воздействуют инсектицидом нервно-паралитического действия, мухи, гетерозиготные по этой мутации, оказываются более жизнеспособными, чем нормальные, — они выживают. Медленное прохождение нервных импульсов у таких мух ослабляет действие яда. Поэтому естественный отбор поддерживает высокую частоту этой мутации во многих популяциях мух. Аналогично, во многих популяциях людей, живущих в районах, где свирепствует малярия, распространена мутация серповидноклеточной анемии (см.
Они вызывают различные нарушения обмена веществ или морфологические изменения — уродства. Однако часть фенотипических проявлений мутаций может оказываться полезной при изменении условий существования популяции. Например, у комнатной мухи известна мутация, снижающая скорость проведения нервных импульсов. В гомозиготном (см. ГОМОЗИГОТА)состоянии такая мутация летальна. Гетерозиготные мутантные мухи проигрывают по приспособленности нормальным, но жизнеспособны, так как нормальный аллель частично компенсирует вредный эффект мутации. Однако, если на популяцию мух, в генофонде которой есть эта мутация, воздействуют инсектицидом нервно-паралитического действия, мухи, гетерозиготные по этой мутации, оказываются более жизнеспособными, чем нормальные, — они выживают. Медленное прохождение нервных импульсов у таких мух ослабляет действие яда. Поэтому естественный отбор поддерживает высокую частоту этой мутации во многих популяциях мух. Аналогично, во многих популяциях людей, живущих в районах, где свирепствует малярия, распространена мутация серповидноклеточной анемии (см. СЕРПОВИДНОКЛЕТОЧНАЯ АНЕМИЯ). Эта мутация снижает сродство гемоглобина к кислороду и в гомозиготном состоянии тоже летальна, причем дети погибают в раннем возрасте. Однако, в гетерозиготном состоянии эта мутация, хотя и вызывает анемию, поддерживается естественным отбором. Одним из многих ее проявлений является изменение формы эритроцитов. В норме они округлые, а при серповидноклеточной анемии похожи на молодой месяц. Плазмодии — возбудители малярии — не могут проникнуть в такие эритроциты, и люди не болеют малярией. За приспособленность гетерозигот популяция «платит» детской смертностью гомозигот по этой мутации.
СЕРПОВИДНОКЛЕТОЧНАЯ АНЕМИЯ). Эта мутация снижает сродство гемоглобина к кислороду и в гомозиготном состоянии тоже летальна, причем дети погибают в раннем возрасте. Однако, в гетерозиготном состоянии эта мутация, хотя и вызывает анемию, поддерживается естественным отбором. Одним из многих ее проявлений является изменение формы эритроцитов. В норме они округлые, а при серповидноклеточной анемии похожи на молодой месяц. Плазмодии — возбудители малярии — не могут проникнуть в такие эритроциты, и люди не болеют малярией. За приспособленность гетерозигот популяция «платит» детской смертностью гомозигот по этой мутации. Отбор по генетически детерминированным признакам
Случаи, когда материалом естественного отбора служат непосредственно мутации, относительно редки. Большинство признаков многоклеточных организмов зависит от действия многих генов — они полигенны. Грубо говоря, нет гена формы носа, но есть много генов, влияющих на его форму. Большинство мутаций проявляется в фенотипе в сочетании с действием многих других генов, причем генов, уже прошедших естественный отбор, то есть полезных и нормальных.
 Это ослабляет вредное действие мутаций и предотвращает их элиминацию (выбраковку) из генофонда вместе с гибелью менее приспособленных организмов. Такие мутантные гены включаются в процесс естественного отбора и в череде поколений продолжают комбинироваться с другими генами таким образом, что возникающие комбинации повышают приспособительное значение признаков фенотипа. Параллельно, благодаря влиянию других генов, первично рецессивный мутантный ген постепенно становится доминантным и распространяется в генофонде как нормальный наследственный элемент особей данной популяции.
Это ослабляет вредное действие мутаций и предотвращает их элиминацию (выбраковку) из генофонда вместе с гибелью менее приспособленных организмов. Такие мутантные гены включаются в процесс естественного отбора и в череде поколений продолжают комбинироваться с другими генами таким образом, что возникающие комбинации повышают приспособительное значение признаков фенотипа. Параллельно, благодаря влиянию других генов, первично рецессивный мутантный ген постепенно становится доминантным и распространяется в генофонде как нормальный наследственный элемент особей данной популяции. Известным примером подобного действия естественного отбора является так называемый индустриальный меланизм у бабочки березовой пяденицы. Эта бабочка широко распространена в Европе и Северной Америке. Бабочки активны в сумерках, а день проводят, сидя на коре берез. Крылья их имеют покровительственную окраску: они бледно-серые с разводами, что имитирует цвет бересты, заросшей лишайниками. Такая окраска защищает бабочек от насекомоядных птиц — синиц, поползней и др.
 История эволюции окраски английских популяций березовой пяденицы началась в 1735 году, когда англичане впервые в мире применили каменный уголь для выплавки чугуна. Промышленная копоть во влажном климате Англии оседала на коре берез. Лишайники постепенно погибали, а береста становилась сначала серой, а затем черной. Пяденицы на темном фоне стали легкой добычей птиц. В 1848 году английские энтомологи-любители обнаружили и поместили в свои коллекции первых березовых пядениц с темноокрашенными крыльями. Такие бабочки в окрестностях Манчестера составляли не более 1% от численности популяции. К 1960-м годам промышленная копоть покрыла почти все леса Великобритании, и темные пяденицы постепенно вытеснили светлых. Светлая форма преобладает только там, где атлантические циклоны уносят копоть на восток. В результате серии экологических экспериментов и наблюдений было доказано, что птицы действительно чаще съедают тех бабочек, окраска которых контрастирует с фоном — корой дерева. В задымленном лесу близ Бирмингема выпустили одновременно 50 светлых и 50 темных бабочек и затем посчитали, сколько бабочек съели птицы.
История эволюции окраски английских популяций березовой пяденицы началась в 1735 году, когда англичане впервые в мире применили каменный уголь для выплавки чугуна. Промышленная копоть во влажном климате Англии оседала на коре берез. Лишайники постепенно погибали, а береста становилась сначала серой, а затем черной. Пяденицы на темном фоне стали легкой добычей птиц. В 1848 году английские энтомологи-любители обнаружили и поместили в свои коллекции первых березовых пядениц с темноокрашенными крыльями. Такие бабочки в окрестностях Манчестера составляли не более 1% от численности популяции. К 1960-м годам промышленная копоть покрыла почти все леса Великобритании, и темные пяденицы постепенно вытеснили светлых. Светлая форма преобладает только там, где атлантические циклоны уносят копоть на восток. В результате серии экологических экспериментов и наблюдений было доказано, что птицы действительно чаще съедают тех бабочек, окраска которых контрастирует с фоном — корой дерева. В задымленном лесу близ Бирмингема выпустили одновременно 50 светлых и 50 темных бабочек и затем посчитали, сколько бабочек съели птицы. Оказалось, что было съедено 43 светлых и 15 темных. Показательно, что, хотя темная окраска защищает бабочек, эта защита не абсолютна. Более высокая приспособленность не гарантирует выживания, а только повышает его вероятность. Естественный отбор — вероятностный, а не детерминистический процесс.
Оказалось, что было съедено 43 светлых и 15 темных. Показательно, что, хотя темная окраска защищает бабочек, эта защита не абсолютна. Более высокая приспособленность не гарантирует выживания, а только повышает его вероятность. Естественный отбор — вероятностный, а не детерминистический процесс. Были проведены и генетические опыты, показавшие, что темная окраска крыльев доминантна по отношению к светлой и определяется, на первый взгляд, одним геном. Во втором поколении, когда происходит расщепление генов, строго выдерживается соотношение: 3 черных к 1 светлой, как в знаменитых опытах Г. Менделя (см. МЕНДЕЛЬ Грегор Иоганн) по наследованию признаков у гороха. Казалось бы, что у пядениц возникла полезная доминантная мутация, которая затем распространилась во многих популяциях этой бабочки. Однако, когда для проверки этого предположения ученые скрестили английских черных бабочек со светлыми бабочками из Канады, результат оказался неожиданным. В первом поколении окраска бабочек была промежуточной — серой, а расщепление во втором поколении дало соотношение, характерное для неполного доминирования: 1 черная, 2 серые, 1 светлая.
 Это означает, что черная окраска — результат не единичной доминантной мутации, а следствие комбинирования генов, в ходе которого черная окраска стала доминантной, а исходная светлая — рецессивной. Бабочки, сохранившиеся в коллекциях энтомологов с середины прошлого века, имеют не столь темную окраску, как современные. Это указывает на то, что потемнение бабочек происходило постепенно, в череде поколений. Таким образом, преобразование генетического определения признаков фенотипа — это сложный и постепенный процесс, основанный на комбинировании генов под действием естественного отбора.
Это означает, что черная окраска — результат не единичной доминантной мутации, а следствие комбинирования генов, в ходе которого черная окраска стала доминантной, а исходная светлая — рецессивной. Бабочки, сохранившиеся в коллекциях энтомологов с середины прошлого века, имеют не столь темную окраску, как современные. Это указывает на то, что потемнение бабочек происходило постепенно, в череде поколений. Таким образом, преобразование генетического определения признаков фенотипа — это сложный и постепенный процесс, основанный на комбинировании генов под действием естественного отбора. Причины отбора — гибель или устранение от размножения
Естественный отбор не обязательно связан с гибелью менее приспособленных организмов. У многих насекомых, пауков, крабов и позвоночных существуют сложные ритуалы поведения, обуславливающие неслучайный выбор брачного партнера, место данного животного в иерархии главенства/подчинения в стае или стаде, распределение территории индивидуальных участков для выведения потомства.
 Все эти эколого-этологические системы направлены на повышение надежности размножения, а тем самым — на устранение от размножения тех особей, которые не способны конкурировать с элитой популяции, чья относительная приспособленность выше средней приспособленности особей данной популяции.
Все эти эколого-этологические системы направлены на повышение надежности размножения, а тем самым — на устранение от размножения тех особей, которые не способны конкурировать с элитой популяции, чья относительная приспособленность выше средней приспособленности особей данной популяции. Устранение от размножения не менее эффективно, чем гибель менее приспособленных организмов, так как неразмножающиеся особи не передают свои гены следующему поколению. Кроме того, дифференциальное размножение предоставляет животным два преимущества. Во-первых, селективность (избирательность) скрещиваний означает более всестороннюю «оценку» приспособленности. Для успеха размножения важно не только быть приспособленным по отношению к внешним для популяции экологическим факторам — успешно добывать пищу, избегать врагов и т. п., но и быть в этих отношениях лучше своих собратьев. Во-вторых, селективность скрещиваний уменьшает вредное для естественного отбора значение случайной гибели. Неразмножающаяся часть популяции, так называемый популяционный резерв, оказывается в худших условиях существования, чем репродуктивная часть популяции.
 Неразмножающиеся особи не имеют индивидуальных участков и вынуждены перемещаться по территории, становясь заметными для хищников. В стаях или стадах животные, занимающие низкое положение в иерархии, третируются более высокоранговыми особями, что вызывает у первых стресс и затрудняет им доступ к источникам пищи. В результате организмы, обладающие более низкой относительной приспособленностью, выполняют роль буфера, смягчающего неблагоприятные воздействия среды на репродуктивную часть популяции.
Неразмножающиеся особи не имеют индивидуальных участков и вынуждены перемещаться по территории, становясь заметными для хищников. В стаях или стадах животные, занимающие низкое положение в иерархии, третируются более высокоранговыми особями, что вызывает у первых стресс и затрудняет им доступ к источникам пищи. В результате организмы, обладающие более низкой относительной приспособленностью, выполняют роль буфера, смягчающего неблагоприятные воздействия среды на репродуктивную часть популяции. Механизмы устранения от размножения, то есть селективного скрещивания более приспособленных, видоспецифичны, а потому очень многообразны. В простейшем случае, названном Дарвином половым отбором (см. ПОЛОВОЙ ОТБОР), они приводят к выработке вторичных половых признаков (см. ВТОРИЧНЫЕ ПОЛОВЫЕ ПРИЗНАКИ). Яркая окраска самцов и сложные ритуалы ухаживания у многих птиц, гигантские размеры и драчливость самцов котиков и сивучей, кваканье лягушек и любые другие способы продемонстрировать самке свою привлекательность — результаты полового отбора.

Обычно устраняются от размножения молодые особи, часто даже после того, как они достигли половой зрелости. Например, у морских котиков половая зрелость наступает на втором году жизни, а к размножению секачи приступают на шестом году. Часть неразмножающихся молодых животных замещает постепенно уходящих из жизни иерархов, часть гибнет, а часть так и остается в популяционном резерве. Например, у ворон такой резерв составляет от 20 до 30% общей численности популяции. При этом старые вороны в возрасте 6—7 лет, из года в год занимающие свой гнездовой участок, лучше выкармливают птенцов, чем молодые 2—3-летние, только что занявшие свой гнездовой участок. Установлено, что старые вороны в Наурзумском заповеднике (Казахстан) приносят корм птенцам в среднем 64 раза в день, а молодые — не более 50 раз.
Пример с воронами показывает, насколько важна для естественного отбора не только та изменчивость, которая непосредственно детерминирована генетическими особенностями организмов, но и широта нормы реакции генотипов — возможности приобретения индивидуального опыта, сложные поведенческие реакции, физиологические процессы, при помощи которых организм адаптируется к изменениям условий среды, например, выдерживает голод или зимние холода.
Отбор по признакам с широкой нормой реакции
Норма реакции — диапазон изменений фенотипа при неизменном генотипе — важнейший способ адаптации организмов к изменениям условий их существования. Сами по себе изменения в пределах нормы реакции не наследуются, но способность к этим изменениям наследственно обусловлена. Например, яркость летнего загара у людей зависит от того, сколько времени человек провел на солнце, но смуглые брюнеты загорают быстрее и загар у них темнее, чем у светлокожих блондинов. Норма реакции, как и ее изменение, тоже вырабатывается под действием естественного отбора. Однако, механизм действия естественного отбора на подобные пластичные признаки фенотипа отличается от механизма его действия на признаки, жестко детерминированные генами, такие, например, как цвет крыльев у березовых пядениц. Если изменение условий существования не выходит за пределы нормы реакции большинства особей популяции, то организмы и приспосабливаются к нему в пределах своих индивидуальных возможностей, без изменения генетического состава популяции.
 Отбор вступает в действие несколько позже — он отбирает и комбинирует гены таким образом, чтобы обеспечить надежное, не зависящее ни от случайных колебаний среды, ни от случайных наследственных отклонений (мутаций), проявление нормы реакции в новых условиях.
Отбор вступает в действие несколько позже — он отбирает и комбинирует гены таким образом, чтобы обеспечить надежное, не зависящее ни от случайных колебаний среды, ни от случайных наследственных отклонений (мутаций), проявление нормы реакции в новых условиях. Одним из немногих исследованных примеров подобной адаптации может служить приспособление лесных мышей к жизни в высокогорьях Кавказа. Количество гемоглобина в крови у сухопутных млекопитающих соответствует парциальному давлению кислорода в местах их обитания. На равнине гемоглобина меньше, в горах, при пониженном атмосферном давлении, его становится больше. Количество гемоглобина — физиологический признак с широкой нормой реакции. Именно это свойство используют альпинисты, проходя адаптацию в горах перед высотными восхождениями. Через несколько дней пребывания в высокогорье количество гемоглобина в их крови увеличивается, что обеспечивает возможность подняться еще выше. То же самое происходит и с любыми другими млекопитающими при стойком понижении атмосферного давления.

Лесные мыши живут и на равнине, в Краснодарском крае (300 м над уровнем моря), и на высоте 1600—1700 м в горах. У равнинных мышей количество гемоглобина на 9—12 % ниже, чем у горных. При перемещении равнинных мышей в горы (или при помещении их в барокамеру) после 5—6 дней акклимации количество гемоглобина у них повышалось, но это повышение не превышало 7% и не достигало уровня, свойственного горным мышам. Аналогично, при переселении горных мышей на равнину, количество гемоглобина в их крови снижалось, но оставалось на 2—3% выше, чем у равнинных мышей. Другими словами, при освоении лесными мышами высокогорья у них произошел генетически детерминированный сдвиг нормы реакции по количеству гемоглобина в крови. Объяснить это изменение можно только естественным отбором на адаптацию к недостатку кислорода.
Формы естественного отбора
Естественный отбор далеко не всегда приводит к возникновению новых адаптаций. Достаточно часто он лишь сохраняет уже существующие приспособления организмов (и в этом случае действует как стабилизирующий отбор) или, меняя свое направление по сезонам года, в зависимости от циклических изменений состава пищи или врагов и паразитов, приводит не к выработке новых приспособлений, а только к циклическим изменениям состава популяций.

Например, российский биолог Н. В. Тимофеев-Ресовский (см. ТИМОФЕЕВ-РЕСОВСКИЙ Николай Владимирович) описал в Южной Германии циклические изменения состава популяций у двухточечной божьей коровки. В течение 10 лет каждую осень в этой популяции преобладали жуки с черной окраской спинки и надкрыльев, а каждую весну — с красной. Оказалось, что черные жуки быстрее размножаются, а красные более устойчивы к холоду во время зимовки. Благодаря большей смертности черных коровок зимой, весной красных становится примерно на 25% больше, а осенью — в среднем настолько же меньше. Действовал циклический отбор, менявший свое направление по сезонам года.
Если естественный отбор ответственен за формирование новых адаптивных признаков, он называется движущим. Однако, в постоянных условиях среды, когда признаки организмов сохраняют свое адаптивное значение, действует стабилизирующий отбор, направленный на поддержание в неизменном состоянии уже существующих приспособлений. Дело в том, что генетическая изменчивость, поскольку она не направлена, стремится разрушить приспособительные признаки и разрушает их в том случае, если по ним не идет отбор.
 Например, у всех диких млекопитающих окраска шерсти имеет защитное, маскирующее животное, или сигнальное значение. Только у домашних животных возникает пегая окраска — неправильное чередование пятен окрашенной и белой шерсти, как у пестрых коров. Домашним животным защитная окраска не нужна.
Например, у всех диких млекопитающих окраска шерсти имеет защитное, маскирующее животное, или сигнальное значение. Только у домашних животных возникает пегая окраска — неправильное чередование пятен окрашенной и белой шерсти, как у пестрых коров. Домашним животным защитная окраска не нужна. При стабилизирующем отборе отсекаются все сильные уклонения признаков, выживают особи, близкие по своим признакам к среднему их значению, нормальному для популяции. Такой отбор может поддерживать признак в течение миллионов лет.
Эффекты естественного отбора
Движущая форма естественного отбора приводит на основе генетической изменчивости к возникновению новых приспособлений организмов. В этом выражаются два главных результата его действия: накапливающий и преобразующий эффекты. Накапливающий эффект представляет собой усиление в череде поколений полезных организму признаков. Например, если жертва изначально крупнее нападающих на нее хищников, то дальнейшее увеличение размеров жертвы будет лучше защищать ее от нападений.
 Конечно, отбор на увеличение размеров может происходить только тогда, когда крупные жертвы, такие как титанотерий третичного периода или современные слоны, вполне обеспечены пищей. Но отбор на увеличение размеров и скорости роста жертв вызовет аналогичное направление отбора у хищников — они тоже станут более крупными. Это было характерно не только для третичных млекопитающих, но и для динозавров, например, тираннозавров (см. ТИРАННОЗАВРЫ)и бронтозавров (см. АПАТОЗАВРЫ). Накапливающий эффект движущего отбора проявляется не только по отношению к целостному организму, но и по отношению к отдельным органам. Увеличение размеров переднего мозга, прогрессивное развитие коры больших полушарий в ряду позвоночных — тоже пример накапливающего эффекта отбора.
Конечно, отбор на увеличение размеров может происходить только тогда, когда крупные жертвы, такие как титанотерий третичного периода или современные слоны, вполне обеспечены пищей. Но отбор на увеличение размеров и скорости роста жертв вызовет аналогичное направление отбора у хищников — они тоже станут более крупными. Это было характерно не только для третичных млекопитающих, но и для динозавров, например, тираннозавров (см. ТИРАННОЗАВРЫ)и бронтозавров (см. АПАТОЗАВРЫ). Накапливающий эффект движущего отбора проявляется не только по отношению к целостному организму, но и по отношению к отдельным органам. Увеличение размеров переднего мозга, прогрессивное развитие коры больших полушарий в ряду позвоночных — тоже пример накапливающего эффекта отбора. Преобразующий эффект, или как его называли в конце 19 — начале 20 века, творческая роль естественного отбора, состоит в том, что, изменяя признаки в соответствии с изменениями условий среды — усиливая полезные и ослабляя признаки, утратившие приспособительное значение, естественный отбор преобразует организацию потомков по сравнению с предками, создавая новые виды.
 Один из основателей популяционной генетики, американский ученый С. Райт (см. РАЙТ Сьюалл) точно охарактеризовал творческую роль естественного отбора, сказав, что он из хаоса изменчивости создает новую организацию.
Один из основателей популяционной генетики, американский ученый С. Райт (см. РАЙТ Сьюалл) точно охарактеризовал творческую роль естественного отбора, сказав, что он из хаоса изменчивости создает новую организацию. Генетическая изменчивость, проявляясь в фенотипе, меняет отдельные признаки, а в процессе борьбы за существование гибнут, устраняются от размножения или, напротив, выживают и размножаются целостные организмы. Поэтому, несмотря на то, что отбор идет по признакам фенотипа, каждый раз «оценивается» приспособленность особи, отличающейся от других особей популяции именно своими признаками. Поэтому творческая роль отбора выражается именно в преобразовании всей организации особей.
Кроме накапливающего и преобразующего, естественному отбору свойственны также поддерживающий и распределяющий эффекты.
Поддерживающий эффект выражается в том, что приспособленность организмов, подвергающихся естественному отбору, не может уменьшаться. Она может увеличиваться или оставаться на том же уровне, что и у предков.
 Сказанное не означает, что все признаки обязательно должны обладать приспособительным значением, быть полезными организму. Рудиментарные органы, например, остаток костей тазового пояса у усатых китов, таковыми не обладают. Иногда у животных возникают признаки не только бесполезные, но и вредные (см. Адаптация (см. АДАПТАЦИЯ (в биологии))).
Сказанное не означает, что все признаки обязательно должны обладать приспособительным значением, быть полезными организму. Рудиментарные органы, например, остаток костей тазового пояса у усатых китов, таковыми не обладают. Иногда у животных возникают признаки не только бесполезные, но и вредные (см. Адаптация (см. АДАПТАЦИЯ (в биологии))). Распределяющий эффект отбора состоит в том, что в более благоприятных условиях среды организмы чаще выживают и оставляют потомство. В менее благоприятных это происходит реже. В результате отбор распределяет организмы данного вида в пределах его ареала. Популяция — элементарная единица эволюции — сама формируется под действием естественного отбора.
Таким образом, естественный отбор — не единственный, но важнейший движущий фактор эволюции. Все биологическое разнообразие, составляющее биосферу Земли, сформировалось в результате естественного отбора.
Естественный отбор, генетический дрейф и поток генов не действуют изолированно в природных популяциях
Естественный отбор, генетический дрейф и поток генов — это механизмы, вызывающие изменения частот аллелей с течением времени. Когда одна или несколько из этих сил действуют в популяции, популяция нарушает предположения Харди-Вайнберга, и происходит эволюция. Таким образом, теорема Харди-Вайнберга обеспечивает нулевую модель для изучения эволюции, и основное внимание популяционной генетики уделяется пониманию последствий нарушения этих предположений.
Когда одна или несколько из этих сил действуют в популяции, популяция нарушает предположения Харди-Вайнберга, и происходит эволюция. Таким образом, теорема Харди-Вайнберга обеспечивает нулевую модель для изучения эволюции, и основное внимание популяционной генетики уделяется пониманию последствий нарушения этих предположений.
Естественный отбор имеет место, когда особи с определенными генотипами имеют больше шансов, чем особи с другими генотипами, выжить и размножиться и, таким образом, передать свои аллели следующему поколению. Как утверждал Чарльз Дарвин (1859 г.) в книге «О происхождении видов» , естественный отбор должен происходить при соблюдении следующих условий:
- Среди особей внутри популяции существуют различия по некоторым признакам.
- Эта вариация является наследственной (т. е. существует генетическая основа вариации, так что потомство имеет тенденцию напоминать своих родителей по этому признаку).
- Изменчивость по этому признаку связана с изменчивостью приспособленности (среднее чистое воспроизводство особей с данным генотипом по отношению к особям с другими генотипами).


Направленный отбор со временем приводит к увеличению частоты предпочтительного аллеля. Рассмотрим три генотипа ( АА , Аа и аа ), которые различаются по приспособленности, так что особи АА производят в среднем больше потомства, чем особи других генотипов. В этом случае, если предположить, что селективный режим остается постоянным и что действие отбора является единственным нарушением предположений Харди-Вайнберга, Аллель становился все более распространенным с каждым поколением и в конечном итоге закреплялся в популяции. Скорость, с которой полезный аллель приближается к фиксации, частично зависит от доминантных отношений между аллелями в рассматриваемом локусе (Рис. 1). Начальное увеличение частоты редкого, благоприятного, доминантного аллеля происходит быстрее, чем редкого, благоприятного, рецессивного аллеля, потому что редкие аллели обнаруживаются в основном у гетерозигот. Таким образом, новая рецессивная мутация не может быть «увидена» естественным отбором до тех пор, пока она не достигнет достаточно высокой частоты (возможно, из-за случайных эффектов дрейфа генов — см. ниже), чтобы начать появляться у гомозигот. Однако новая доминантная мутация сразу становится видимой для естественного отбора, потому что ее влияние на приспособленность наблюдается у гетерозигот. Как только полезный аллель достигает высокой частоты, вредные аллели обязательно редки и, таким образом, в основном присутствуют у гетерозигот, так что окончательный подход к фиксации является более быстрым для благоприятного рецессивного аллеля, чем для благоприятного доминантного аллеля. Как следствие, естественный отбор не так эффективен, как можно было бы наивно ожидать, в устранении вредных рецессивных аллелей из популяций.
ниже), чтобы начать появляться у гомозигот. Однако новая доминантная мутация сразу становится видимой для естественного отбора, потому что ее влияние на приспособленность наблюдается у гетерозигот. Как только полезный аллель достигает высокой частоты, вредные аллели обязательно редки и, таким образом, в основном присутствуют у гетерозигот, так что окончательный подход к фиксации является более быстрым для благоприятного рецессивного аллеля, чем для благоприятного доминантного аллеля. Как следствие, естественный отбор не так эффективен, как можно было бы наивно ожидать, в устранении вредных рецессивных аллелей из популяций.
Рисунок 1: Изменение частоты аллелей при направленном отборе в пользу (а) доминантного полезного аллеля и (б) рецессивного благоприятного аллеля
Балансирующий отбор, в отличие от направленного отбора, поддерживает генетический полиморфизм в популяциях. Например, если гетерозиготы в локусе имеют более высокую приспособленность, чем гомозиготы (сценарий, известный как преимущество или сверхдоминирование гетерозигот), естественный отбор будет поддерживать множественные аллели на стабильных равновесных частотах.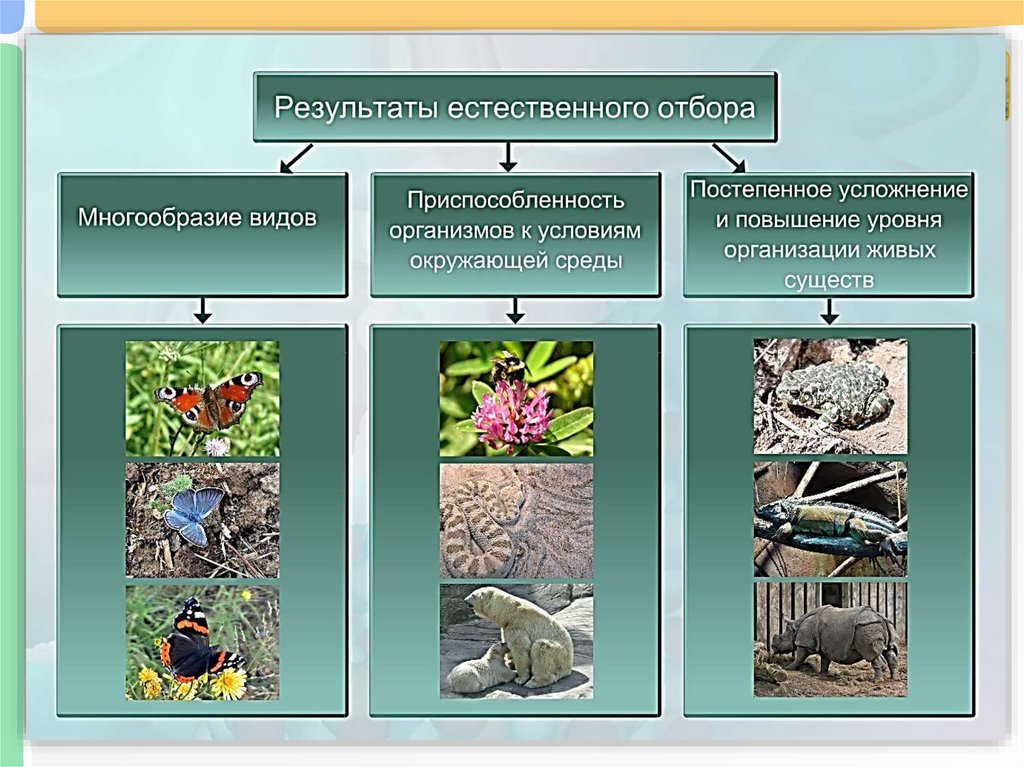 Стабильный полиморфизм также может сохраняться в популяции, если приспособленность, связанная с генотипом, снижается по мере увеличения частоты этого генотипа (т. е. если существует отрицательный частотно-зависимый отбор). Важно отметить, что недостаток гетерозигот (недостаточное доминирование) и положительный частотно-зависимый отбор также могут действовать на локус, но ни один из них не поддерживает множественные аллели в популяции, и, таким образом, ни один из них не является формой уравновешивающего отбора.
Стабильный полиморфизм также может сохраняться в популяции, если приспособленность, связанная с генотипом, снижается по мере увеличения частоты этого генотипа (т. е. если существует отрицательный частотно-зависимый отбор). Важно отметить, что недостаток гетерозигот (недостаточное доминирование) и положительный частотно-зависимый отбор также могут действовать на локус, но ни один из них не поддерживает множественные аллели в популяции, и, таким образом, ни один из них не является формой уравновешивающего отбора.
Генетический дрейф возникает из-за ошибки выборки, присущей передаче гамет особями в ограниченной популяции. Пул гамет популяции в поколении t — это общий пул яйцеклеток и сперматозоидов, произведенных особями в этом поколении. Если бы пул гамет был бесконечен по размеру и если бы не было отбора или мутации, действующих в локусе с двумя аллелями ( А и А ), мы могли бы ожидать, что доля гамет, содержащих аллель А, точно равна частоте из A , а доля гамет, содержащих a , равна частоте a . Сравните эту ситуацию с подбрасыванием правильной монеты. Если бы вы подбрасывали монету бесконечное количество раз, доля выпадения орла была бы 0,50, а доля решки — 0,50. Однако если вы подбросите монету всего 10 раз, не стоит слишком удивляться, если выпадет 7 орлов и 3 решки. Это отклонение от ожидаемых частот начала и окончания связано с ошибкой выборки. Чем больше раз вы подбрасываете монету, тем ближе эти частоты должны приближаться к 0,50, поскольку ошибка выборки уменьшается по мере увеличения размера выборки.
Сравните эту ситуацию с подбрасыванием правильной монеты. Если бы вы подбрасывали монету бесконечное количество раз, доля выпадения орла была бы 0,50, а доля решки — 0,50. Однако если вы подбросите монету всего 10 раз, не стоит слишком удивляться, если выпадет 7 орлов и 3 решки. Это отклонение от ожидаемых частот начала и окончания связано с ошибкой выборки. Чем больше раз вы подбрасываете монету, тем ближе эти частоты должны приближаться к 0,50, поскольку ошибка выборки уменьшается по мере увеличения размера выборки.
В конечной популяции взрослые особи в поколении t передадут конечное число гамет, чтобы произвести потомство в поколении t + 1. Частоты аллелей в этом пуле гамет обычно отклоняются от частот популяции в поколение t из-за ошибки выборки (опять же, если предположить, что в локусе нет выбора). Таким образом, частоты аллелей в этой популяции со временем будут меняться из-за случайных событий, то есть популяция будет подвергаться генетическому дрейфу. Чем меньше численность населения ( N ), тем важнее эффект генетического дрейфа. На практике при моделировании эффектов дрейфа мы должны учитывать эффективный размер популяции ( N e ), который по существу представляет собой количество размножающихся особей и может отличаться от учетного размера, N , при различных сценариях. включая неравное соотношение полов, определенные структуры спаривания и временные колебания численности популяции.
Чем меньше численность населения ( N ), тем важнее эффект генетического дрейфа. На практике при моделировании эффектов дрейфа мы должны учитывать эффективный размер популяции ( N e ), который по существу представляет собой количество размножающихся особей и может отличаться от учетного размера, N , при различных сценариях. включая неравное соотношение полов, определенные структуры спаривания и временные колебания численности популяции.
В локусе с множественными нейтральными аллелями (аллелями, идентичными по своему влиянию на приспособленность) генетический дрейф приводит к фиксации одного из аллелей в популяции и, таким образом, к потере других аллелей, так что гетерозиготность в популяции спадает до нуля. В любой момент времени вероятность того, что один из этих нейтральных аллелей в конечном итоге закрепится, равна частоте этого аллеля в популяции. Мы можем рассматривать эту проблему с точки зрения множества повторяющихся популяций, каждая из которых представляет дему (субпопуляцию) внутри метапопуляции (набора дем). Учитывая 10 конечных демов, равных N e , каждая из которых имеет начальную частоту аллеля A , равную 0,5, мы ожидаем возможную фиксацию A в 5 демах и возможную потерю A в 5 демах. Наши наблюдения, вероятно, в некоторой степени отклоняются от этих ожиданий, поскольку мы рассматриваем конечное число демов (рис. 2). Таким образом, дрейф генов устраняет генетическую изменчивость внутри демов, но приводит к дифференциации между демами полностью за счет случайных изменений частот аллелей.
Учитывая 10 конечных демов, равных N e , каждая из которых имеет начальную частоту аллеля A , равную 0,5, мы ожидаем возможную фиксацию A в 5 демах и возможную потерю A в 5 демах. Наши наблюдения, вероятно, в некоторой степени отклоняются от этих ожиданий, поскольку мы рассматриваем конечное число демов (рис. 2). Таким образом, дрейф генов устраняет генетическую изменчивость внутри демов, но приводит к дифференциации между демами полностью за счет случайных изменений частот аллелей.
Рисунок 2: Моделирование изменения частоты аллеля в 10 повторных популяциях (N = 20)
Поскольку начальная частота аллеля A = 0,5, мы ожидаем, что A будет зафиксирован в 5 популяциях и потерян в 5 популяциях, но наши наблюдения отклоняются от ожиданий из-за конечного числа популяций. В этом цикле симуляций мы видим 7 случаев фиксации (p = 1), 2 случая потери (p = 0) и один случай, в котором остаются два аллеля после 100 поколений. В этой последней популяции A в конечном итоге достигнет фиксации или потери.
Поток генов — это перемещение генов в популяцию или из нее. Такое перемещение может быть связано с миграцией отдельных организмов, размножающихся в своих новых популяциях, или с перемещением гамет (например, вследствие переноса пыльцы между растениями). В отсутствие естественного отбора и генетического дрейфа поток генов приводит к генетической однородности среди демов в метапопуляции, так что для данного локуса частоты аллелей достигают равновесных значений, равных средним частотам в метапопуляции. Напротив, ограниченный поток генов способствует дивергенции популяций за счет отбора и дрейфа, которые, если они сохраняются, могут привести к видообразованию.
Естественный отбор, генетический дрейф и поток генов не действуют изолированно, поэтому мы должны рассмотреть, как взаимодействие этих механизмов влияет на эволюционные траектории в природных популяциях. Этот вопрос имеет решающее значение для специалистов по природоохранной генетике, которые борются с последствиями этих эволюционных процессов, разрабатывая заповедники и моделируя динамику популяций находящихся под угрозой исчезновения видов в фрагментированных средах обитания. Все реальные популяции конечны и, следовательно, подвержены влиянию генетического дрейфа. Мы ожидаем, что в бесконечной популяции направленный отбор в конце концов зафиксирует выгодный аллель, но это не обязательно произойдет в конечной популяции, потому что эффекты дрейфа могут преодолеть эффекты отбора, если отбор слаб и/или популяция мала. Потеря генетической изменчивости из-за дрейфа вызывает особую озабоченность в небольших популяциях, находящихся под угрозой исчезновения, в которых фиксация вредных аллелей может снизить жизнеспособность популяции и повысить риск исчезновения. Даже если усилия по сохранению увеличат рост популяции, низкая гетерозиготность, вероятно, сохранится, поскольку узкие места (периоды сокращения численности популяции) оказывают более выраженное влияние на Ne, чем периоды большей численности популяции.
Все реальные популяции конечны и, следовательно, подвержены влиянию генетического дрейфа. Мы ожидаем, что в бесконечной популяции направленный отбор в конце концов зафиксирует выгодный аллель, но это не обязательно произойдет в конечной популяции, потому что эффекты дрейфа могут преодолеть эффекты отбора, если отбор слаб и/или популяция мала. Потеря генетической изменчивости из-за дрейфа вызывает особую озабоченность в небольших популяциях, находящихся под угрозой исчезновения, в которых фиксация вредных аллелей может снизить жизнеспособность популяции и повысить риск исчезновения. Даже если усилия по сохранению увеличат рост популяции, низкая гетерозиготность, вероятно, сохранится, поскольку узкие места (периоды сокращения численности популяции) оказывают более выраженное влияние на Ne, чем периоды большей численности популяции.
Мы уже видели, что генетический дрейф ведет к дифференциации демов внутри метапопуляции. Если принять простую модель, в которой индивидуумы имеют равные вероятности расселения среди всех демов (каждый из которых имеет эффективный размер N и ) внутри метапопуляции, то скорость миграции ( м ) представляет собой долю копий гена в пределах дема, введенная через иммиграцию в поколение. Согласно общепринятому приближению, интродукция только одного мигранта на поколение ( N e m = 1) составляет достаточный поток генов, чтобы противодействовать диверсифицирующим эффектам генетического дрейфа в метапопуляции.
Согласно общепринятому приближению, интродукция только одного мигранта на поколение ( N e m = 1) составляет достаточный поток генов, чтобы противодействовать диверсифицирующим эффектам генетического дрейфа в метапопуляции.
Естественный отбор может производить генетическую изменчивость среди демов в пределах метапопуляции, если в разных демах преобладает различное давление отбора. Если N e достаточно велико, чтобы не учитывать влияние генетического дрейфа, то мы ожидаем, что направленный отбор закрепит предпочтительный аллель в пределах данной фокальной демы. Однако постоянное введение посредством генного потока аллелей, полезных в других демах, но вредных в фокальном деме, может противодействовать эффектам отбора. В этом сценарии вредоносный аллель останется на промежуточной равновесной частоте, которая отражает баланс между потоком генов и естественным отбором.
Естественный отбор
Английский натуралист Чарльз Дарвин разработал идею естественного отбора после пятилетнего путешествия для изучения растений, животных и окаменелостей в Южной Америке и на островах Тихого океана. В 1859 году он довел до сведения всего мира идею естественного отбора в своем бестселлере « О происхождении видов ».
В 1859 году он довел до сведения всего мира идею естественного отбора в своем бестселлере « О происхождении видов ».
Естественный отбор — это процесс адаптации и изменения популяций живых организмов. Люди в популяции естественно изменчивы, а это означает, что все они в чем-то разные. Эта вариация означает, что некоторые люди обладают чертами, более подходящими для окружающей среды, чем другие. Особи с адаптивными чертами — чертами, дающими им некоторое преимущество, — с большей вероятностью выживут и размножатся. Затем эти особи передают адаптивные черты своим потомкам. Со временем эти полезные черты становятся более распространенными в популяции. Благодаря этому процессу естественного отбора благоприятные черты передаются из поколения в поколение.
Естественный отбор может привести к видообразованию, когда один вид дает начало новому и совершенно другому виду. Это один из процессов, который движет эволюцией и помогает объяснить разнообразие жизни на Земле.
Дарвин выбрал название «естественный отбор», чтобы противопоставить его «искусственному отбору» или селекционному размножению, контролируемому людьми. Он указывал на разведение голубей, популярное в его время хобби, как на пример искусственного отбора. Выбирая, какие голуби спариваться с другими, любители создали разные породы голубей с причудливыми перьями или акробатическим полетом, которые отличались от диких голубей.
Он указывал на разведение голубей, популярное в его время хобби, как на пример искусственного отбора. Выбирая, какие голуби спариваться с другими, любители создали разные породы голубей с причудливыми перьями или акробатическим полетом, которые отличались от диких голубей.
Дарвин и другие ученые того времени утверждали, что процесс, очень похожий на искусственный отбор, происходил в природе без какого-либо вмешательства человека. Он утверждал, что естественный отбор объясняет, как большое разнообразие форм жизни развилось с течением времени от одного общего предка.
Дарвин не знал, что гены существуют, но он мог видеть, что многие признаки являются наследственными — передаются от родителей к потомству.
Мутации — это изменения в структуре молекул, из которых состоят гены, называемые ДНК. Мутация генов является важным источником генетической изменчивости в популяции. Мутации могут быть случайными (например, когда реплицирующиеся клетки делают ошибку при копировании ДНК) или происходить в результате воздействия чего-то в окружающей среде, например вредных химических веществ или радиации.