Рецептор | Мир Психологии
РЕЦЕПТОР
Словарь практического психолога. С.Ю. Головин
Рецептор — нервные образования, служащие для превращения световой, механической, химической, термической энергии агентов среды внешней и внутренней в импульсы нервные. Периферические специализованные части анализаторов, через которые лишь определенный вид энергии трансформируется в процесс возбуждения нервного. Рецепторы широко варьируют по сложности структуры и уровню приспособленности к своей функции. Отдельные рецепторы анатомически связаны друг с другом и образуют рецептивные поля, способные перекрываться. В зависимости от энергии соответственного раздражения рецепторы делятся на механорецепторы и хеморецепторы.
- Механорецепторы обнаружены в ухе, вестибулярном аппарате, мышцах, суставах, в коже и внутренних органах.
- Хеморецепторы обслуживают обонятельную и вкусовую чувствительность; многие из них находятся в мозге, реагируя на изменения химического состава жидкой среды организма.


Зрительные рецепторы, по существу, тоже являются хеморецепторами. Иногда также выделяются терморецепторы, фоторецепторы и электрорецепторы.В зависимости от положения в организме и выполняемой функции выделяются:
- экстероцепторы — сюда относятся рецепторы дистантные, получавшие информацию на некоим расстоянии от источника раздражения — обонятельные, слуховые, зрительные, вкусовые;
- интероцепторы — сигнализируют о раздражителях внутренней среды;
- проприоцепторы — сигнализируют о состоянии системы двигательной организма.
Словарь психиатрических терминов. В.М. Блейхер, И.В. Крук
нет значения и толкования слова
Неврология. Полный толковый словарь. Никифоров А.С.
Рецептор — специализированная нервная структура с особенно высокой степенью раздражимости, способная воспринимать раздражение и трансформировать его в биоэлектрический потенциал – нервный импульс. Обладает специфичностью к определенным раздражителям, которая определяет строение рецептора и место его расположения (экстерорецептор, проприорецептор, интерорецептор).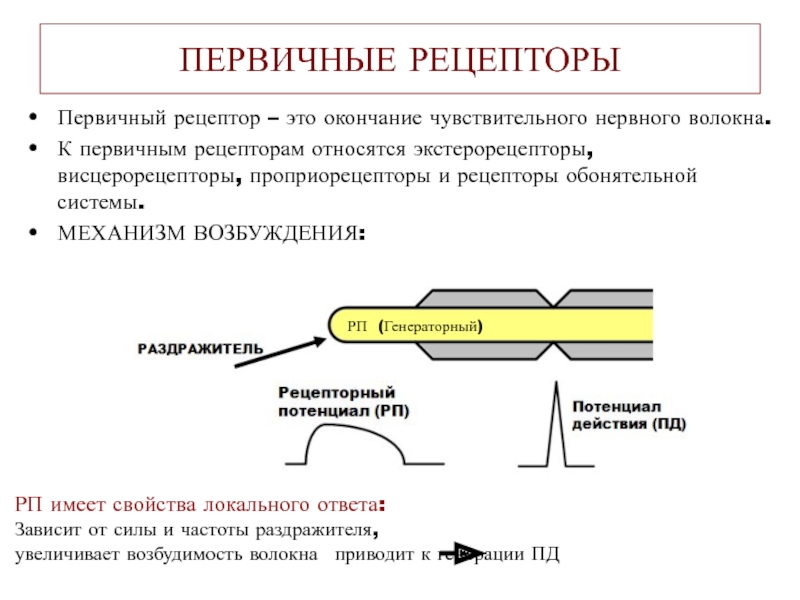
Оксфордский толковый словарь по психологии
Рецептор — специализированная нервная летка или ее часть, которая преобразует физические стимулы в потенциалы рецептора. То есть клетка, чувствительная к определенной форме стимуляции и надежно претерпевающая определенную модель изменений. Такое определение достаточно широко для всего, о чем говорится ниже и что следует отнести к рецепторам,
- Периферийные клетки в различных сенсорных системах, которые реагируют на определенные формы физической энергии, например, палочки и колбочки в сетчатке, волосковые клетки в кортиевом органе внутреннего уха, чувствительные к давлению клетки в коже, вкусовые сосочки на языке и т.д.
- Проприорецепторы, которые реагируют на внешнюю стимуляцию, например, волосковые клетки в полукруглых каналах утреннего уха, рецепторы растяжения во внутренних органах, кинестетические рецепторы в суставах и сухожилиях и т.д.
- Постсинаптические нейроны, которые реагируют на высвобождение нейротрансмиттерных веществ в нерв системе; см. здесь рецепторное место.
 здесь рецепторное место.
здесь рецепторное место.За последние годы использовалось несколько систем классификации рецепторов. Некоторые из них основываются на локализации рецепторов в теле, например, экстероцепторы, интероцепторы и проприоцепторы. Некоторые основываются на определенной обслуживаемой модальности, например, зрительные рецепторы, слуховые рецепторы и т.д. Некоторые зависят от определения формы физических стимулов, к которым чувствительны рецепторы, например, химические рецепторы, типа тех, которые обслуживают вкус и запах, механические рецепторы для давления и слуха, световые рецепторы в зрении, температурные рецепторы для тепла и холода и т.д. Другие системы ориентиру на вещества-нейротрансмиттеры, связывающие нервные пути, обслуживающие определенную систему рецепторов, например, холинергические рецепторы, паминэргические рецепторы и т.д.
Обратите внимание, что эта последняя с тема классификации строится на анализе скорее центральной нервной системы, чем определенных сенсорных систем, инициирующих нервные измене Обычно контекст, в котором обсуждаются определенные рецепторы, понять, какая система классификации используется.
предметная область термина
назад в раздел : словарь терминов / глоссарий / таблица
Рецепторы. Основы общей психологии
Рецепторы
Рецептор — орган, специально приспособленный для рецепции раздражений, легче, чем прочие органы или нервные волокна, поддается раздражению; он отличается особенно низкими порогами раздражения, т. е. его чувствительность, обратно пропорциональная порогу, особенно высока. В этом первая особенность рецептора как специализированного аппарата: обладая особенно большой чувствительностью, он специально приспособлен для рецепции раздражений.
При этом рецепторы приспособлены для рецепции не любых раздражителей. Каждый рецептор специализируется применительно к определенному раздражителю. <…> Так, образуются тангорецепторы, приспособленные к рецепции прикосновения, густорецепторы — для рецепции вкусовых раздражении, стиборецепторы — для обонятельных, приспособленные для рецепции звука и света фоно— и фоторецепторы.
Таким образом, специальная приспособленность к рецепции раздражений, выражающаяся в особо высокой чувствительности, — во-первых, и приспособленность к рецепции специальных раздражителей, т. е. специализация рецепторов по виду раздражителей, — во-вторых, составляют основные черты, характеризующие рецепторный аппарат.
В парадоксальной форме специализация органов чувств, или рецепторов, выражается в том, что и неадекватный раздражитель, воздействуя на определенный рецептор, может вызвать специфические для него ощущения. Так, сетчатка дает световые ощущения при воздействии на нее как светом, так и электрическим током или давлением («искры из глаз сыплются» при ударе). Но и механический раздражитель может дать ощущение давления, звука или света в зависимости от того, воздействует ли он на осязание, слух или зрение. <…> Основываясь на этих фактах и опираясь на специализацию «органов чувств», Й. Мюллер выдвинул свой принцип специфической энергии органов чувств. Основу его составляет бесспорное положение, заключающееся в том, что все специфицированные ощущения находятся в определенном соотношении с гистологически специфицированными органами, их обусловливающими.
На этой основе Мюллер выдвигает другую идею, согласно которой ощущение зависит не от природы раздражителя, а от органа или нерва, в котором происходит процесс раздражения, и является выражением его специфической энергии. Посредством зрения, например, по Мюллеру, познается несуществующий во внешнем мире свет, поскольку глаз наш доставляет впечатление света и тогда, когда на него действует электрический или механический раздражитель, т. е. в отсутствие физического света. Ощущение света признается выражением специфической энергии сетчатки: оно — лишь субъективное состояние сознания. Включение физиологических процессов в соответствующем аппарате в число объективных, опосредующих условий ощущения превращается, таким образом, в средство отрыва ощущения от его внешней причины и признания субъективности ощущения. 62 Из связи субъекта с объектом ощущение превращается во включенную между субъектом и объектом завесу.
Стоит подойти к интерпретации того позитивного фактического положения, которое лежит в основе субъективно-идеалистической надстройки, возведенной над нею Мюллером, чтобы те же факты предстали в совсем ином освещении. В процессе биологической эволюции сами органы чувств формировались в реальных взаимоотношениях организма со средой, под воздействием внешнего мира. Специализация органов чувств совершалась под воздействием внешних раздражителей; воздействие внешнего мира формирует сами рецепторы. Рецепторы являются как бы анатомически закрепленными в строении нервной системы отпечатками эффектов процессов раздражения. Нужно, собственно, говорить не столько о специфической энергии органов чувств, сколько об органах чувств специфической энергии. «Специфическая энергия» органов чувств или нервов, взятая в генетическом плане, выражает, таким образом, пластичность нерва по отношению к специфичности внешнего раздражителя. Источники специфичности нужно первично искать не внутри, а во вне. Она свидетельствует не о субъективности ощущения, а об его объективности.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРесЧувствительность к андрогенам: за пределами общеизвестного | Хрипун
Действие андрогенов затрагивает практически все органы и системы. Формирование половых признаков в процессе эмбриогенеза, становление и функционирование репродуктивной системы и обеспечение фертильности — это лишь малая часть эффектов тестостерона (Т) и его метаболитов. Гораздо шире внегонадное действие андрогенов, представленное их влиянием на сердечно-сосудистую систему, липидный и углеводный обмены, минерализацию костной ткани, стимуляцию кроветворения, формирование и функционирование когнитивных функций, психосоциальную адаптацию и сексуальность. Одним из наиболее важных вопросов клинической практики является различная выраженность симптомов гипогонадизма, а также разный ответ на заместительную терапию у пациентов с одинаковым уровнем сывороточного Т. Ответ на этот вопрос кроется в определении чувствительности к андрогенам и является на сегодняшний день крайне малоизученным. Данный обзор содержит анализ наиболее актуальных исследований, опубликованных в рецензируемых журналах баз данных PubMed и eLibrary.
Формирование половых признаков в процессе эмбриогенеза, становление и функционирование репродуктивной системы и обеспечение фертильности — это лишь малая часть эффектов тестостерона (Т) и его метаболитов. Гораздо шире внегонадное действие андрогенов, представленное их влиянием на сердечно-сосудистую систему, липидный и углеводный обмены, минерализацию костной ткани, стимуляцию кроветворения, формирование и функционирование когнитивных функций, психосоциальную адаптацию и сексуальность. Одним из наиболее важных вопросов клинической практики является различная выраженность симптомов гипогонадизма, а также разный ответ на заместительную терапию у пациентов с одинаковым уровнем сывороточного Т. Ответ на этот вопрос кроется в определении чувствительности к андрогенам и является на сегодняшний день крайне малоизученным. Данный обзор содержит анализ наиболее актуальных исследований, опубликованных в рецензируемых журналах баз данных PubMed и eLibrary.
Структура и функции андрогенного рецептора
Многогранное действие Т реализуется через андрогенные рецепторы (AR), расположенные внутриклеточно и структурно относящиеся к рецепторам стероидных гормонов. AR присутствуют в клетках семенников, простаты, кожи, нервной системы и других тканей. Генетический контроль за активностью этих рецепторов осуществляется геном AR, который и обусловливает чувствительность к андрогенам и реализацию действия Т. Широта и разносторонность эффектов, реализуемых через ген AR, впечатляют — от дифференцировки пола в эмбриогенезе, регуляции сперматогенеза, прогрессирования рака и доброкачественной гиперплазии предстательной железы (ДГПЖ), симптоматики гипогонадизма до влияния на углеводный и липидный обмены, минеральную плотность костной ткани, эндотелий сосудов и даже психосоциальные аспекты личности.
AR присутствуют в клетках семенников, простаты, кожи, нервной системы и других тканей. Генетический контроль за активностью этих рецепторов осуществляется геном AR, который и обусловливает чувствительность к андрогенам и реализацию действия Т. Широта и разносторонность эффектов, реализуемых через ген AR, впечатляют — от дифференцировки пола в эмбриогенезе, регуляции сперматогенеза, прогрессирования рака и доброкачественной гиперплазии предстательной железы (ДГПЖ), симптоматики гипогонадизма до влияния на углеводный и липидный обмены, минеральную плотность костной ткани, эндотелий сосудов и даже психосоциальные аспекты личности.
Ген AR локализован на длинном плече Х-хромо сомы и состоит из 8 экзонов, кодирующих 3 структурно-функциональных белковых домена. N-конце вой домен участвует в гомодимеризации рецептора и связывании с ко-активаторами или ко-репрессорами и представлен экзоном 1. Второй — ДНК-связы вающий домен содержит петлевой участок из двух «цинковых пальцев» (экзоны 2 и 3). С-концевой домен обеспечивает связывание со стероидными гормонами и представлен экзонами 4—8 [1].
С-концевой домен обеспечивает связывание со стероидными гормонами и представлен экзонами 4—8 [1].
В последние годы внимание исследователей особенно сосредоточено на изучении первого экзона гена AR, для которого характерны повторы последовательности из трех нуклеотидов — цитозина, аденина и гуанина — CAG. Триплет CAG кодирует аминокислоту глутамин, поэтому от числа тринуклеотидных повторов зависит количество глутамина в белке. Так, меньшему числу CAG-повторов соответствует меньшая степень конформационных изменений рецептора, что делает связь гормона и рецептора прочнее и приводит к повышению транскрипционной активности AR. Напротив, с увеличением числа CAG-повторов связь между андрогенами и рецептором становится слабее и транскрипционная активность AR снижается. Таким образом, чем больше длина CAG-повторов, тем слабее оказывается эффект андрогенов при одинаковым уровне Т [2]. Слабая активность андрогенов, вызванная удлинением триплета, находит отражение в механизме обратной связи в пределах гипоталамо-гипофизарно-гонадной оси. У здоровых мужчин удлинение CAG-повторов сопровождается усилением секреции лютеинизирующего гормона (ЛГ) [3], что при интактных клетках Лейдига вызывает повышенную продукцию Т и, следовательно, компенсирует ослабление действия андрогенов. Результатом гипертестостеронемии будет повышенная ароматизация Т в эстрадиол, концентрация которого возрастет. По-видимому, именно с этим связана гинекомастия у пациентов с увеличенным числом CAG-повторов [4]. Высокие концентрации эстрадиола у мужчин с удлинением CAG-по второв и сохраненной обратной связью в оси гипоталамус-гипофиз-гонады может усиливать эффекты этих гормонов в эстроген-зависимых тканях, например в костной [3]. Рядом исследователей обнаружена отрицательная корреляция между длиной CAG—повто ров и минеральной плотностью костной ткани [5—7]. Эта связь прослеживалась и у молодых субъектов, но с увеличением возраста мужчин негативное влияние полиморфизма гена AR на минеральную плотность костной ткани усиливалось.
У здоровых мужчин удлинение CAG-повторов сопровождается усилением секреции лютеинизирующего гормона (ЛГ) [3], что при интактных клетках Лейдига вызывает повышенную продукцию Т и, следовательно, компенсирует ослабление действия андрогенов. Результатом гипертестостеронемии будет повышенная ароматизация Т в эстрадиол, концентрация которого возрастет. По-видимому, именно с этим связана гинекомастия у пациентов с увеличенным числом CAG-повторов [4]. Высокие концентрации эстрадиола у мужчин с удлинением CAG-по второв и сохраненной обратной связью в оси гипоталамус-гипофиз-гонады может усиливать эффекты этих гормонов в эстроген-зависимых тканях, например в костной [3]. Рядом исследователей обнаружена отрицательная корреляция между длиной CAG—повто ров и минеральной плотностью костной ткани [5—7]. Эта связь прослеживалась и у молодых субъектов, но с увеличением возраста мужчин негативное влияние полиморфизма гена AR на минеральную плотность костной ткани усиливалось.
Для мужчин европейской популяции нормальным считается количество CAG-повторов от 9 до 37. Однако в этом нормативе существуют популяционные различия. Так, среднее число CAG-повторов в гене AR у мужчин-европейцев составляет 21, у африканцев — 17, у азиатов — 23. При этом, например 25 CAG-повторов хотя и укладывается в нормативные значения, зачастую бывает связано с ослаблением действия андрогенов, проявляющимся соответствующей клинической симптоматикой [8].
Эмбриогенез и дифференцировка пола
Клеточный каскад нормальной дифференциации мужского пола также моделируется молекулярным взаимодействием Т и дигидротестостерона (ДГТ) с AR в андроген-зависимых тканях-мишенях. От этого взаимодействия еще на 7—8-й неделе эмбриогенеза зависит правильность формирования мужского пола. Наиболее ярким примером участия AR в формировании пола является синдром тестикулярной феминизации, возникающий вследствие мутации гена AR с развитием нечувствительности к андрогенам [1]. У эмбриона с генотипом 46,XY при выпадении эффектов Т и ДГТ бипотенциальные закладки внутренних и наружных половых органов развиваются по базовому пути с формированием женского фенотипа. Такие пациенты в постпубертатном периоде зачастую имеют достаточно развитые молочные железы и распределение жировой ткани по женскому типу, что связано с ароматизацией синтезирующихся в яичках андрогенов в эстрогены.
У эмбриона с генотипом 46,XY при выпадении эффектов Т и ДГТ бипотенциальные закладки внутренних и наружных половых органов развиваются по базовому пути с формированием женского фенотипа. Такие пациенты в постпубертатном периоде зачастую имеют достаточно развитые молочные железы и распределение жировой ткани по женскому типу, что связано с ароматизацией синтезирующихся в яичках андрогенов в эстрогены.
Предстательная железа
Одним из наиболее андроген-чувствительных органов является простата, имеющая большое число рецепторов к андрогенам. Предполагается, что полиморфизм гена AR с его способностью моделировать эффекты андрогенов влияет на долю злокачественных клеток в предстательной железе (ПЖ) [9]. Согласно метаанализу множества исследований [10], отношение шансов возникновения рака ПЖ при снижении частоты тринуклеотидных повторов составляет 1,19.
Возможная связь между ДГПЖ и CAG-полимор физмом гена AR имеет также другой аспект: ДГПЖ представляет собой разрастание ткани переходной зоны и периуретральной области ПЖ (эпителиальная и фибромышечная гиперплазия). В 2 крупных исследованиях сравнивали сопоставимые когорты здоровых добровольцев и пациентов с ДГПЖ. Отношение шансов увеличения размеров простаты до необходимого хирургического вмешательства в группе с частотой CAG-повторов <20 и >24 составляло 1,92. Более того, при снижении длины CAG-повторов увеличивалась частота средних и тяжелых обструктивных симптомов нижних мочевых путей. Также была найдена обратная корреляция между частотой аденом и размером простаты, с одной стороны, и длиной CAG-повторов — с другой [11, 12]. Помимо этого, была выявлена связь полиморфизма гена AR с сексуальной активностью мужчин с ДГПЖ [13].
В 2 крупных исследованиях сравнивали сопоставимые когорты здоровых добровольцев и пациентов с ДГПЖ. Отношение шансов увеличения размеров простаты до необходимого хирургического вмешательства в группе с частотой CAG-повторов <20 и >24 составляло 1,92. Более того, при снижении длины CAG-повторов увеличивалась частота средних и тяжелых обструктивных симптомов нижних мочевых путей. Также была найдена обратная корреляция между частотой аденом и размером простаты, с одной стороны, и длиной CAG-повторов — с другой [11, 12]. Помимо этого, была выявлена связь полиморфизма гена AR с сексуальной активностью мужчин с ДГПЖ [13].
Фертильность
Установлена связь числа CAG-повторов в гене AR с олигозооспермией [14, 15]. Хорошо известно, что для осуществления сперматогенеза необходима стимуляция клеток Сертоли фолликулостимулирующим гормоном (ФСГ). Так, интратестикулярная активность андрогенов опосредуется важным кофактором, положительно влияющим на функцию клеток Сертоли.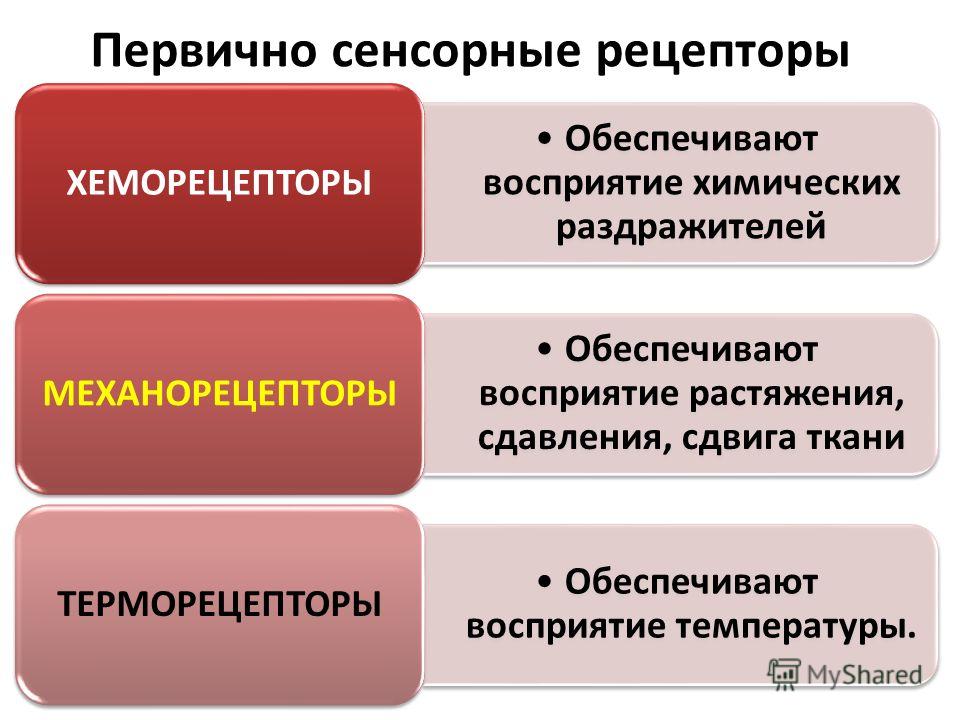 Установлена отрицательная связь числа CAG-повторов с эффективностью сперматогенеза [16]. Наиболее тяжелые формы нарушений сперматогенеза наблюдаются у пациентов со спинобульбарной атрофией Кеннеди — редким наследственным заболеванием, характеризующимся яркой неврологической симптоматикой в сочетании с гипогонадизмом и бесплодием. При этом число тринуклеотидных повторов в гене AR достигает 36 и более [17].
Установлена отрицательная связь числа CAG-повторов с эффективностью сперматогенеза [16]. Наиболее тяжелые формы нарушений сперматогенеза наблюдаются у пациентов со спинобульбарной атрофией Кеннеди — редким наследственным заболеванием, характеризующимся яркой неврологической симптоматикой в сочетании с гипогонадизмом и бесплодием. При этом число тринуклеотидных повторов в гене AR достигает 36 и более [17].
Клинические проявления гипогонадизма и психосоциальные аспекты личности
Анализируя клинические проявления андрогенного дефицита, следует подчеркнуть их тесную связь с полиморфизмом гена AR. Так, среди пациентов с нормальными концентрациями Т у мужчин с большим числом CAG-повторов увеличен риск развития симптомов андрогенного дефицита [18, 19]. Регрессионный анализ данных международного опросника Aging Males’ Symptomsвыявил нарастание симптомов дефицита Т по двум шкалам (психологической и соматической) по мере увеличения длины CAG—повторов в гене AR, тогда как симптомы сексуальной шкалы были связаны непосредственно с уровнем эндогенного Т [18]. Азиатское исследование с участием 702 мужчин, не выявив связи между уровнем Т и распределением в популяции полиморфизма гена AR, показало, что при уровне Т выше 340 нг/дл пациенты с числом CAG-повторов более 25 имели достоверно более высокий риск развития симптомов андропаузы, чем лица со значением тринуклеотидных повторов менее 22 [19]. Иными словами, мужчины с нормотестостеронемией, имеющие большую длину CAG—повторов в гене AR, подвержены более высокому риску возникновения симптомов андрогенного дефицита.
Азиатское исследование с участием 702 мужчин, не выявив связи между уровнем Т и распределением в популяции полиморфизма гена AR, показало, что при уровне Т выше 340 нг/дл пациенты с числом CAG-повторов более 25 имели достоверно более высокий риск развития симптомов андропаузы, чем лица со значением тринуклеотидных повторов менее 22 [19]. Иными словами, мужчины с нормотестостеронемией, имеющие большую длину CAG—повторов в гене AR, подвержены более высокому риску возникновения симптомов андрогенного дефицита.
Кроме того, установлена связь между длиной CAG—повторов в гене AR с неспецифическими проявлениями андрогенного дефицита, такими как тревожность и депрессия [20, 21]. Регрессионный анализ выявил положительную связь числа CAG-повторов с выраженностью депрессии у мужчин старше 50 лет. Это демонстрирует нарастание симптомов депрессии синхронно с увеличением длины тринуклеотидных повторов по мере ослабления чувствительности рецепторов к андрогенам независимо от уровня эндогенного Т [21]. Показано также, что генетически детерминированное удлинение CAG—повторов в гене AR является независимым фактором риска высокой тревожности, панических атак и фобических расстройств [20].
Показано также, что генетически детерминированное удлинение CAG—повторов в гене AR является независимым фактором риска высокой тревожности, панических атак и фобических расстройств [20].
Удивительными кажутся данные о связи полиморфизма гена AR с интеллектуальной одаренностью мальчиков [22]. Так, установлено, что высокий уровень интеллекта (IQ более 130) у мальчиков ассоциирован с малой длиной CAG-повторов в гене AR, даже при допубертатно низких уровнях Т. Другими словами, высокая чувствительность к андрогенам является фактором, предрасполагающим к интеллектуальной одаренности вне зависимости от уровня самого Т.
Представляют интерес данные о полиморфизме гена AR у мужчин с синдромом Клайнфельтера, имеющих дополнительную Х-хромосому, на которой и расположен обсуждаемый ген [23]. Длина тринуклео тидных повторов имела положительную связь с ростом, но в то же время была отрицательно связана с минеральной плотностью костной ткани и отношением размаха рук к росту, что отражает негативное влияние андрогенов на эпифизарные зоны роста кости. Кроме того, удлинение CAG—повторов способствовало развитию гинекомастии и уменьшению размеров тестикул, тогда как укорочение длины триплета ассоциировалось со стабильными партнерскими отношениями и профессиями, требующими более высокого уровня образования.
Кроме того, удлинение CAG—повторов способствовало развитию гинекомастии и уменьшению размеров тестикул, тогда как укорочение длины триплета ассоциировалось со стабильными партнерскими отношениями и профессиями, требующими более высокого уровня образования.
Поразительно, что чувствительность к андрогенам может обусловливать и такое явление, как транссексуализм. Установлено, что удлинение цепи CAG—повторов в гене AR является одной из трех изученных генетических причин мужского транссексуализма [24].
Метаболический статус
Выявлена положительная независимая корреляция числа CAG—повторов в гене AR с содержанием жировой ткани тела, уровнем лептина и инсулина. Малое число CAG-повторов было независимо ассоциировано с протективными параметрами (низкая жировая масса, низкий уровень инсулина плазмы), но одновременно и с нежелательными факторами (низкий уровень липопротеинов высокой плотности) [25]. Сходные данные в отношении дислипидемии были получены и в исследовании больных сахарным диабетом 2-го типа [26].
Сходные данные в отношении дислипидемии были получены и в исследовании больных сахарным диабетом 2-го типа [26].
Еще одним интереснейшим аспектом влияния полиморфизма гена AR является эффективность заместительной терапии тестостероном (ЗТТ) у пациентов с различными формами гипогонадизма. Крупное исследование TIMES 2 показало, что высокая чувствительность к андрогенам, моделируемая малым числом CAG-повторов в гене AR, независимо положительно связана с более значимой динамикой уровня инсулина натощак, триглицеридов, диастолического артериального давления и индекса инсулинорезистентности НОМА на фоне ЗТТ. Однако связи полиморфизма гена AR с уровнем гликированного гемоглобина, другими показателями липидограммы, ожирением или распределением жировой ткани не выявлено [10]. Негативное влияние удли нения CAG—повторов было обнаружено и в другом исследовании [27], показавшем более высокие цифры артериального давления и худшие значения липидограммы на фоне ЗТТ в сочетании с более низкими значениями гематокрита, находящегося под контролем андрогенов. Еще одним параметром, реагирующим на ЗТТ, является размер ПЖ [28]. Выявлена негативная связь между длиной тринуклеотидных повторов и изменением объема ПЖ на фоне терапии Т [10]. Причем 20 триплетов гена AR и менее повышают риск увеличения размеров ПЖ и темпы роста уровня простат-специфического антигена (ПСА) [27].
Еще одним параметром, реагирующим на ЗТТ, является размер ПЖ [28]. Выявлена негативная связь между длиной тринуклеотидных повторов и изменением объема ПЖ на фоне терапии Т [10]. Причем 20 триплетов гена AR и менее повышают риск увеличения размеров ПЖ и темпы роста уровня простат-специфического антигена (ПСА) [27].
Терапия Т у пациентов с синдромом Клайнфельтера при коротких триплетных повторах вызывает более глубокое подавление уровня ЛГ, выраженный рост концентрации ПСА и повышает уровень гемоглобина [23].
Эндотелий
Одной из важнейших точек приложения внегонадного действия Т является эндотелий, причем данное взаимодействие моделируется полиморфизмом гена AR. Этот вопрос наименее изучен. Ранее у здоровых добровольцев была обнаружена связь низкого числа CAG-повторов в гене AR с нарушениями артериальной вазореактивности вне зависимости от уровня общего и свободного T [29]. Некоторыми исследованиями [30, 31] доказано ухудшение функции эндотелия у больных сахарным диабетом 2-го типа. В то же время существуют данные о негативном влиянии дефицита Т на сердечно-сосудистую систему у мужчин [32, 33]. При исследовании влияния полиморфизма гена AR на формирование нарушений функции эндотелия у мужчин с сахарным диабетом 2-го типа [34] было установлено, что большее количество CAG-повторов в гене рецептора AR через ослабление чувствительности к андрогенам обусловливает снижение выраженности эндотелий-зависимой вазодилатации в ходе пробы с реактивной гиперемией одновременно с усилением продукции таких маркеров дисфункции эндотелия, как P-селектин и резистин. Таким образом, увеличение числа CAG-повто ров в гене AR ухудшает как сосудодвигательную, так и секреторную функцию эндотелия и может рассматриваться как предиктор развития и прогрессирования сердечно-сосудистых поражений у мужчин с сахарным диабетом.
В то же время существуют данные о негативном влиянии дефицита Т на сердечно-сосудистую систему у мужчин [32, 33]. При исследовании влияния полиморфизма гена AR на формирование нарушений функции эндотелия у мужчин с сахарным диабетом 2-го типа [34] было установлено, что большее количество CAG-повторов в гене рецептора AR через ослабление чувствительности к андрогенам обусловливает снижение выраженности эндотелий-зависимой вазодилатации в ходе пробы с реактивной гиперемией одновременно с усилением продукции таких маркеров дисфункции эндотелия, как P-селектин и резистин. Таким образом, увеличение числа CAG-повто ров в гене AR ухудшает как сосудодвигательную, так и секреторную функцию эндотелия и может рассматриваться как предиктор развития и прогрессирования сердечно-сосудистых поражений у мужчин с сахарным диабетом.
Заключение
Нарастающий интерес исследователей к изучению биохимических и молекулярных механизмов действия Т диктует необходимость расшифровки генетических аспектов функционирования AR. Изучение чувствительности к андрогенам, определяемой длиной тринуклеотидного повтора CAG в гене AR, не только объясняет феномен различного ответа на ЗТТ у пациентов с одинаковым уровнем эндогенного Т, но и необходимо для понимания половой дифференцировки, психологического статуса, сексуальности и репродуктивного потенциала, а также рисков развития рака ПЖ и ДГПЖ, остеопороза, нарушений углеводного, липидного обменов и даже сердечно-сосудистых заболеваний у мужчин. Наиболее важной в клинической практике является возможность прогнозирования ответа пациента на ЗТТ. У лиц с малым числом CAG-повторов в гене AR из-за высокой чувствительности к андрогенам могут быть использованы более низкие дозы препаратов Т, тогда как неэффективность стартовой дозы ТЗТ у мужчин с большим числом тринуклеотидных повторов, свидетельствует о необходимости повышения дозы вводимого Т. На сегодняшний день определение CAG-полиморфизма гена AR не рекомендовано для рутинной практики, однако в ближайшем будущем оно может быть использовано, в частности для подбора индивидуальной терапии андрогенного дефицита.
Изучение чувствительности к андрогенам, определяемой длиной тринуклеотидного повтора CAG в гене AR, не только объясняет феномен различного ответа на ЗТТ у пациентов с одинаковым уровнем эндогенного Т, но и необходимо для понимания половой дифференцировки, психологического статуса, сексуальности и репродуктивного потенциала, а также рисков развития рака ПЖ и ДГПЖ, остеопороза, нарушений углеводного, липидного обменов и даже сердечно-сосудистых заболеваний у мужчин. Наиболее важной в клинической практике является возможность прогнозирования ответа пациента на ЗТТ. У лиц с малым числом CAG-повторов в гене AR из-за высокой чувствительности к андрогенам могут быть использованы более низкие дозы препаратов Т, тогда как неэффективность стартовой дозы ТЗТ у мужчин с большим числом тринуклеотидных повторов, свидетельствует о необходимости повышения дозы вводимого Т. На сегодняшний день определение CAG-полиморфизма гена AR не рекомендовано для рутинной практики, однако в ближайшем будущем оно может быть использовано, в частности для подбора индивидуальной терапии андрогенного дефицита.
Дополнительная информация
Источник финансирования. Работа выполнена при поддержке Российского научного фонда (грант РНФ №14-25-00052).
Конфликт интересов. Авторы статьи заявляют об отсутствии финансовой заинтересованности и других потенциальных конфликтов интересов, связанных с публикацией настоящей статьи.
Участие авторов:
Сбор и анализ материала, написание текста — Хрипун И.А.; написание и редактирование текста — Воробьев С.В. Оба автора внесли существенный вклад в проведение поисково-аналитической работы и подготовки статьи, прочли и одобрили финальную версию текста перед публикацией.
1. Omim.Org [Internet]. Androgen Receptor; Ar [Cited 2018 Sep 26]. Available From: https://www.omim.org/entry/313700
2. Zitzmann M. Hypogonadism: biochemical diagnosis, treatment, and the myth of uniform thresholds. J Mens Health 2013;10(3):73-75. doi: https://doi.org/10.1089/jomh.2009.00030
Zitzmann M. Hypogonadism: biochemical diagnosis, treatment, and the myth of uniform thresholds. J Mens Health 2013;10(3):73-75. doi: https://doi.org/10.1089/jomh.2009.00030
3. Limer KL, Pye SR, Thomson W, et al. Genetic variation in sex hormone genes influences heel ultrasound parameterso in middle-aged and elderly men: results from the European male aging study (EMAS). J Bone Miner Res. 2009;24(2):314-323. doi: https://doi.org/10.1359/jbmr.080912
4. Zitzmann M. Mechanisms of disease: pharmacogenetics of testosterone therapy in hypogonadal men. Nat Clin Pract Urol. 2007;4(3):161-166. doi: https://doi.org/10.1038/ncpuro0706
5. Guadalupe-Grau A, Rodriguez-Gonzalez FG, Ponce-Gonza lez JG, et al. Bone mass and the CAG and GGN androgen receptor polymorphisms in young men. Plos One. 2010;5(7):E11529. doi: https://doi.org/10.1371/journal.pone.0011529
6. Rodriguez-Garcia L, Ponce-Gonzalez JG, Gonzalez-Henriquez JJ, et al. Androgen receptor CAG and GGN repeat polymorphisms and bone mass in boys and girls. Nutr Hosp. 2015;32(6):2633-2639. doi: https://doi.org/10.3305/nh.2015.32.6.9767
7. Stiger F, Brandstrom H, Gillberg P, et al. Association between repeat length of Exon 1 CAG microsatellite in the androgen receptor and bone density in men is modulated by sex hormone levels. Calcif Tissue Int. 2008;82(6):427-435. doi: https://doi.org/10.1007/s00223-008-9128-3
8. Canale D, Caglieresi C, Moschini C, et al. Androgen receptor polymorphism (CAG repeats) and androgenicity. Clin Endocrinol (Oxf.). 2005;63(3):356-361. doi: https://doi.org/10.1111/j.1365-2265.2005.02354.x
9. Grassetti D, Giannandrea F, Paoli D, et al. Androgen receptor polymorphisms and testicular cancer risk. Andrology. 2015;3(1):27-33. doi: https://doi.org/10.1111/j.2047-2927.2014.00252.x
10. Stanworth RD, Akhtar S, Channer KS, Jones TH. The role of androgen receptor CAG repeat polymorphism and other factors which affect the clinical response to testosterone replacement in metabolic syndrome and type 2 diabetes: times 2 Sub-Study. Eur J Endocrinol. 2014;170(2):193-200. doi: https://doi.org/10.1530/eje-13-0703
11. Giovannucci E, Stampfer MJ, Chan A, et al. CAG repeat within the androgen receptor gene and incidence of surgery for benign prostatic hyperplasia in U.S. Physicians. Prostate. 1999;39(2): 130-134. doi: https://doi.org/10.1002/(sici)1097-0045(19990501)39:2<130:: aid-pros8>3.0.co;2-#
12. Giovannucci E, Platz EA, Stampfer MJ, et al. The CAG repeat within the androgen receptor gene and benign prostatic hyperplasia. Urology. 1999;53(1):121-125. doi: https://doi.org/10.1016/s0090-4295(98)00468-3
13. Коган М.И., Киреев А.Ю. Различия уровней тестостерона и полиморфизма андрогенового рецептора у пациентов с симптомной доброкачественной гиперплазией простаты в зависимости от сексуальной активности. // Андрология и генитальная хирургия. — 2009. — Т. 10. — №2. — C. 118. [Kogan MI, Kireev AYu. Razlichiya urovney testosterona i polimorfizma androgenovogo retseptora u patsientov s simptomnoy dobrokachestvennoy giperplaziey prostaty v zavisimosti ot seksual’noy aktivnosti. Andrology and Genital Surgery Journal. 2009;10(2):118. (In Russ.)].
14. Giagulli VA, Carbone MD, De Pergola G, et al. Could androgen receptor gene CAG tract polymorphism affect spermatogenesis in men with idiopathic infertility? J Assist Reprod Genet. 2014. doi: https://doi.org/10.1007/s10815-014-0221-4
15. Delli Muti N, Tirabassi G, Buldreghini E, et al. Synergistic effect of androgen receptor (CAG repeat length) and endothelial nitric oxide synthase (Glu298asp Variant) gene polymorphisms on seminal parameters in men with idiopathic oligoasthenozoospermia. Endocrine. 2014;47(1):322-324. doi: https://doi.org/10.1007/s12020-014-0193-2
16. Xiao F, Lan A, Lin Z, et al. Impact of CAG repeat length in the androgen receptor gene on male infertility — a metaanalysis. Reprod Biomed Online. 2016;33(1):39-49. doi: https://doi.org/10.1016/j.rbmo.2016.03.012
17. Finsterer J, Soraru G. Onset manifestations of spinal and bulbar muscular atrophy (Kennedy’s disease). J Mol Neurosci. 2016; 58(3):321-329. doi: https://doi.org/10.1007/s12031-015-0663-x
18. Schneider G, Nienhaus K, Gromoll J, et al. Aging males’ symptoms in relation to the genetically determined androgen receptor CAG polymorphism, sex hormone levels and sample membership. Psychoneuroendocrinology. 2010;35(4):578-587. doi: https://doi.org/10.1016/j.psyneuen.2009.09.008
19. Liu CC, Lee YC, Wang CJ, et al. The impact of androgen receptor CAG repeat polymorphism on andropausal symptoms in different serum testosterone levels. J Sex Med. 2012;9(9):2429-2437. doi: https://doi.org/10.1111/j.1743-6109.2012.02672.x
20. Schneider G, Nienhaus K, Gromoll J, et al. Sex gormone levels, genetic androgen receptor polymorphism, and anxiety in >/=50-year-old males. J Sex Med. 2011;8(12):3452-3464. doi: https://doi.org/10.1111/j.1743-6109.2011.02443.x
21. Schneider G, Nienhaus K, Gromoll J, et al. Depressive symptoms in men aged 50 years and older and their relationship to genetic androgen receptor polymorphism and sex hormone levels in three different samples. Am J Geriatr Psychiatry. 2011;19(3):274-283. doi://https://doi.org/10.1097/jgp.0b013e381e70c22
22. Celec P, Tretinarova D, Minarik G, et al. Genetic polymorphisms related to testosterone metabolism in intellectually gifted boys. Plos One. 2013;8(1):E54751. doi: https://doi.org/10.1371/journal.pone.0054751
23. Zitzmann M, Depenbusch M, Gromoll J, Nieschlag E. X-Chromosome inactivation patterns and androgen receptor functionality influence phenotype and social characteristics as well as pharmacogenetics of testosterone therapy in Klinefelter patients. J Clin Endocrinol Metab. 2004;89(12):6208-6217. doi: https://doi.org/10.1210/jc.2004-1424
24. Hare L, Bernard P, Sanchez FJ, et al. Androgen receptor repeat length polymorphism associated with male-to-female transsexualism. Biol Psychiatry. 2009;65(1):93-96. doi: https://doi.org/10.1016/j.biopsych.2008.08.033
25. Zitzmann M, Gromoll J, Von Eckardstein A, Nieschlag E. The CAG repeat polymorphism in the androgen receptor gene modulates body fat mass and serum concentrations of leptin and insulin in men. Diabetologia. 2003;46(1):31-39. doi: https://doi.org/10.1007/s00125-002-0980-9
26. Stanworth RD, Kapoor D, Channer KS, Jones TH. Dyslipidaemia is associated with testosterone, oestradiol and androgen receptor CAG repeat polymorphism in men with type 2 diabetes. Clin Endocrinol (Oxf). 2011;74(5):624-630. doi: https://doi.org/10.1111/j.1365-2265.2011.03969.x
27. Zitzmann M, Nieschlag E. Androgen receptor gene CAG repeat length and body mass index modulate the safety of long-term intramuscular testosterone undecanoate therapy in hypogonadal men. J Clin Endocrinol Metab. 2007;92(10):3844-3853. doi: https://doi.org/10.1210/jc.2007-0620
28. Zitzmann M, Depenbusch M, Gromoll J, Nieschlag E. Prostate volume and growth in testosterone-substituted hypogonadal men are dependent on the CAG repeat polymorphism of the androgen receptor gene: a longitudinal pharmacogenetic study. J Clin Endocrinol Metab. 2003;88(5):2049-2054. doi: https://doi.org/10.1210/jc.2002-021947
29. Zitzmann M, Brune M, Kornmann B, et al. The CAG repeat polymorphism in the AR gene affects high density lipoprotein cholesterol and arterial vasoreactivity. J Clin Endocrinol Metab. 2001;86(10):4867-4873. doi: https://doi.org/10.1210/jcem.86.10.7889
30. Хрипун И.А., Воробьев С.В., Моргунов М.Н., Коган М.И. Функция эндотелия у мужчин с сахарным диабетом 2-го типа, не имеющих клинических признаков сердечно-сосудистых заболеваний. // Сахарный диабет. — 2016. — Т. 19. — №5. — С. 383-387. [Khripun IA, Vorobyev SV, Morgunov MN, Kogan MI. Endothelial function in men with type 2 diabetes without clinical signs of cardiovascular disease. Diabetes Mellitus. 2016;19(5):383-387. (In Russ.)]. doi: https://doi.org/10.14341/dm8017
31. Van Sloten TT, Henry RM, Dekker JM, et al. Endothelial dysfunction plays a key role in increasing cardiovascular risk in type 2 diabetes: the Hoorn Study. Hypertension. 2014;64(6):1299-1305. doi: https://doi.org/10.1161/hypertensionaha.114.04221
32. Хрипун И.А., Воробьев С.В., Пузырева В.П., и др. Дисфункция эндотелия как следствие андрогенного дефицита у мужчин с сахарным диабетом 2-го типа. // Современные проблемы науки и образования. — 2015. — № 6. [Khripun IA, Vorobyev SV, Puzyreva VP, et al. Dysfunction of endothelium, as a result of androgen deficiency in men with type 2 diabetes. Modern Problems Of Science And Education. 2015;(6). (In Russ.)].
33. Kelly DM, Jones TH. Testosterone and cardiovascular risk in men. Front Horm Res. 2014;43:1-20. doi: https://doi.org/10.1159/000360553
34. Хрипун И.А., Воробьев С.В., Коган М.И. Взаимосвязь полиморфизма гена рецептора андрогенов и функции эндотелия у мужчин с сахарным диабетом 2-го типа. // Сахарный диабет. — 2015. — Т. 18. — №4. — С. 35-40. [Khripun IA, Vorobyev SV, Kogan MI. Association of the polymorphism in the androgen receptor gene and endothelial function in men with type 2 diabetes. Diabetes Mellitus. 2015;18(4):35-40. (In Russ.)]. doi: https://doi.org/10.14341/dm7622
РЕЦЕПТОРЫ | Психологическая энциклопедия 1vc0
Оглавление раздела ощущения:
Рецептор — орган, специально приспособленный для рецепции раздражений, легче, чем прочие органы или нервные волокна, поддаётся раздражению; он отличается особенно низкими порогами раздражения, т. е. его чувствительность, обратно пропорциональная порогу, особенно высока. В этом первая особенность рецептора как специализированного аппарата: обладая особенно большой чувствительностью, он специально приспособлен для рецепции раздражений.
При этом рецепторы приспособлены для рецепции не любых раздражителей. Каждый рецептор специализируется применительно к определённому раздражителю. Формируясь в процессе развития под воздействием определённого вида раздражителей, каждый рецептор является поэтому приспособленным для рецепции именно этого вида раздражителей. Так, образуются тангорецепторы, приспособленные к рецепции прикосновения, густорецепторы — для рецепции вкусовых раздражений, стиборецепторы — для обонятельных, приспособленные для рецепции звука и света фоно- и фоторецепторы — слух, зрение. Каждый из этих рецепторов установлен на особый вид раздражителя, который является адекватным для него.
Таким образом, специальная приспособленность к рецепции раздражений, выражающаяся в особо высокой чувствительности, — во-первых, и приспособленность к рецепции специальных раздражителей, т. е. специализация рецепторов по виду раздражителей, — во-вторых, составляют основные черты, характеризующие рецепторный аппарат.
В парадоксальной форме специализация органов чувств, или рецепторов, выражается в том, что и неадекватный раздражитель, воздействуя на определённый рецептор, может вызвать специфические для него ощущения. Так, сетчатка даёт световые ощущения при воздействии на неё как светом, так и электрическим током или давлением («искры из глаз сыплются» при ударе). С другой стороны, механический раздражитель может дать ощущение давления, звука или света в зависимости от того, воздействует ли он на осязание, слух или зрение. Получающееся таким образом нарушение адекватного соотношения между ощущением и раздражителем представляет собой всё же лишь исключение. Тем не менее, основываясь на этих фактах и опираясь на специализацию «органов чувств», И. Мюллер выдвинул свой принцип специфической энергии органов чувств. Основу его составляет бесспорное положение, заключающееся в том, что все специфицированные ощущения находятся в определённом соотношении с гистологически специфицированными органами, их обусловливающими. Это правильное положение, подтверждаемое обширными психофизиологическими данными, завоевало принципу специфической энергии органов чувств универсальное признание у физиологов.
На этой основе И. Мюллер выдвигает другую идею, согласно которой ощущение зависит не от природы раздражителя, а от органа или нерва, в котором происходит процесс раздражения, и является выражением его специфической энергии. Посредством зрения, например, по И. Мюллеру, познаётся несуществующий во внешнем мире свет, поскольку глаз наш доставляет впечатление света и тогда, когда на него действует электрический или механический раздражитель, т. е. в отсутствие физического света. Ощущение света признаётся выражением специфической энергии сетчатки: оно — лишь субъективное состояние сознания. Включение физиологических процессов в соответствующем аппарате в число объективных, опосредующих условий ощущения превращается, таким образом, в средство отрыва ощущения от его внешней причины и признания субъективности ощущения. Из связи субъекта с объектом ощущение превращается во включённую между субъектом и объектом завесу.
На этой основе и сложилась та концепция, которую сформулировал Гельмгольц. Плеханов, опираясь на неё, выдвинул свою теорию иероглифов, которую Ленин подверг острой критике: «Бесспорно, что изображение никогда не может всецело сравниться с моделью, но одно дело изображение, другое дело символ, условный знак. Изображение необходимо и неизбежно предполагает объективную реальность того, что «отображается». «Условный знак», символ, иероглиф суть понятия, вносящие совершенно ненужный элемент агностицизма». [В. И. Ленин, Соч., т. XIII, 1931. стр. 193.]
Стоит подойти к интерпретации того позитивного фактического положения, которое лежит в основе субъективно-идеалистической надстройки, возведённой над нею И. Мюллером, чтобы те же факты предстали в совсем ином освещении. В процессе биологической эволюции сами органы чувств формировались в реальных взаимоотношениях организма со средой, под воздействием внешнего мира. Специализация самых органов чувств совершалась под воздействием внешних раздражителей; воздействие внешнего мира формирует сами рецепторы. Рецепторы являются как бы анатомически закреплёнными в строении нервной системы отпечатками эффектов процессов раздражения. Нужно, собственно, говорить не столько о специфической энергии органов чувств, сколько об органах чувств специфической энергии. «Специфическая энергия» органов чувств или нервов, взятая в генетическом плане, выражает, таким образом, пластичность нерва по отношению к специфичности внешнего раздражителя. Источников специфичности нужно первично искать не внутри, а во вне. Она свидетельствует не о субъективности ощущения, а об его объективности. Эта объективность, конечно, не абсолютная. Ощущение и степень его адекватности действительности обусловлены и состоянием рецептора, а также и воспринимающего организма в целом. Существуют и иллюзии и галлюцинации, существуют обманы чувств. Но именно поэтому мы и можем говорить о некоторых показаниях чувств как иллюзиях, галлюцинациях и обманах чувств, что они в этом отношении отличаются от других объективных, адекватных действительности показаниях органов чувств. Критерием для различения одних от других служит действие, практика, контролирующая объективность наших ощущений как субъективного образа объективного мира.
Элементы психофизики
Наличие зависимости ощущений от внешних раздражений заставляет поставить вопрос о характере этой зависимости, т. е. об основных закономерностях, которым она подчиняется. Это центральный вопрос так называемой психофизики. Её основы заложены исследованиями Э. Г. Вебера и Г. Т. Фехнера. Оформление она получила в «Элементах психофизики» (1859) Фехнера, оказавших значительное влияние на дальнейшие исследования. Основной вопрос психофизики — это вопрос о порогах. Различают абсолютные и разностные пороги ощущения или пороги ощущения и пороги различения.
Исследования по психофизике установили прежде всего, что не всякий раздражитель вызывает ощущение. Он может быть так слаб, что не вызовет никакого ощущения. Мы не слышим множества вибраций окружающих нас тел, не видим невооружённым глазом множества постоянно вокруг нас происходящих микроскопических изменений. Нужна известная минимальная интенсивность раздражителя для того, чтобы вызвать ощущение. Эта минимальная интенсивность раздражения называется нижним абсолютным порогом. Нижний порог даёт количественное выражение для чувствительности: чувствительность рецептора выражается величиной, обратно пропорциональной порогу: , где E — чувствительность и J — пороговая величина раздражителя.
Наряду с нижним существует и верхний абсолютный порог, т. е. максимальная интенсивность, возможная для ощущения данного качества. В существовании порогов рельефно выступает диалектическое соотношение между количеством и качеством. Эти пороги для различных видов ощущений различны. В пределах одного и того же вида они могут быть различны у различных людей, у одного и того же человека в разное время, при различных условиях.
За вопросом о том, имеет ли вообще место ощущение определённого вида (зрительное, слуховое и т. д.), неизбежно следует вопрос об условиях различения различных раздражителей. Оказалось, что наряду с абсолютными существуют разностные пороги различения. Э. Вебер установил, что требуется определённое соотношение между интенсивностями двух раздражителей для того, чтобы они дали различные ощущения. Это соотношение выражено в законе, установленном Э. Вебером: отношение добавочного раздражителя к основному должно быть величиной постоянной:
ΔJ÷J=K,
где J обозначает раздражение, ΔJ — его прирост, K — постоянная величина, зависящая от рецептора.
Так, в ощущении давления величина прибавки, необходимой для получения едва заметной разницы, должна всегда равняться приблизительно 1/30 исходного веса, т. е. для получения едва заметной разницы в ощущении давления к 100 г нужно добавить 3,4 г, к 200 — 6,8 г, к 300 — 10,2 г и т. д. Для силы звука эта константа равна 1/10, для силы света — 1/100 и т. д.
Дальнейшие исследования показали, что закон Э. Вебера действителен лишь для раздражителей средней величины: при приближении к абсолютным порогам величина прибавки перестаёт быть постоянной величиной. Наряду с этим ограничением закон Э. Вебера допускает, как оказалось; и расширение. Он применим не только к едва заметным, но и ко всяким различиям ощущений. Различия между парами ощущений кажутся нам равными, если равны геометрические соотношения соответствующих раздражителей. Так, увеличение силы освещения от 25 до 50 свечей даёт субъективно такой же эффект, как увеличение от 50 до 100.
Исходя из закона Э. Вебера, Г. Фехнер сделал допущение, что едва заметные разницы в ощущениях можно рассматривать как равные, поскольку все они — величины бесконечно малые, и принять их как единицу меры, при помощи которой можно численно выразить интенсивность ощущений как сумму (или интеграл) едва заметных (бесконечно малых) увеличений, считая от порога абсолютной чувствительности. В результате он получил два ряда переменных величин — величины раздражителей и соответствующие им величины ощущений. Ощущения растут в арифметической прогрессии, когда раздражители растут в геометрической прогрессии. Отношение этих двух переменных величин можно выразить в логарифмической формуле:
E = K · log J + C,
где K и C суть некоторые константы. Эта формула, определяющая зависимость интенсивности ощущений (в единицах едва заметных перемен) от интенсивности соответствующих раздражителей, и представляет собой так называемый психофизический закон Вебера-Фехнера.
Допущенная при этом Фехнером возможность суммирования бесконечных, а не только конечных разностей ощущений, большинством исследований считается произвольной. Помимо того нужно отметить, что ряд явлений, вскрытых новейшими исследованиями чувствительности, не укладывается в рамки закона Вебера-Фехнера. Особенно значительное противоречие с законом Вебера-Фехнера обнаруживают явления протопатической чувствительности, поскольку ощущения в области протопатической чувствительности не обнаруживают постепенного нарастания по мере усиления раздражения, а по достижении известного порога сразу же появляются в максимальной степени. Они приближаются по своему характеру к типу реакций по принципу «всё или ничего». Не согласуются, по-видимому, с законом Вебера-Фехнера и некоторые данные современной электрофизиологии органов чувств.
Дальнейшие исследования Г. Гельмгольца, подтверждённые П. П. Лазаревым, заменили первоначальную формулировку закона Вебера-Фехнера более сложной формулой, выражающей очень общий принцип, управляющий всеми явлениями раздражения. Однако и попытка Лазарева выразить переход раздражения в ощущение в математических уравнениях не охватывает всего многообразия процессов чувствительности и перехода раздражения в ощущение.
Для определения порогов была разработана целая система методов психофизического исследования. Из них основные: 1) метод едва заметных различий: прогрессивно изменяют — увеличивают или уменьшают — раздражитель, пока испытуемый не начнёт или не перестанет замечать разницу; 2) метод истинных и ложных случаев: испытуемому предъявляют для сравнения два любых раздражителя и предлагают определить, какой из них больше; 3) метод средних ошибок, или метод константности: испытуемый должен подобрать к данному ему раздражителю равные. При всех этих методах пороги определяются как статистические средние.
Значение измерения порогов заключается в том, что они являются главной основой для точного, количественно выраженного определения дифференциальных различий в сенсорной области — от вида к виду, от индивида к индивиду и у одного и того же индивида в различных условиях — в зависимости от утомления, упражнения, образования и т. д. Тем самым они дают возможность исследовать и значение всех влияющих на сенсорную область высших факторов и косвенно устанавливают их уровень. Они поэтому доставляют данные для ряда выводов, имеющих и практическую значимость.
Пороги и, значит, чувствительность органов никак не приходится представлять как некие раз и навсегда фиксированные неизменные лимиты. Целый ряд исследований советских авторов показал их чрезвычайную изменчивость. Так, А. И. Богословский [А. И. Богословский, Опыт выработки сенсорных условных рефлексов у человека, «Физиологический журнал СССР», 1938, стр. 1017.], К. X. Кекчеев [К. X. Кекчеев, «Бюллетень эксперим. биологии и медицины», 1935, вып. 5—6. стр. 358.] и А. О. Долин [А. О. Долин, «Архив биологических наук», 1936, вып. 1—2.] показали, что чувствительность органов чувств может изменяться посредством образования интерсенсорных условных рефлексов (которые подчиняются вообще тем же законам, что и обычные двигательные и секреторные условные рефлексы). Очень отчётливо явление сенсибилизации было в последнее время выявлено рядом исследовании в отношении слуховой чувствительности. Так, А. И. Бронштейн [А. И. Бронштейн, О синтезирующем влиянии звукового раздражения на орган слуха // Бюллетень экспериментальной биологии и медицины. 1936. Т. I. Вып. 4: Сообщения 1 и 2; Т. II. Вып. 5: Сообщение 3.] констатировал понижение порогов слышимости под влиянием повторяющихся звуковых раздражений. Б. М. Теплов обнаружил резкое понижение порогов различения высоты в результате очень непродолжительных упражнений (см. раздел «Слуховые ощущения» этой главы). В. И. Кауфман — в противовес тенденции К. Сишора (Seashore), Г. М. Уиппла и др. рассматривать индивидуальные различия порогов звуковысотной чувствительности исключительно как неизменяющиеся природные особенности организма — экспериментально показал, во-первых, зависимость порогов (так же как самого типа) восприятия высотных разностей от характера музыкальной деятельности испытуемых (инструменталисты, пианисты и т. д.) и, во-вторых, изменяемость этих порогов (и самого типа) восприятия высотных разностей. Кауфман поэтому приходит к тому выводу, что способность различения высоты звука зависит от конкретных особенностей деятельности данной личности и может в известной мере изменяться в зависимости от неё. [В. И. Кауфман, Восприятие малых высотных разностей. Сб. «Исследования по проблеме чувствительности» под ред. В. П. Осипова и Б. Г. Ананьева, т. XIII, 1940.] Н. К. Гусев пришёл к аналогичным результатам о роли практики дегустации в развитии вкусовой чувствительности. [Там же.]
Вообще пороги чувствительности не представляют собой абсолютной, неизменной величины. Экспериментальное исследование показывает, что даже такие явления, которые обусловлены в основном, казалось бы, периферическими факторами, не предопределяются органом самим по себе. Так, например, снижение световой чувствительности периферического зрения в ходе темновой адаптации, вызванное предварительным «засветом» периферии сетчатки, существенно зависит от центральных психологических факторов и может быть (как показывает проводимое в Институте психологии исследование Семёновской) снято при внимании.
Пороги чувствительности существенно сдвигаются в зависимости от отношения человека к той задаче, которую он разрешает, дифференцируя те или иные чувственные данные. Один и тот же физический раздражитель одной и той же интенсивности может оказаться и ниже и выше порога чувствительности и, таким образом, быть или не быть замеченным в зависимости от того, какое значение он приобретает для человека: появляется ли он как безразличный момент окружения для данного индивида или становится имеющим определённое значение показателем существенных условий его деятельности. Поэтому, для того чтобы исследование чувствительности дало сколько-нибудь законченные результаты и привело к практически значимым выводам, оно должно, не замыкаясь в рамках одной лишь физиологии, перейти и в план психологический. Психологическое исследование имеет, таким образом, дело не только с «раздражителем», но и с предметом, и не только с органом, но и с человеком. Этой более конкретной трактовкой ощущения в психологии, связывающей его со всей сложной жизнью личности в её реальных взаимоотношениях с окружающим миром, обусловлено особое значение психологического и психофизиологического, а не только физиологического, исследования для разрешения вопросов, связанных с нуждами практики.
Психофизиологические закономерности
Характеристика ощущений не исчерпывается психофизическими закономерностями. Для чувствительности органа имеет значение и физиологическое его состояние (или происходящие в нём физиологические процессы). Значение физиологических моментов сказывается прежде всего в явлениях адаптации, в приспособлении органа к длительно воздействующему раздражителю; приспособление это выражается в изменении чувствительности — понижении или повышении её. Примером может служить факт быстрой адаптации к одному какому-либо длительно действующему запаху, в то время как другие запахи продолжают чувствоваться так же остро, как и раньше.
Уменьшение интенсивности ощущения при длительном воздействии раздражителя находит себе объяснение в уменьшении частоты нервных импульсов, пробегающих по афферентному нерву, идущему от рецептора в центральную нервную систему. Эдриан (Adrian) установил, что все рецепторы могут быть разбиты на две группы: 1) рецепторы с быстрой адаптацией к раздражителю и быстрым привыканием к нему, что выражается ослаблением ощущения при продолжающемся раздражении органа чувств (тактильные рецепторы), и 2) рецепторы с медленной адаптацией, при деятельности которых ощущение почти не ослабевает (проприорецепторы).
С адаптацией тесно связано и явление контраста, которое сказывается в изменении чувствительности под влиянием предшествующего (или сопутствующего) раздражения. Так, в силу контраста обостряется ощущение кислого после ощущения сладкого, ощущение холодного после горячего и т. д. Следует отметить также свойство рецепторов задерживать ощущения, выражающееся в более или менее длительном последействии раздражений. Так же как ощущение не сразу достигает своего окончательного значения, оно не сразу исчезает после прекращения раздражения, а держится некоторое время и лишь затем постепенно исчезает. Благодаря задержке при быстром следовании раздражений одного за другим происходит слияние отдельных ощущений в единое, слитное целое, как, например, при восприятии мелодий, кинокартины и пр.
Дифференциация и специализация рецепторов не исключает их единства, выражающегося в их взаимодействии. Это взаимодействие рецепторов выражается, во-первых, во влиянии, которое раздражение одного рецептора оказывает на пороги другого. Так, зрительные раздражения влияют на пороги слуховых, а слуховые раздражения — на пороги зрительных (Лазарев, Кравков), точно так же на пороги зрительных ощущений оказывают влияние и обонятельные ощущения (см. дальше).
На взаимодействии рецепторов основан метод сенсибилизации одних органов чувств и, в первую очередь, глаза и уха путём действия на другие органы чувств слабыми или кратковременными, адекватными для них, раздражениями.
Взаимосвязь ощущений проявляется, во-вторых, в так называемой синестезии. Под синестезией разумеют такое слияние качеств различных сфер чувствительности, при котором качества одной модальности перекосятся на другую, разнородную, — например, при цветном слухе качества зрительной сферы — на слуховую. Формой синестезии, относительно часто наблюдающейся, является так называемый цветной слух (audition colorée). У некоторых людей (например у А. Н. Скрябина; в ряде случаев, которые наблюдал А. Бине; у мальчика, которого исследовал А. Ф. Лазурский; у очень музыкального подростка, которого имеет возможность наблюдать автор) явление цветного слуха выражено очень ярко. Отдельные выражения, отражающие синестезии различных видов ощущений, получили права гражданства в литературном языке; так, например, говорят о кричащем цвете, а также о тёплом или холодном колорите и о тёплом звуке (тембре голоса), о бархатистом голосе.
Теоретически природа этого явления не вполне выяснена. Иные авторы склонны объяснять его общностью аффективных моментов, придающих ощущениям различных видов один и тот же общий им эмоционально-выразительный характер.
Взаимодействие рецепторов выражается, наконец, в той взаимосвязи ощущений различных рецепторов, которое постоянно происходит в каждом процессе восприятия любого предмета или явления. Такое взаимодействие осуществляется в совместном участии различных ощущений, например зрительных и осязательных, в познании какого-нибудь предмета или его свойства, как-то — форма, фактура и т. п. (Даже тогда, когда непосредственно в восприятии участвует лишь один рецептор, ощущения, которые он нам доставляет, бывают опосредованы данными другого. Так, при осязательном распознавании формы предмета, когда зрение почему-либо выключено, осязательные ощущения опосредуются зрительными представлениями.) В самом осязании имеет место взаимодействие собственно кожных ощущений прикосновения с мышечными, кинестетическими ощущениями, к которым при ощущении поверхности предмета примешиваются ещё и температурные ощущения. При ощущении терпкого, едкого и т. п. вкуса какой-нибудь пищи к собственно вкусовым ощущениям присоединяются, взаимодействуя с ними, ощущения осязательные и лёгкие болевые. Это взаимодействие осуществляется и в пределах одного вида ощущений. В области зрения, например, расстояние влияет на цвет, ощущения глубины — на форму и т. д. Из всех форм взаимодействия эта последняя, конечно, важнейшая, потому что без неё вообще не существует восприятия действительности.
Любви не будет. Новость про ген супружеской неверности оказалась преувеличена
Начнем с утверждений в прессе. Вот цитата из «Комсомольской правды» — она достаточно характерна:
Если у женщины изменен ген AVPR1A, то партнеру надо быть готовым, простите уж за невольный каламбур, к ее изменам. Рано или поздно, а скорее рано, такая женщина переспит с кем-нибудь еще.
Ген AVPR1A есть у всех. И у мужчин, и у женщин. Ген — это кусочек ДНК, с которого можно снять своего рода слепок в виде молекулы РНК. Молекулы РНК используются как матрицы для синтеза белков. Ген AVPR1A позволяет нашему организму синтезировать один из вазопрессиновых рецепторов. Или, как говорят биологи, ген AVPR1A кодирует вазопрессиновый рецептор. Вазопрессин — это важный гормон, а рецепторы — молекулы, при помощи которых клетки взаимодействуют с гормоном. Вазопрессиновые рецепторы есть у каждого человека.
Ген AVPR1A — как и практически все прочие гены — бывает у людей в нескольких вариантах. Все варианты позволяют клеткам обзавестись вполне работоспособными рецепторами, однако строение и функции этих рецепторов немного отличаются. Можно провести аналогию с бытовой техникой: в каждом доме есть холодильник, но холодильники бывают разных марок, разного объема и с разными дополнительными функциями. Сказать, что какой-то вариант гена (на языке генетиков — «какая-либо из аллелей») лучше или хуже, зачастую столь же сложно, как и ответить на вопрос: «Какой холодильник вы мне посоветуете?» Одним клеткам в одних условиях лучше один вариант, другим и в других обстоятельствах — другой. Нервная клетка в сетчатке глаза и клетка печени занимаются слишком разной работой, но организм не может себе позволить роскошь держать гены, оптимизированные под каждую отдельную задачу.
Как имногие другие рецепторы, AVPR1A вовлечен сразу во множество разных процессов. Если посмотреть, сколько AVPR1A синтезируется в разных тканях и органах (это называется диаграммой экспрессии), то мы увидим пики активности в печени, почках, а также в стенках сосудов. Все это вряд ли имеет даже косвенное отношение к супружеским изменам. Однако AVPR1A также синтезируется в мозге. Перескажем еще раз главное: большинство генов существуют в нескольких вариантах и выполняют сразу несколько функций. Сообщения вида «найден ген Z, отвечающий за X» по этой причине некорректны в принципе: просто один из вариантов гена Z наряду со всем прочим как-то влияет на проявление X. Как правило, гены не появляются только ради одного-единственного эффекта, это слишком уж невыгодно с точки зрения эволюции.
С учетом сказанного выше, можно, не заглядывая в научную статью, предположить, что влияние гена на частоту супружеских измен будет не абсолютным, а скорее вероятностным. Если статью все-таки открыть, то мы узнаем следующее: вклад генетических факторов во внебрачных связях оценивается примерно в 62% для мужчин и в 40% для женщин. При этом ученые проверили два ранее намеченных гена и выяснили, что один из них, кодирующий окситоциновый рецептор OXTR, к изменам отношения не имеет. А вот один определенный вариант AVPR1A встречался чаще у тех, кто изменял своему партнеру. Правда, не настолько часто, чтобы можно было говорить о непреодолимой тяге носительниц этой аллели к внебрачным связям.
Для терминологической точности определим «внебрачные» связи как секс с кем-либо без согласия постоянного партнера или партнерши. Поскольку часть исследований, о которых мы сейчас расскажем, сделана на мышах или иных животных, это уточнение явно не будет лишним (хотя и тут неясно, что подразумевать под согласием).
Контекст
Ген AVPR1A уже не первый раз исследуют в связи с его возможным влиянием на поведение человека. В самой идее о влиянии на наше поведение гормонов, конечно, нет ничего нового : про влияние адреналина слышали практически все, равно как и про важность йода для умственного развития (йод входит в состав гормонов щитовидной железы). Нарушения сна из-за фонаря под окном или после межконтинентального перелета тоже связаны с действием гормонов, на этот раз мелатонина. Падение уровня тестостерона уменьшает сексуальное влечение у мужчин, а выделение окситоцина после родов у родителей (причем обоих, хотя матери подвержены этому намного больше) увеличивает привязанность к ребенку. А раз гормоны действуют не сами по себе, но в связке с рецепторами, то ученым необходимо изучать и рецепторы, и необходимые для производства гормонов гены. Ниже вы видите даже не все, но лишь избранные труды по теме . Каждый мы снабдили ссылкой на оригинал для всех желающих. Итак…
В 2007 году журнал Human Mutation опубликовал исследование австралийских ученых. Они обнаружили, что вариации гена AVPR1A связаны с сексуальным поведением — есть аллели, носители и носительницы которых несколько раньше начинают половую жизнь. А еще раньше, в 2004-м, Twin Research представил статью британской группы, которая пыталась найти аналогичную связь, но не преуспела в этом. Тогда исследователям удалось лишь проанализировать данные о более чем 1600 сестрах-близнецах (в новой статье их более семи тысяч) и сделать вывод о вкладе генетического фактора во внебрачное поведение на уровне около сорока процентов.
Подчеркнем, что этот показатель — склонность к изменам у женщин примерно на сорок процентов может быть обусловлена генами — хорошо согласуется в двух разных научных публикациях. Но сексуальность — это далеко не единственная причина интересоваться геном AVPR1A в контексте поведения, а не регуляции артериального давления или работы печени.
В 2008 году появилась публикация, авторы которой обнаружили корреляцию между наличием определенной аллели AVPR1A и стилем игры в «Диктатора». «Диктатор» — это специально разработанная экономистами и психологами игра для двух участников. Один, «диктатор», получает некоторую сумму на руки, после чего на свое усмотрение делится (или не делится) с напарником. Ученые показали, что среди игроков, склонных забирать все себе, больше людей с определенным вариантом AVPR1A. Статья с этими выводами обнаруживается в журнале Genes, Brain and Behavior.
Тогда же шведско-американская группа показала, что аллели гена AVPR1A могут помогать или затруднять формирование межличностных контактов, причем преимущественно у мужчин. Выводы ученых опубликовал Proceedings of the National Academy of Sciences.
В 2010 году корейские исследователи нашли возможную связь с аутизмом и написали статью в Neuroscience Letters.
В 2011-м американские психиатры представили данные о влиянии гена на формирование наркотической зависимости. Статья с их аргументами представлена в Biological Psychiatry.
В 2012 году канадские медики выявили, что вариации в гене вазопрессинового рецептора AVPR1B — обратите внимание, это другой ген и немного иной рецептор! — связаны с агрессивным поведением у детей. Свои выводы они опубликовали в Psychiatry Research.
В 2013 году польские исследователи нашли некоторые доказательства, которые говорят о возможном влиянии вазопрессиновых рецепторов AVPR1A на развитие депрессивных расстройств. Эти данные представлены в Psychiatric Genetics. А шведские психиатры выяснили, что вариации гена коррелируют с попытками суицида. Разумеется, в статье для журнала Neuropsychopharmacology уточняется, что наличие определенной аллели вовсе не толкает людей на самоубийство. В конце концов, такой ген, если бы и существовал, оказался бы уничтожен естественным отбором!
В 2014-м американские авторы показали, что женщины (но не мужчины) с определенной аллелью острее реагируют на стресс и скорее испытывают гнев в качестве последующей реакции. Статья — в журнале Emotion.
Ген изучался даже на предмет того, может ли он повлиять на способность к пению и музицированию. В 2012 году британские ученые представили в PLoS One статью, где честно сообщили о нулевом результате. То, что до них такую связь все-таки находили, исследователи объяснили многофакторностью процесса формирования хора или музыкального ансамбля, ведь людей туда набирают не только за их способности, но и с учетом психологической совместимости. Возможно, что вариации гена влияют на способность уживаться с окружающими, а это, в свою очередь, повышает шансы быть приглашенным в ансамбль.
Мы также ничего не пишем про исследование роли полиморфизма (еще один термин, обозначающий изменчивость) AVPR1A в агрессивности свиней или в успешности спортивных тренировок у пенсионеров. Достаточно просто сказать, что есть и такие публикации.
Показанные нами исследования подкрепляют сделанный вначале вывод. Генов много, у каждого есть несколько функций, поэтому никакого «гена супружеской неверности» быть в принципе не может. Может быть разве что предрасположенность, вклад которой зачастую заметно ниже вклада культуры, воспитания и обстоятельств.
Еще шире
Если вернуться к сексуальному поведению, то его связь — в том числе у самок мышей и обезьян разных видов — с полиморфизмом AVPR1A действительно была продемонстрирована несколько раз. Однако подобные данные можно переносить на человека лишь с очень существенными оговорками. В конце концов, два равноудаленных от нас вида шимпанзе, обычный и карликовый бонобо, демонстрируют очень разные репродуктивные стратегии.
Бонобо бисексуальны и главенствующую роль в группе зачастую играют самки. Бонобо, как правило, очень неудобный пример «естественности» для желающих обосновать то или иное сексуальное поведение людей как «унаследованное от природы». Хотя эти наши ближайшие родственники далеко не самые странные, с человеческой точки зрения: им далеко даже до пятнистых гиен, не говоря уж о кальмарах.
Пятнистая гиена в Кении. Снимок Joanne Goldby / WikimediaРепродуктивные органы пятнистых гиен долгое время сбивали с толку натуралистов: самки обладают псевдопенисом, который больше полового органа самцов. Во время родов новорожденные гиены проходят через влагалище, расположенное внутри псевдопениса. Причем с точки зрения анатомии этот орган является сильно увеличенным клитором. Пока не очень понятно, как такая необычная анатомия связана со строением и работой мозга, но у гиен в стаях царит матриархат: самый высокопоставленный самец ниже по положению любой самки.
У морских коньков привычная картина внутреннего оплодотворения выглядит еще более причудливо. Самка помещает внутрь самца яйцеклетку, которая потом делится и развивается внутри отцовского тела. А некоторые виды кальмаров для оплодотворения протыкают шипами тело самок, перенося при этом внутрь свою сперму. У крокодилов биологический пол не связан с генами, но определяется температурным режимом развития эмбриона; кукушки избавились от типичной для птиц заботы о потомстве; у голых землекопов нора организована почти как муравейник (с главной самкой, которая одна лишь дает потомство) — практически любую «естественную» закономерность в сексуальном или родительском поведении можно опровергнуть на примере того или иного животного.
Генетически люди родственны с гиенами более чем на 80%. С утконосами — на 69%, на 65% — с курами, а примерно четверть генов совпадает с виноградом.
Так что когда в очередной раз прочтете заголовок в духе «эксперименты на мышах позволили выявить ген супружеской неверности», помните, что такие заявления как минимум некорректны, а как максимум и вовсе являются передергиванием. Собственно, сами ученые почти всегда честно делают все необходимые оговорки, которые таинственным образом исчезают при пересказе.Алексей Тимошенко
Получены структуры веществ для эффективной борьбы против астмы
Новости онкологии
13 декабря 2019
Биофизики из Центра изучения молекулярных механизмов старения и возрастных заболеваний МФТИ в сотрудничестве с коллегами из Канады, США, Японии, Франции и Германии пролили свет на структуру и особенности работы рецепторов CysLT, регулирующих провоспалительные реакции, связанные с аллергическими расстройствами. Работа опубликована в журнале Nature Communications.
Рисунок. Общая структура CysLT2R (синий), а также ее сравнение со структурой CysL1R (желтый).
Источник: Anastasiia Gusach et al., Nature Communications.
«Клеточное общение» – ключевой этап жизнедеятельности, благодаря которому происходит запуск или приостановка клеточных процессов. Такое общение происходит с помощью рецепторов – молекул белков на поверхности клетки и клеточных органелл или же белков, растворенных в цитоплазме, которые способны специфично реагировать на присоединение к ним молекул определенного химического вещества. Вещество, специфически соединяющееся с рецептором, называется лигандом этого рецептора. Рецепторы, сопряженные с G-белком (сокращенно GPCR от G-protein-coupled receptors), – это белковые молекулярные машины, встроенные в мембрану клетки. CysLT1R и CysLT2R относятся к GPCR-рецепторам, а их лигандами являются липидные высокоактивные вещества, так называемые «пептидные (цистеиновые) лейкотриены» (сокращенно CysLT от Cysteinyl leukotrienes).
Обе подгруппы рецепторов – CysLT1 и CysLT2 – регулируют мобилизацию воспалительного ответа, вызванного аллергическими расстройствами. Более двух десятилетий для лечения астмы и сопутствующих заболеваний применяются препараты, подавляющие работу рецепторов CysLT1, однако множество пациентов такой терапии не поддаются. В то же время значение CysLT2R в физиологии и патогенезе воспалительных процессов мало изучено. Лишь недавно рецептор CysLT2 был предложен в качестве потенциальной лекарственной мишени против атопической астмы, травм головного мозга и расстройств центральной нервной системы, а также нескольких видов рака. В настоящий момент созданию более эффективных препаратов против астмы и сопутствующих заболеваний мешает нехватка информации о том, с какими лигандами и как связываются рецепторы CysLT, а также недостаточное понимание их механизмов действия – это требует данных высокого разрешения о структурном устройстве рецепторов. Получив такие данные с помощью методов структурной биологии, дальнейшее исследование ученые могут проводить при помощи компьютерного моделирования.
Метод молекулярного моделирования, целью которого является поиск наиболее достоверной ориентации и конформации лиганда в центре связывания рецептора, называется молекулярным докингом (от docking – стыковка). Данные о пространственной структуре рецептора, известной с разрешением в несколько ангстрем, а также о химической структуре лиганда позволяют предсказать пространственную структуру комплекса «рецептор – лиганд» и свободную энергию его образования. Знание пространственной структуры таких комплексов – ключевой момент на пути к пониманию механизмов их функционирования.
Рисунок. Схематическая иллюстрация докинга малой молекулы лиганда (синий)
с рецептором (красный). Источник: «Википедия».
В новой работе ученые из МФТИ выявили решающие факторы связывания с лигандами рецепторов CysLT1 и CysLT2 на основании данных проведенного ими структурного анализа CysLT2R, а также структурных данных о CysLT1R, опубликованных лабораторией ранее.
«Новые структуры позволили сделать докинг лигандов намного точнее и объяснить их свойства относительно каждого из рецепторов. Теперь мы знаем, как менять шаблон для дизайна лекарств так, чтобы подавлять работу рецепторов CysLT1 и CysLT2 одновременно или же селективно любого из них», – прокомментировала Анастасия Гусач, аспирантка МФТИ и младший научный сотрудник лаборатории структурной биологии рецепторов, сопряженных с G-белком, МФТИ.
В дальнейшем такое вещество может быть использовано в качестве лекарства или же инструмента, помогающего расшифровать значение каждого из подтипов рецептора CysLT при различных физиологических и патологических процессах.
«На основании недавно полученных результатов предполагается, что в случае, если мишенью для подавления лекарствами выбрать сразу оба рецептора CysLT1 и CysLT2 или же только CysLT2R, такое лечение будет более эффективным по сравнению с подавлением только CysLT1R, особенно при тяжелой астме. Кроме того, CysLT2R обещает быть перспективной мишенью против травм головного мозга и при нейродегенеративных расстройствах. Все вышесказанное делает изучение CysLT2R перспективным направлением. В данной работе мы описали четыре кристаллические структуры CysLT2R в комплексе с тремя лигандами, блокирующими действие рецепторов CysLT1 и CysLT2», – рассказал Алексей Мишин, старший научный сотрудник лаборатории структурной биологии рецепторов, сопряженных с G-белком, МФТИ.
Другое многообещающее применение структурной информации, полученной в ходе исследования, – возможность обоснования влияний точечных мутаций, в результате которых в аминокислотной последовательности белка появляется «чужая» аминокислота, что в свою очередь влияет на работу рецептора. Для этого исследователи собрали информацию о мутациях в последовательности рецептора CysLT2 от 60 000 здоровых людей и отобразили положения новых аминокислот на структуре рецептора. Оказалось, что около четверти мутаций находятся в функционально важных областях и могут влиять на то, какие процессы будет включать или выключать рецептор при взаимодействии со своим лигандом. Это значит, что в условиях стремительного развития геномного секвенирования и накопления большого объема статистических данных структурно-функциональные исследования позволят каждому желающему предсказать не только заболевания, но и влияние мутаций на действенность и безопасность лекарств.
Работа выполнена при поддержке Российского научного фонда и Министерства науки и высшего образования РФ.
Источник: mipt.ru
Ожирение у детей и подростков (недостаточность рецептора меланокортина)
Генетическое исследование, предназначенное для диагностики ожирения, ассоциированного с рецептором меланокортина 4-го типа. Мутации в гене MC4R являются наиболее распространенной генетической причиной тяжелого ожирения, проявляющегося в раннем возрасте.
Состав исследования
Синонимы английские
MC4R.
Метод исследования
Автоматическое секвенирование ДНК.
Генетические маркеры
Рецептор меланокортина 4 (MC4R) (все мутации).
Какой биоматериал можно использовать для исследования?
Буккальный (щечный) эпителий, венозную кровь.
Как правильно подготовиться к исследованию?
- Подготовки не требуется.
Общая информация об исследовании
До 6 % людей с тяжелым ожирением, развивающимся с детства, несут мутации в гене рецептора меланокортина 4-го типа (MC4R). Поэтому недостаточность MC4R – самая распространенная причина моногенного ожирения у человека.
Клиническая картина ожирения, связанного с недостаточностью рецептора меланокортина, может проявляться уже на первом году жизни:
- повышенный аппетит, переедание, отсутствие чувства сытости,
- быстрый набор веса (за счёт регулярного переедания),
- увеличение жировой, мышечной ткани сочетается с высоким ростом,
- как правило, не наблюдается нарушений функций щитовидной железы,
- половое развитие соответствует возрасту,
- задержек психомоторного развития, нарушения интеллекта нет,
- раннее развитие гиперинсулинемии, которая в 30 % случаев впоследствии приводит к сахарному диабету 2-го типа.
В зрелом возрасте это грозит метаболическим синдромом, сопутствующими сердечно-сосудистыми заболеваниями, сахарным диабетом 2-го типа, ретинопатией (невоспалительным поражением сетчатки глаза). Кроме того, люди с ожирением более склонны к развитию стеатоза печени, обструктивного апноэ во время сна (остановки дыхания), ортопедических осложнений.
Ген MC4R кодирует нейрональный меланокортиновый рецептор, который является фактором регуляции пищевого поведения (подавляющим аппетит) и энергетического баланса.
Экспрессия гена MC4R широко распространена в центральной нервной системе: таламусе, гипоталамусе, стволе мозга и коре. Распространение в этих регионах мозга рецепторов свидетельствует об их возможном участии в регуляции вегетативных и нейроэндокринных функций. Существуют данные, что MC4R экспрессируется также в спинном мозге.
Различные по степени тяжести формы ожирения, связанного с геном MC4R, являются следствием изменений последовательности гена, в результате чего белок – меланокортиновый рецептор – становится неактивен или менее активен. Тяжесть ожирения зависит от типа мутации и от того, находится ли она в гомозиготном или гетерозиготном состоянии.
Белок, кодируемый геном MC4R, формирует меланокортин, отвечающий за чувство насыщения. Соответственно, при мутации гена MC4R ребенок может переедать просто потому, что не способен контролировать аппетит по физиологическим причинам.
Ранняя диагностика причин ожирения позволяет существенно сократить его физические и психологические последствия. Родителям информация, что MC4R-ассоциированное ожирение у ребенка связано с тем, что у него проблемы с достижением сытости, указывает на необходимость с ранних лет регулировать пищевое поведение ребенка, следить за его массой тела.
В настоящее время несколько препаратов для целенаправленной терапии данного варианта ожирения проходят клинические испытания.
Генетическое исследование особенно важно для дифференциальной диагностики ожирения, вызванного мутацией гена MC4R, с другими формами ожирения; это необходимо для подбора адекватного метода терапии и определения прогноза заболевания.
Спектр мутаций в гене MC4R очень широк. Тест заключается в исследовании полной кодирующей области гена рецептора меланокортина 4 (MC4R) и позволяет выявить любые возможные мутации, приводящие к ожирению.
С помощью предлагаемого генетического исследования можно либо подтвердить недостаточность MC4R, либо исключить ее и перейти к поиску других причин, в том числе более редких форм наследственно обусловленного ожирения.
Конечно, даже наличие гена не означает, что у человека будет ожирение. Проявится ген или нет, зависит от образа жизни, пищевых привычек. Однако при любой наследственности ожирение легче предотвратить, чем потом избавляться от него.
Когда назначается исследование?
- При переедании у детей раннего возраста.
- При тяжелом ожирении с раннего возраста без нарушения полового и умственного развития.
- При дифференциальной диагностике с другими формами ожирения.
- Когда известно, что у родственников пациента было ожирение в раннем возрасте.
Что означают результаты?
Результатом генетического исследования является заключение о наличии или отсутствии мутаций гена MC4R;. выдается инф-ция о генотипе и наличии/отсутствии риска мутаций.
Скачать пример результатаТакже рекомендуется
- Ген, ассоциированный с жировой массой и ожирением (FTO). Выявление мутации G(45+52261)A (регуляторная область гена)
- Лептин
- Глюкоза в плазме
- Инсулин
- Гликированный гемоглобин (HbA 1c)
- Тиреотропный гормон (ТТГ)
- Тироксин свободный (Т4 свободный)
Литература
- Farooqi IS,et al. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N Engl J Med 2003; 348: 1085−1095.
- Lubrano-Berthelier C. et al., Melanocortin 4 receptor mutations in a large cohort of severely obese adults: prevalence, functional classification, genotype-phenotype relationship, and lack of association with binge eating, J Clin Endocrinol Metab. 2006.
Сенсорные рецепторы | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма.В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Содержание
- 1 Расположение
- 2 функции
- 3 Классификация по адекватному стимулу
- 4 Классификация по местонахождению
- 5 Классификация по морфологии
- 6 См. Также
- 7 Артикул
Сенсорный рецептор может быть специализированной частью плазматической мембраны или отдельной клеткой, связанной с окончанием нейрона.
Сенсорные рецепторы, отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители. Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы.Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию.
Классификация по адекватному стимулу [править | править источник]
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Электромагнитные рецепторы реагируют на инфракрасное излучение, видимый свет или магнитные поля. [1]
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на вредные или потенциально вредные раздражители
- Осморецепторы реагируют на осмолярность жидкостей (например, в гипоталамусе
- Фоторецепторы реагируют на свет
- Проприоцепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру, либо тепло, либо холод, либо и то, и другое.
Классификация по местоположению [править | править источник]
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Классификация по морфологии [править | править источник]
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы включают остальные типы кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
- Перехватчики
- Нейроанализаторы
- Проприоцепция
- Sense
- Сенсорный нейрон
- ↑ Майкл Дж. Грегори. Сенсорные системы. Общественный колледж Клинтона. URL-адрес, доступ к которому осуществлен 06.06.2013.
| Нервная система — Сенсорная система — редактировать |
|---|
| Особые чувства : Визуальная система | Слуховая система | Обонятельная система | Вкусовая система Соматосенсорная система : Ноцицепция | Терморецепция | Вестибулярная система | Механорецепция (давление, вибрация и проприоцепция) | Эквилибриоцепция |
| На этой странице используется содержимое Википедии под лицензией Creative Commons (просмотреть авторов). |
Сенсорные рецепторы | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма.В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Содержание
- 1 Расположение
- 2 функции
- 3 Классификация по адекватному стимулу
- 4 Классификация по местонахождению
- 5 Классификация по морфологии
- 6 См. Также
- 7 Артикул
Сенсорный рецептор может быть специализированной частью плазматической мембраны или отдельной клеткой, связанной с окончанием нейрона.
Сенсорные рецепторы, отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители. Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы.Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию.
Классификация по адекватному стимулу [править | править источник]
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Электромагнитные рецепторы реагируют на инфракрасное излучение, видимый свет или магнитные поля. [1]
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на вредные или потенциально вредные раздражители
- Осморецепторы реагируют на осмолярность жидкостей (например, в гипоталамусе
- Фоторецепторы реагируют на свет
- Проприоцепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру, либо тепло, либо холод, либо и то, и другое.
Классификация по местоположению [править | править источник]
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Классификация по морфологии [править | править источник]
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы включают остальные типы кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
- Перехватчики
- Нейроанализаторы
- Проприоцепция
- Sense
- Сенсорный нейрон
- ↑ Майкл Дж. Грегори. Сенсорные системы. Общественный колледж Клинтона. URL-адрес, доступ к которому осуществлен 06.06.2013.
| Нервная система — Сенсорная система — редактировать |
|---|
| Особые чувства : Визуальная система | Слуховая система | Обонятельная система | Вкусовая система Соматосенсорная система : Ноцицепция | Терморецепция | Вестибулярная система | Механорецепция (давление, вибрация и проприоцепция) | Эквилибриоцепция |
| На этой странице используется содержимое Википедии под лицензией Creative Commons (просмотреть авторов). |
Как нейроны общаются | Введение в психологию
Цели обучения
- Опишите, как нейроны взаимодействуют друг с другом
- Объясните, как лекарства действуют как агонисты или антагонисты данной системы нейротрансмиттеров
Теперь, когда мы узнали об основных структурах нейрона и о роли, которую эти структуры играют в нейронной коммуникации, давайте более подробно рассмотрим сам сигнал — то, как он проходит через нейрон, а затем переходит к следующему нейрону, где процесс повторяется.
Мы начинаем с нейрональной мембраны. Нейрон существует в жидкой среде — он окружен внеклеточной жидкостью и содержит внутриклеточную жидкость (т. Е. Цитоплазму). Мембрана нейрона разделяет эти две жидкости — критически важная роль, потому что электрический сигнал, проходящий через нейрон, зависит от того, насколько электрически различаются внутри- и внеклеточные жидкости. Эта разница в заряде через мембрану, называемая мембранным потенциалом , обеспечивает энергию для сигнала.
Электрический заряд жидкостей вызывается заряженными молекулами (ионами), растворенными в жидкости. Полупроницаемая природа нейрональной мембраны несколько ограничивает движение этих заряженных молекул, и в результате некоторые из заряженных частиц имеют тенденцию становиться более концентрированными внутри или снаружи клетки.
Между сигналами потенциал мембраны нейрона поддерживается в состоянии готовности, называемом потенциалом покоя . Подобно растянутой резинке, ожидающей начала действия, ионы выстраиваются в линию по обе стороны от клеточной мембраны, готовые устремиться через мембрану, когда нейрон становится активным и мембрана открывает свои ворота (т.например, натрий-калиевый насос , который позволяет перемещать ионы через мембрану). Ионы в областях с высокой концентрацией готовы перейти в области с низкой концентрацией, а положительные ионы готовы перейти в области с отрицательным зарядом.
В состоянии покоя натрий (Na + ) находится в более высоких концентрациях вне клетки, поэтому он будет стремиться перемещаться в клетку. Калий (K + ), с другой стороны, больше сконцентрирован внутри клетки и имеет тенденцию выходить из клетки (рис. 1).Кроме того, внутренняя часть элемента заряжена немного отрицательно по сравнению с внешней стороной. Это создает дополнительную силу для натрия, заставляя его перемещаться в клетку.
Рис. 1. При потенциале покоя Na + (синие пятиугольники) более высококонцентрирован вне клетки во внеклеточной жидкости (показан синим цветом), тогда как K + (фиолетовые квадраты) более высококонцентрирован возле мембраны в цитоплазма или внутриклеточная жидкость. Другие молекулы, такие как ионы хлорида (желтые кружки) и отрицательно заряженные белки (коричневые квадраты), способствуют созданию положительного чистого заряда во внеклеточной жидкости и отрицательного чистого заряда во внутриклеточной жидкости.
Из этого потенциального состояния покоя нейрон получает сигнал, и его состояние резко меняется (рис. 2). Когда нейрон получает сигналы от дендритов — из-за нейротрансмиттеров от соседнего нейрона, связывающегося с его рецепторами — на нейронной мембране открываются маленькие поры или ворота, позволяя ионам Na + , движущимся за счет разницы в заряде и концентрации, перемещаться в камеру. Благодаря этому притоку положительных ионов внутренний заряд клетки становится более положительным. Если этот заряд достигает определенного уровня, называемого порогом возбуждения , нейрон становится активным, и начинается потенциал действия.
Многие дополнительные поры открываются, вызывая массивный приток ионов Na + и огромный положительный всплеск мембранного потенциала, пикового потенциала действия. На пике пика натриевые вентили закрываются, а калиевые ворота открываются. Когда положительно заряженные ионы калия уходят, клетка быстро начинает реполяризацию. Сначала он гиперполяризуется, становясь немного более отрицательным, чем потенциал покоя, а затем выравнивается, возвращаясь к потенциалу покоя.
Рисунок 2.Во время действия потенциала электрический заряд на мембране резко меняется.
Этот положительный импульс составляет потенциал действия : электрический сигнал, который обычно движется от тела клетки вниз по аксону к терминалам аксона. Электрический сигнал движется вниз по аксону, как волна; в каждой точке некоторые из ионов натрия, которые входят в клетку, диффундируют к следующему участку аксона, поднимая заряд выше порога возбуждения и вызывая новый приток ионов натрия.Потенциал действия перемещается вниз по аксону к кнопкам терминала. Этот процесс объясняется в следующем видео:
Потенциал действия — это феномен «все или ничего» . Проще говоря, это означает, что входящего сигнала от другого нейрона либо достаточно, либо недостаточно для достижения порога возбуждения. Между ними нет и нет отключения потенциала действия после его запуска. Думайте об этом как об отправке электронного письма или текстового сообщения.Вы можете думать об отправке всего, что хотите, но сообщение не будет отправлено, пока вы не нажмете кнопку отправки. Более того, как только вы отправите сообщение, его уже не остановить.
Поскольку это все или ничего, потенциал действия воссоздается или распространяется с полной силой в каждой точке аксона. Подобно зажженному фитилю фейерверка, он не гаснет при движении по аксону. Именно это свойство «все или ничего» объясняет тот факт, что ваш мозг воспринимает травму удаленной части тела, например, пальца ноги, столь же болезненной, как травму носа.
Как отмечалось ранее, когда потенциал действия достигает конечной кнопки, синаптические пузырьки высвобождают свои нейротрансмиттеры в синапс . Нейромедиаторы перемещаются по синапсу и связываются с рецепторами на дендритах соседнего нейрона, и процесс повторяется в новом нейроне (при условии, что сигнал достаточно силен, чтобы запустить потенциал действия). Как только сигнал доставлен, избыточные нейротрансмиттеры в синапсе уносятся прочь, распадаются на неактивные фрагменты или реабсорбируются в процессе, известном как обратный захват. Reuptake вовлекает нейротрансмиттер, перекачиваемый обратно в нейрон, который его высвободил, чтобы очистить синапс (рис. 3). Очистка синапса служит как для обеспечения четкого состояния «включено» и «выключено» между сигналами, так и для регулирования выработки нейротрансмиттера (полные синаптические везикулы предоставляют сигналы, которые не требуют производства дополнительных нейротрансмиттеров).
Рис. 3. Обратный захват включает перемещение нейромедиатора из синапса обратно в терминал аксона, из которого он был выпущен.
Нейронную коммуникацию часто называют электрохимическим явлением. Движение потенциала действия по длине аксона является электрическим событием, а движение нейромедиатора через синаптическое пространство представляет собой химическую часть процесса.
Ссылка на обучение
Посмотрите следующее видео, чтобы увидеть, как нейроны взаимодействуют в организме.
Проверьте свое понимание нейронной коммуникации и потенциала действия в этом упражнении и анимации Гарвардского колледжа.
Нейротрансмиттеры и лекарственные препараты
Существует несколько различных типов нейротрансмиттеров , высвобождаемых разными нейронами, и мы можем в общих чертах говорить о видах функций, связанных с различными нейротрансмиттерами (Таблица 1). Многое из того, что психологи знают о функциях нейромедиаторов, основано на исследованиях воздействия наркотиков на психологические расстройства. Психологи, которые принимают биологическую перспективу и сосредотачиваются на физиологических причинах поведения, утверждают, что психологические расстройства, такие как депрессия и шизофрения, связаны с дисбалансом в одной или нескольких системах нейромедиаторов.С этой точки зрения психотропных препаратов могут помочь улучшить симптомы, связанные с этими расстройствами. Психотропные препараты — это препараты, которые лечат психические симптомы, восстанавливая баланс нейромедиаторов.
| Нейротрансмиттер | Участвует в | Потенциальное влияние на поведение |
|---|---|---|
| Ацетилхолин | Действие мышц, память | Повышенное возбуждение, улучшение познания |
| Бета-эндорфин | Боль, удовольствие | Снижение тревожности, снижение напряжения |
| Дофамин | Настроение, сон, обучение | Повышенное удовольствие, подавление аппетита |
| Гамма-аминомасляная кислота (ГАМК) | Функции мозга, сон | Снижение тревожности, снижение напряжения |
| Глутамат | Память, обучение | Улучшенное обучение, улучшенная память |
| Норэпинефрин | Сердце, кишечник, бдительность | Повышенное возбуждение, подавление аппетита |
| Серотонин | Настроение, сон | Регулировка настроения, подавление аппетита |
Психоактивные препараты могут действовать как агонисты или антагонисты данной системы нейротрансмиттеров. Агонисты — это химические вещества, которые имитируют нейромедиатор на участке рецептора и, таким образом, усиливают его эффекты. Антагонист , , с другой стороны, блокирует или препятствует нормальной активности нейромедиатора на рецепторе. Препараты-агонисты и антагонисты назначаются для коррекции дисбаланса конкретных нейромедиаторов, лежащих в основе состояния человека. Например, болезнь Паркинсона, прогрессирующее заболевание нервной системы, связано с низким уровнем дофамина. Поэтому агонисты дофамина, которые имитируют эффекты дофамина за счет связывания с дофаминовыми рецепторами, являются одной из стратегий лечения.
Определенные симптомы шизофрении связаны со сверхактивной нейротрансмиссией дофамина. Нейролептики, используемые для лечения этих симптомов, являются антагонистами дофамина — они блокируют действие дофамина, связывая его рецепторы, не активируя их. Таким образом, они не позволяют допамину, высвобождаемому одним нейроном, передавать информацию соседним нейронам.
В отличие от агонистов и антагонистов, которые действуют путем связывания с рецепторными участками, ингибиторы обратного захвата предотвращают транспортировку неиспользованных нейротрансмиттеров обратно в нейрон.Это оставляет больше нейротрансмиттеров в синапсе на более длительное время, увеличивая его эффекты. Депрессия, которая постоянно связана со снижением уровня серотонина, обычно лечится селективными ингибиторами обратного захвата серотонина (СИОЗС). Предотвращая обратный захват, СИОЗС усиливают действие серотонина, давая ему больше времени для взаимодействия с рецепторами серотонина на дендритах. Сегодня на рынке распространены такие СИОЗС, как Прозак, Паксил и Золофт. Препарат ЛСД структурно очень похож на серотонин и влияет на те же нейроны и рецепторы, что и серотонин.Психотропные препараты не являются мгновенным решением для людей, страдающих психологическими расстройствами. Часто человек должен принимать лекарство в течение нескольких недель, прежде чем заметит улучшение, а многие психоактивные препараты имеют серьезные отрицательные побочные эффекты. Кроме того, люди сильно различаются по тому, как они реагируют на лекарства. Чтобы повысить шансы на успех, люди, получающие фармакотерапию, нередко также проходят психологическую и / или поведенческую терапию. Некоторые исследования показывают, что сочетание лекарственной терапии с другими формами терапии, как правило, более эффективно, чем какое-либо отдельное лечение (один такой пример см. В March et al., 2007).
Ссылка на обучение
Просмотрите процесс нейронной коммуникации в следующем видео по психологии CrashCourse:
Глоссарий
потенциал действия: электрический сигнал, который движется вниз по аксону нейрона
Агонист : препарат , имитирующий или усиливающий эффекты нейромедиатора
all-or-none: явление, при котором входящий сигнал от другого нейрона либо достаточен, либо недостаточен для достижения порога возбуждения
антагонист: лекарственное средство, которое блокирует или препятствует нормальной активности данного нейромедиатора
биологическая перспектива: точка зрения, что психологические расстройства, такие как депрессия и шизофрения, связаны с дисбалансом в одной или нескольких системах нейромедиаторов
мембранный потенциал: разность зарядов через нейрональную мембрану
нейрон: клеток нервной системы, которые действуют как взаимосвязанные информационные процессоры, которые необходимы для всех задач нервной системы
нейромедиатор: химический посланник нервной системы
психотропные препараты: лекарств, которые лечат психические симптомы путем восстановления баланса нейромедиаторов
рецептор : белок на поверхности клетки, где прикрепляются нейротрансмиттеры
потенциал покоя: состояние готовности мембранного потенциала нейрона между сигналами
обратный захват: нейромедиатор перекачивается обратно в нейрон, который его выпустил
полупроницаемая мембрана: клеточная мембрана, которая позволяет более мелким молекулам или молекулам без электрического заряда проходить через нее, останавливая при этом более крупные или сильно заряженные молекулы
синапс: небольшой промежуток между двумя нейронами, где происходит коммуникация
синаптический пузырек: место хранения нейромедиаторов
терминальная кнопка: терминал аксона, содержащий синаптические пузырьки
порог возбуждения: уровень заряда в мембране, который вызывает активацию нейрона
Прикосновение и боль | Введение в психологию
Потенциалы действия в рецепторных клетках перемещаются как нервные импульсы с разной скоростью
Когда вы наступаете на булавку, это активирует множество механорецепторов, многие из которых являются ноцицепторами.Вы могли заметить, что ощущение со временем меняется. Сначала вы чувствуете резкий удар, который заставляет вас убрать ногу, и только потом вы чувствуете волну еще более ноющей боли. Об остром ударе сигнализирует быстро проводящие А-волокна, которые проецируются в соматосенсорную кору. Эта часть коры соматотопически организована, то есть сенсорные сигналы представлены в зависимости от того, откуда они берутся в теле (см. Иллюстрацию гомункула, рис. 2). Неприятная боль, которую вы чувствуете после укола острым булавкой, — это отдельный одновременный сигнал, который посылается от ноцицепторов в вашей стопе через тонкие C-боли или Aδ-волокна в кору островка и другие области мозга, участвующие в обработке эмоций и интероцепции (см. Рис. 3а для схематического изображения этого пути).Другими словами, опыт наступления на булавку складывается из двух отдельных сигналов: один различающий сигнал, позволяющий нам локализовать сенсорный стимул и различать, тупой это или острый удар; и один аффективный сигнал, который сообщает нам, что наступать на булавку — плохо. Боль принято разделять на сенсорно-дискриминационный и аффективно-мотивационный аспекты (Auvray, Myin, & Spence, 2010). Это различие соответствует, по крайней мере частично, тому, как эта информация передается от периферии к центральной нервной системе и как она обрабатывается в головном мозге (Price, 2000).
Рисунок 2 . Гомункул: Гомункул означает «маленький человек», и здесь вы видите масштабную модель человеческого тела, искаженную, чтобы отразить относительное пространство, которое части тела занимают в соматосенсорной коре. Как видите, губы, руки, ступни и гениталии посылают в мозг больше соматосенсорных проекций, чем любые другие части тела. Рисунок 2b. Кортикальное картирование сенсорного гомункула: части тела представлены в определенных местах соматосенсорной коры. Представления отображаются соматотопически: ступни расположены медиально, а плечи и руки — латеральнее межполушарной щели.Лицевые структуры представлены в другом месте, нежели на коже черепа и головы; лицо ориентировано «вверх ногами», лоб обращен к плечам.
Боль необходима для выживания, но наш мозг может ее остановить, если потребуется
В апреле 2003 года альпинист Арон Ралстон оказался на дне каньона Блю Джон в штате Юта, вынужденный сделать ужасный выбор: встретить медленную, но верную смерть — или ампутировать правую руку. Пятью днями ранее он упал в каньон — с тех пор его правая рука застряла между валуном весом 800 фунтов и крутой стеной из песчаника.Ослабев от нехватки еды и воды и близок к тому, чтобы сдаться, ему пришло в голову, как прозрение, что, если он сломает две кости в предплечье, ему удастся отрезать остальные своим карманным ножом. Мысль о том, чтобы освободиться и выжить, привела его в такое возбуждение, что он провел следующие 40 минут полностью поглощенный задачей: сначала сломал кости, используя свое тело в качестве рычага, затем сунул пальцы в руку, сжимая пучки мышечных волокон и разрывая их. одну за другой, прежде чем перерезать синие артерии и бледные, похожие на лапшу нервы.Боль была неважной. Только прорезание толстого белого главного нерва заставило его остановиться на минуту — поток боли, как он описывает, был как будто его всю руку толкали «в котел с магмой». Наконец, освободившись, он спустился со скалы и прошел еще 7 миль, пока его не спасли туристы (Ralston, 2010).
Как возможно сделать что-то настолько мучительно болезненное для себя, как это сделал Арон Ральстон, и после этого все еще умудряться ходить, говорить и мыслить рационально? Ответ кроется в мозге, где интерпретируются сигналы тела.Когда мы воспринимаем соматосенсорные и ноцицептивные сигналы от тела, этот опыт очень субъективен и податлив в зависимости от мотивации, внимания, эмоций и контекста.
Рисунок 3 . Пути обработки боли. Слева — восходящие болевые пути: о травме одновременно сигнализируют быстро проводящие Aα или Aβ-волокна и медленно проводящие C-боли или Aδ-волокна. Быстрые А-волокна сигнализируют о давлении, растяжении и других движениях ткани в соматосенсорную кору через ядра спинного столба.С-боль и Aδ-волокна отправляют информацию о боли от ноцицепторов в ткани или коже и передают эти сигналы нейронам второго порядка в спинном роге спинного мозга. Затем нейроны второго порядка переходят на противоположную сторону, где они образуют восходящий спиноталамический тракт. Этот тракт передает сигналы ядрам в продолговатом и среднем мозге на пути к таламусу (Т). Таламус передает информацию соматосенсорной и островной коре, а также кортикальным областям, опосредующим различные аспекты переживания боли, такие как аффективные реакции в поясной коре головного мозга.Справа — нисходящие пути модуляции боли: информация из окружающей среды и определенные мотивационные состояния могут активировать этот путь сверху вниз. Несколько областей лимбического отдела переднего мозга, включая переднюю поясную извилину и островную кору, ядра миндалины и гипоталамуса (H), проецируются в периакведуктальный серый (PAG) среднего мозга, который затем модулирует восходящую передачу боли от афферентной системы боли косвенно через ростральную вентромедиальный мозг (RVM) в стволе мозга. Эта модулирующая система производит анальгезию за счет высвобождения эндогенных опиоидов и использует ON- и OFF-клетки для осуществления либо тормозящего (зеленый), либо облегчающего (красный) контроля ноцицептивных сигналов в спинном роге спинного мозга.
Модель мотивации – решения и нисходящая модуляция боли
Согласно модели мотивации-решения , мозг автоматически и непрерывно оценивает плюсы и минусы любой ситуации, взвешивая надвигающиеся угрозы и доступные награды (Fields, 2004, 2006). Все, что важнее для выживания, чем избегание боли, активирует нисходящую модуляторную систему боли в головном мозге — нисходящую систему, включающую несколько частей мозга и ствола мозга, которая подавляет ноцицептивную передачу сигналов, чтобы можно было уделить внимание более важным действиям.
В крайнем случае Арона, его действия, вероятно, были основаны на таком бессознательном процессе принятия решений — принимая во внимание его гомеостатическое состояние (его голод, жажда, воспаление и разложение его раздавленной руки, медленно влияющие на остальную часть его тела), сенсорный ввод доступный (сладкий запах растворяющейся кожи, тишина вокруг него, указывающая на его одиночество) и его знания об угрозах, с которыми он сталкивается (смерть или мучительная боль, которая не убьет его), по сравнению с потенциальными наградами (выживание, встречи с семьей очередной раз).История Арона иллюстрирует эволюционное преимущество способности отключать боль: нисходящая модуляторная система боли позволяет нам выполнять действия, которые могут спасти жизнь.
Однако, когда кто-то достиг безопасности или получил награду, исцеление становится более важным. Та же самая нисходящая система может затем «вызвать» болевые ощущения от тела, чтобы способствовать исцелению и мотивировать нас избегать потенциально болезненных действий. Чтобы облегчить или подавить ноцицептивные сигналы от тела, нисходящая система модуляции боли использует набор ВКЛЮЧЕННЫХ или ВЫКЛЮЧЕННЫХ клеток в стволе мозга, которые регулируют, какая часть ноцицептивного сигнала достигает мозга.Нисходящая система зависит от передачи сигналов опиоидов, и анальгетики, такие как морфин, снимают боль через этот контур (Petrovic, Kalso, Petersson, & Ingvar, 2002).
Обезболивающая сила вознаграждения
Размышления о хороших вещах, таких как его близкие и предстоящая жизнь, были, вероятно, решающими для выживания Арона. Обещания награды может быть достаточно, чтобы облегчить боль. Ожидание облегчения боли (уменьшение боли часто является наилучшим возможным результатом, если вы испытываете боль, т. Е. Это награда) от лечения, способствует эффекту плацебо, когда облегчение боли происходит, по крайней мере, частично за счет нисходящей модуляции вашего мозга. цепи, и такое облегчение зависит от собственной опиоидной системы мозга (Eippert et al., 2009; Eippert, Finsterbusch, Bingel, & Buchel, 2009; Левин, Гордон и Филдс, 1978). Вкусная еда, прослушивание хорошей музыки или приятные прикосновения к коже также уменьшают боль как у животных, так и у людей, предположительно, благодаря тому же механизму в мозге (Leknes & Tracey, 2008).
В уже ставшем классическим эксперименте Дам и Герц (1984) либо кормили крыс нормальной крысиной едой, либо позволяли им лакомиться очень полезными конфетами в шоколаде (крысы любят сладости), стоя на металлической тарелке, пока они не узнали, чего именно ожидать, когда их поместят. там.Когда тарелку нагревали до вредного / болезненного уровня, крысы, ожидавшие леденцов, выдерживали температуру в два раза дольше, чем крысы, ожидающие нормальной еды. Более того, этот эффект полностью исчез, когда опиоидная (эндорфиновая) система крыс была заблокирована лекарством, что указывает на то, что обезболивающий эффект ожидания вознаграждения был вызван высвобождением эндорфина.
Для Арона-альпиниста стресс от осознания надвигающейся смерти и ожидание награды за выживание, вероятно, наводнили его мозг эндорфинами, что способствовало волне возбуждения и эйфории, которую он испытал, выполняя ампутацию ». как пятилетний ребенок, получивший рождественские подарки »(Ralston, 2010).Это изменило его переживание боли от сильного повреждения тканей, которое он причинял, и позволило ему сосредоточиться на освобождении. Оказывается, наш мозг может модулировать восприятие того, насколько неприятна боль, сохраняя при этом способность ощущать интенсивность ощущения (Rainville, Duncan, Price, Carrier, & Bushnell, 1997; Rainville, Feine, Bushnell, & Дункан, 1992). Социальные награды, например, держание за руку парня или подруги, обладают обезболивающим эффектом. Даже просмотр его фотографии может иметь аналогичные эффекты — на самом деле, видение фотографии человека, с которым мы чувствуем близость, не только снижает субъективную оценку боли, но также снижает активность связанных с болью областей мозга (Eisenberger et al., 2011). Таким образом, наиболее распространенные действия, которые следует делать, когда вы хотите помочь кому-то пережить болезненный опыт, — присутствовать и держать человека за руку, — таким образом, они имеют ощутимый положительный эффект.
Сила разума
Контекст боли и прикосновения имеет большое влияние на то, как мы их интерпретируем. Только представьте, как иначе было бы Арону, если бы кто-нибудь ампутировал ему руку против его воли и без видимой причины. Продолжительную боль от травм будет легче перенести, если инцидент, вызвавший их, дает положительный контекст — например, военная рана, свидетельствующая о храбрости и целеустремленности солдата, — или фантомная боль от руки, отрезанной для продолжения жизни.
Относительное значение боли проиллюстрировано недавним экспериментом, в котором одно и то же умеренно болезненное тепло применялось к участникам в двух разных контекстах — в одном контрольном контексте, где альтернативой было безболезненное тепло; и другой, где альтернативой была очень болезненная жара. В контексте контроля, где умеренная жара была наименее предпочтительным исходом, она (неудивительно) была оценена как болезненная. В другом контексте это был наилучший возможный результат, и здесь точно такая же умеренно болезненная жара была фактически оценена как приятная — потому что это означало, что очень болезненной жары удалось избежать.Это несколько неожиданное изменение восприятия — когда боль становится приятной, потому что она представляет собой облегчение от чего-то худшего — подчеркивает важность того значения, которое люди приписывают своей боли, что может иметь решающее влияние на лечение боли (Leknes et al., 2013). В случае прикосновения, знание того, кто или что гладит вашу кожу, может иметь решающее значение — попробуйте подумать о слизнях в следующий раз, когда кто-то поглаживает вашу кожу, если вы хотите проиллюстрировать этот момент.
У боли и удовольствия есть общие модуляторные системы — еще один общий атрибут состоит в том, что нам не обязательно принимать их сами, чтобы испытать их.Что вы почувствовали, когда прочитали об Ароне, разрезавшем свои собственные ткани, или о том, что «Томас» бессознательно разрушил свои собственные кости? Вы съежились? Вполне вероятно, что некоторые области вашего мозга, обрабатывающие аффективные аспекты боли, были активны, хотя ноцицепторы в вашей коже и глубоких тканях не срабатывали. Боль может ощущаться косвенно, как и зуд, приятные прикосновения и другие ощущения. Таня Сингер и ее коллеги обнаружили в исследовании фМРТ, что некоторые из тех же областей мозга, которые были активны, когда участники чувствовали боль на собственной коже (передняя поясная извилина и островок), также были активны, когда им давали сигнал о том, что любимый человек чувствует боль.Те, кто был наиболее «чутким», также показали самые сильные реакции мозга (Singer et al., 2004). Аналогичный эффект был обнаружен для приятного прикосновения: задний островок у участников, просматривающих видео, где нежно поглаживают чью-то руку, демонстрирует такую же активацию, как если бы они сами получали прикосновение (Morrison, Bjornsdotter, & Olausson, 2011).
Определение сенсорного рецептора в психологии.
Примеры сенсорных рецепторов в следующих разделах:
Введение в Sensation
- Ощущение включает передачу информации от сенсорных рецепторов в мозг и позволяет человеку познавать окружающий мир.
- Сенсорная информация (например, вкус, свет, запах, давление, вибрация, тепло и боль) воспринимается телом сенсорными рецепторами .
- Эти сенсорные рецепторы включают глаза, уши, рот, нос, руки и ноги (и кожу в целом).
- Специализированные клетки сенсорных рецепторов преобразуют поступающую энергию (например, свет) в нервные импульсы.
- Объясните, как мозг и сенсорные рецепторы работают вместе в процессе ощущения
Сенсорная адаптация
- Сенсорная адаптация — это снижение отзывчивости сенсорной системы , которая сталкивается с постоянным стимулом.
- Сенсорная адаптация , также называемая нейронной адаптацией, представляет собой изменение реакции сенсорной системы , которая сталкивается с постоянным стимулом.
- Одним из примеров сенсорной адаптации является длительное прикосновение.
- Когда вы кладете руки на стол или надеваете одежду на тело, сначала сенсорные рецепторы распознают, что они активируются, и вы почувствуете ощущение прикосновения к объекту.
- Однако после продолжительного воздействия сенсорные рецепторы больше не будут активироваться так сильно, и вы больше не будете осознавать, что вы что-то касаетесь.
Сенсорная память
- Сенсорная память позволяет людям сохранять впечатления от сенсорной информации в течение короткого времени после того, как первоначальный стимул исчез.
- Сенсорная память — это автоматическая реакция, которая, как считается, выходит за рамки когнитивного контроля.
- Эхогенная память — это ветвь сенсорной памяти , используется слуховой системой.
- Тактильная память — это ветвь сенсорной памяти , используется осязанием.
- Сенсорные рецепторы по всему телу улавливают такие ощущения, как давление, зуд и боль, которые ненадолго сохраняются в тактильной памяти, прежде чем исчезнут или перенесены в кратковременную память.
Сенсорные абсолютные пороги
- В нейробиологии и психофизике существует несколько типов сенсорного порога .
- Однако, возможно, наиболее важным сенсорным порогом является абсолютный порог, который является наименьшим обнаруживаемым уровнем стимула.
- Сенсорная адаптация происходит, когда наши органы чувств больше не воспринимают стимул из-за постоянного контакта с ним нашего сенсорного рецептора .
- Если вы когда-либо входили в комнату с ужасным запахом, но через несколько минут понимали, что почти не замечаете этого, значит, вы испытали сенсорную адаптацию .
- Объясните, что такое абсолютный порог чувствительности и как на него можно повлиять
Знакомство с нервной системой
- Нервная система контролирует функции тела, собирая сенсорную информацию, вводя, интегрируя эту информацию внутри и сообщая надлежащую моторную мощность.
- Общий поток информации заключается в том, что периферическая нервная система (ПНС) принимает информацию через сенсорных нейронов, а затем отправляет ее в центральную нервную систему (ЦНС) для обработки.
- Основная функция ЦНС — интеграция и обработка сенсорной информации .
- Он синтезирует сенсорный входной сигнал для вычисления соответствующей двигательной реакции или выходного сигнала.
- Он состоит из сенсорных рецепторов , которые обрабатывают изменения внутренних и внешних раздражителей и передают эту информацию в ЦНС.
Этапы потенциала действия
- Центральная нервная система (ЦНС) во время своей работы проходит трехэтапный процесс: сенсорный ввод, ввод, нейронная обработка и двигательный вывод.
- Сенсорная входная ступень — это когда нейроны (или возбудимые нервные клетки) сенсорных органов возбуждаются электрически.
- Нервные импульсы от сенсорных рецепторов отправляются в головной и спинной мозг для обработки.
- Нейрон влияет на другие нейроны, высвобождая нейромедиатор, который связывается с химическими рецепторами .
- В этом отличие от рецепторных потенциалов , амплитуды которых зависят от интенсивности стимула.
Соматосенсорное ощущение: давление, температура и боль
- Рецепторные клетки в мышцах и суставах, называемые проприорецепторами, также помогают в соматосенсорной системе, но иногда их разделяют на другую сенсорную категорию , называемую кинестезией.
- Сенсорная функция клеток в соматосенсорной системе определяется местоположением.
- Мышцы Рецепторы наиболее активны в крупных суставах, таких как тазобедренные и коленные суставы, в то время как суставные и кожные рецепторы более значимы для суставов пальцев рук и ног. Механорецепторы
- могут быть свободными рецепторами или инкапсулированными.
- Примерами свободных рецепторов являются рецепторы волос у корней волос, в то время как инкапсулированные рецепторы представляют собой тельца Пачини и рецепторы в гладкой (безволосой) коже: тельца Мейснера, тельца Руффини и диски Меркеля. .
Вкус: вкусовые рецепторы и вкус
- Язык — главный сенсорный орган вкусовой системы.
- Каждый из них содержит вкусовую пору на поверхности языка, которая является местом сенсорной трансдукции .
- Солевой рецептор , NaCl, можно утверждать, что это самый простой из всех рецепторов , обнаруженных во рту.
- Три разных белка рецептора работают с кислым вкусом.
- Считается, что рецепторы умами действуют аналогично рецепторам горького и сладкого (включая GPCR), но об их действительной функции известно очень мало.
Центральная нервная система (ЦНС)
- Центральная нервная система состоит из головного и спинного мозга, которые обрабатывают сенсорный ввод и выдают инструкции телу.
- На этом этапе процесса головной и спинной мозг выбирает соответствующую моторную мощность, которая вычисляется на основе типа входного сенсорного сигнала .
- В нем находятся нервные центры, отвечающие за координацию сенсорной и двигательных систем в организме.
- Он регулирует моторную функцию и позволяет моторной и сенсорной информации передаваться от мозга к остальным частям тела.
- Импульсы отправляются от рецепторов через спинной мозг в головной мозг, где они обрабатываются и синтезируются в инструкции для остального тела.
Обоняние: носовая полость и запах
- Восприятие сложных ароматов требует одновременного распознавания вкусовых и обонятельных ощущений, взаимодействия, известного как хеморецептивное сенсорное взаимодействие.
- Однако обоняние анатомически отличается от вкуса, поскольку оно использует сенсорные органы носа и носовой полости для улавливания запахов.
- Каждый из 350 типов рецепторов характерен только для одного типа одоранта.
- У млекопитающих обонятельные рецепторы передают сигнал через G-белок.
- Это аналогичный тип передачи сигналов других известных сопряженных с G-белком рецепторов (GPCR).
5.4 Дегустация, обоняние и прикосновение — Введение в психологию — 1-е канадское издание
Цели обучения
- Обобщите, как чувства вкуса и обоняния преобразуют стимулы в восприятие.
- Опишите процесс трансдукции посредством осязания и проприоцепции.
- Изложите теорию контроля боли через врата. Объясните, почему боль так важна и как ее можно контролировать.
Хотя зрение и слух на сегодняшний день являются наиболее важными органами чувств, человеческие ощущения дополняются четырьмя другими, каждое из которых обеспечивает важный путь к лучшему пониманию окружающего мира и реакции на него.Этими другими чувствами являются осязание , вкус и запах , а также наше чувство положения и движения тела (проприоцепция) .
Дегустация
Вкус важен не только потому, что он позволяет нам наслаждаться едой, которую мы едим, но, что еще более важно, потому, что он ведет нас к продуктам, обеспечивающим энергию (например, сахар), и отдаляет от продуктов, которые могут быть вредными. Многие дети разборчивы в еде по одной причине — они биологически предрасположены очень осторожно относиться к тому, что они едят.Вместе с обонянием вкус помогает нам поддерживать аппетит, оценивать потенциальные опасности (например, запах утечки газа или горящего дома) и избегать употребления ядовитой или испорченной пищи.
Наша способность ощущать вкус начинается со вкусовых рецепторов на языке. Язык распознает шесть различных вкусовых ощущений, известных соответственно как сладкий, соленый, кислый, горький, пикантный (пряный) и умами (острый) . Умами — это мясной вкус, связанный с мясом, сырами, соей, водорослями и грибами, и он особенно присутствует в глутамате натрия (MSG), популярном усилителе вкуса (Ikeda, 1909/2002; Sugimoto & Ninomiya, 2005).
Наши языки покрыты вкусовыми рецепторами , которые предназначены для улавливания химикатов во рту . Большинство вкусовых рецепторов расположено в верхних внешних краях языка, но есть также рецепторы на задней части языка, а также на стенках рта и задней части глотки. Когда мы пережевываем пищу, она растворяется и попадает во вкусовые рецепторы, вызывая нервные импульсы, которые передаются в мозг (Northcutt, 2004). Человеческие языки покрыты от 2 000 до 10 000 вкусовых рецепторов, каждая из которых содержит от 50 до 100 вкусовых рецепторных клеток.Вкусовые рецепторы активируются очень быстро; соленый или сладкий вкус, который затрагивает вкусовые рецепторы даже на одну десятую секунды, вызывает нервный импульс (Kelling & Halpern, 1983). В среднем вкусовые рецепторы живут около пяти дней, после чего вместо них создаются новые вкусовые рецепторы. Однако по мере того, как мы становимся старше, скорость создания уменьшается, что делает нас менее чувствительными к вкусу. Это изменение помогает объяснить, почему некоторые продукты, которые в детстве кажутся такими неприятными, приносят больше удовольствия во взрослом возрасте.
Область сенсорной коры, которая реагирует на вкус, находится в очень похожем месте с областью, которая реагирует на запах, факт, который помогает объяснить, почему обоняние также влияет на наше восприятие того, что мы едим.Возможно, вы помните, что у вас были проблемы с дегустацией еды, когда вы сильно простудились, и если вы закроете нос и попробуете ломтики сырого картофеля, яблока и пастернака, вы не сможете почувствовать разницу между ними. Наше ощущение текстуры пищи (то, как мы ощущаем ее на языке) также влияет на то, как мы ее пробуем.
Запах
Когда мы вдыхаем воздух через ноздри, мы вдыхаем находящиеся в воздухе химические молекулы, которые обнаруживаются 10-20 миллионами рецепторных клеток , встроенных в обонятельную мембрану верхнего носового хода .Обонятельные рецепторные клетки имеют на вершине щупальцеобразные выступы, которые содержат рецепторные белки . Когда рецептор запаха стимулируется, мембрана посылает нейронные сообщения по обонятельному нерву в мозг (см. Рис. 5.20. «Обонятельные рецепторы»).
Рисунок 5.20. Обонятельные рецепторы. В обонятельной мембране более 1000 типов рецепторных клеток.У нас есть приблизительно 1000 типов рецепторных клеток запаха (Bensafi et al., 2004), и, по оценкам, мы можем обнаружить 10 000 различных запахов (Malnic, Hirono, Sato, & Buck, 1999).Рецепторы бывают разных форм и избирательно реагируют на разные запахи. Подобно замку и ключу, разные химические молекулы подходят к разным рецепторным клеткам, и запахи обнаруживаются в зависимости от их влияния на комбинацию рецепторных клеток. Подобно тому, как 10 цифр от 0 до 9 могут сочетаться множеством различных способов для создания бесконечного набора телефонных номеров, молекулы запаха связываются с различными комбинациями рецепторов, и эти комбинации декодируются в обонятельной коре головного мозга. Как видно на рисунке 5.21, «Возрастные различия в обонянии», обоняние достигает пика в раннем взрослом возрасте, а затем начинает медленно снижаться. К 60-70 годам обоняние резко ухудшается. Кроме того, у женщин, как правило, более острое обоняние, чем у мужчин.
Рисунок 5.21 Возрастные различия обоняния. Способность идентифицировать распространенные запахи заметно снижается в возрасте от 20 до 70 лет.Прикосновение
Чувство осязания необходимо для развития человека. Младенцы процветают, когда их обнимают и заботятся, но не тогда, когда они лишены человеческого контакта (Baysinger, Plubell, & Harlow, 1973; Feldman, 2007; Haradon, Bascom, Dragomir, & Scripcaru, 1994).Прикосновение передает тепло, заботу и поддержку и является неотъемлемой частью удовольствия, которое мы получаем от нашего социального взаимодействия с близкими людьми (Field et al., 1997; Keltner, 2009).
Кожа, самый большой орган в теле, является сенсорным органом. Кожа содержит множество нервных окончаний, комбинации которых реагируют на определенные типы давления и температуры. Прикоснувшись к разным частям тела, вы обнаружите, что некоторые области более чувствительны к щекотке, тогда как другие области больше реагируют на боль, холод или тепло.
Тысячи нервных окончаний в коже реагируют на четыре основных ощущения — давление, горячее, холодное и боль — но только ощущение давления имеет свои собственные специализированные рецепторы. Другие ощущения создаются комбинацией четырех других. Например:
- Ощущение щекотки вызвано раздражением соседних рецепторов давления.
- Ощущение жара вызывается стимуляцией рецепторов горячего и холода.
- Ощущение зуда вызвано многократной стимуляцией болевых рецепторов.
- Ощущение влажности вызвано многократным раздражением рецепторов холода и давления.
Кожа важна не только для получения информации о прикосновении и температуре, но и для проприоцепции — для способности ощущать положение и движение частей нашего тела . Проприоцепция осуществляется специализированными нейронами, расположенными в коже, суставах, костях, ушах и сухожилиях, которые посылают сообщения о сжатии и сокращении мышц по всему телу.Без этой обратной связи со стороны наших костей и мышц мы не смогли бы заниматься спортом, ходить или даже стоять.
Способность отслеживать, где движется тело, также обеспечивается вестибулярной системой , — набором заполненных жидкостью областей во внутреннем ухе, которые отслеживают положение и движение головы, поддерживая равновесие тела . Как вы можете видеть на Рисунке 5.22, «Вестибулярная система», вестибулярная система включает полукружные каналы и вестибулярные мешочки .Эти мешочки соединяют каналы с улиткой. Полукружные каналы воспринимают вращательные движения тела, а вестибулярные мешки воспринимают линейные ускорения. Вестибулярная система посылает сигналы нервным структурам, которые контролируют движение глаз, и мышцам, которые удерживают тело в вертикальном положении.
Рис. 5.22. Вестибулярная система. Вестибулярная система включает полукружные каналы (коричневые), которые передают вращательные движения тела, и вестибулярные мешки (синие), которые воспринимают линейные ускорения.Испытывая боль
Нам это не нравится, но переживание боли — это то, как тело сообщает нам, что мы в опасности. Ожог, когда мы касаемся горячего радиатора, и острый удар, когда мы наступаем на гвоздь, заставляют нас изменить свое поведение, предотвращая дальнейшее повреждение нашего тела. Люди, которые не могут испытывать боль, подвергаются серьезной опасности получить повреждения от ран, которые другие, страдающие болью, быстро заметят и займутся лечением.
Теория боли , управляющая воротами, предполагает, что боль определяется работой двух типов нервных волокон в спинном мозге .Один набор меньших нервных волокон передает боль от тела к мозгу, тогда как второй набор более крупных волокон предназначен для остановки или запуска (как ворота) потока боли (Melzack & Wall, 1996). Именно по этой причине массаж области, где вы чувствуете боль, может помочь облегчить ее — массаж активирует большие нервные волокна, которые блокируют болевые сигналы мелких нервных волокон (Wall, 2000).
Однако испытать боль намного сложнее, чем просто ответить на нейронные сообщения.Это тоже вопрос восприятия. Мы меньше чувствуем боль, когда заняты сложной деятельностью (Bantick et al., 2002), что может помочь объяснить, почему спортивные игроки могут чувствовать свои травмы только после игры. Мы также чувствуем меньше боли, когда нас отвлекает юмор (Zweyer, Velker, & Ruch, 2004). И боль успокаивается за счет выработки мозгом эндорфинов, естественных гормональных болеутоляющих. Высвобождение эндорфинов может объяснить эйфорию, испытываемую при беге на марафон (Sternberg, Bailin, Grant, & Gracely, 1998).
Основные выводы
- Способность ощущать вкус, запах и осязание важна, потому что помогает нам избежать вреда от токсинов окружающей среды.
- Множество вкусовых рецепторов на нашем языке и во рту позволяют нам различать шесть основных вкусовых ощущений: сладкий, соленый, кислый, горький, пикантный и умами.
- При обонянии трансдукция происходит, когда находящиеся в воздухе химические вещества, которые вдыхаются через ноздри, обнаруживаются рецепторами обонятельной мембраны. Различные химические молекулы вписываются в разные рецепторные клетки, создавая разные запахи.
- Обоняние снижается с возрастом, и в среднем у женщин обоняние лучше, чем у мужчин.
- У нас есть ряд различных нервных окончаний, встроенных в кожу, комбинации которых реагируют на четыре основных ощущения давления: жара, холода и боли. Но только ощущение давления имеет свои специализированные рецепторы.
- Проприоцепция — это наша способность ощущать положение и движения частей тела. Информация о позе и движении обнаруживается специальными нейронами, расположенными в коже, суставах, костях, ушах и сухожилиях, которые улавливают сообщения от сжатия и сокращения мышц по всему телу.
- Вестибулярная система, состоящая из структур внутреннего уха, контролирует положение и движения головы, поддерживая равновесие тела. Теория управления воротами
- объясняет, как большие и маленькие нейроны работают вместе, чтобы передавать и регулировать поток боли в мозг.
Упражнения и критическое мышление
- Подумайте о продуктах, которые вы больше всего любите есть. Какие из шести вкусовых ощущений вызывают эти продукты и почему, по вашему мнению, вам нравятся именно эти вкусы?
- Как вы думаете, почему у женщин может быть более развитое обоняние, чем у мужчин?
- Почему переживание боли приносит пользу людям?
Список литературы
Бантик, С.Дж., Уайз, Р. Г., Плогхаус, А., Клэр, С., Смит, С. М., и Трейси, И. (2002). Визуализация того, как внимание модулирует боль у людей, с помощью функциональной МРТ. Мозг: журнал неврологии, 125 (2), 310–319.
Байзингер, К. М., Плубелл, П. Э. и Харлоу, Х. Ф. (1973). Суррогатная мать с переменной температурой для изучения привязанности у детенышей обезьян. Методы исследования поведения и инструменты, 5 (3), 269–272.
Бенсафи, М., Зелано, К., Джонсон, Б., Материк, Дж., Кан Р. и Собел Н. (2004). Обоняние: от запаха к восприятию. В М. С. Газзанига (ред.), Когнитивные нейронауки (3-е изд.). Кембридж, Массачусетс: MIT Press.
Фельдман Р. (2007). Контакт матери и ребенка и развитие ребенка: выводы из вмешательства кенгуру. В L. L’Abate (Ed.), Недорогие подходы к укреплению физического и психического здоровья: теория, исследования и практика (стр. 323–351). Нью-Йорк, штат Нью-Йорк: Springer Science + Business Media.
Поле, Т., Ласко, Д., Манди, П., Хентелефф, Т., Кабат, С., Талпинс, С., и Доулинг, М. (1997). Краткий отчет: внимательность и отзывчивость аутичных детей улучшаются после сенсорной терапии. Журнал аутизма и нарушений развития, 27 (3), 333–338.
Харадон, Г., Баском, Б., Драгомир, К., и Скрипкару, В. (1994). Сенсорные функции институционализированных румынских младенцев: пилотное исследование. Международная ассоциация профессиональной терапии, 1 (4), 250–260.
Икеда, К.(1909/2002). [Новые приправы]. Chemical Senses, 27 (9), 847–849. Переведено и сокращено до 75% Я. Огивара и Я. Ниномия из журнала Токийского химического общества, 30 , 820–836. (Оригинальная работа опубликована в 1909 г.).
Келлинг, С. Т., и Халперн, Б. П. (1983). Вкусовые вспышки: время реакции, интенсивность и качество. Science, 219 , 412–414.
Келтнер, Д. (2009). Рожденный быть хорошим: наука о содержательной жизни. Нью-Йорк, Нью-Йорк: Нортон.
Малник Б., Хироно Дж., Сато Т. и Бак Л. Б. (1999). Комбинаторные рецепторные коды запахов. Cell, 96 , 713–723.
Мелзак Р. и Уолл П. (1996). Вызов боли. Лондон, Англия: Пингвин.
Мерфи, К. (1986). Вкус и запах у пожилых людей. В H. L. Meiselman & R. S. Rivlin (Eds.), Клиническое измерение вкуса и запаха (Vol. 1, pp. 343–371). Нью-Йорк, Нью-Йорк: Макмиллан.
Норткатт, Р.Г. (2004). Вкусовые рецепторы: развитие и эволюция. Мозг, поведение и эволюция, 64 (3), 198–206.
Штернберг, В. Ф., Бейлин, Д., Грант, М., и Грейсли, Р. Х. (1998). Соревнования меняют восприятие вредных раздражителей у спортсменов мужского и женского пола. Пейн, 76 (1–2), 231–238.
Сугимото, К., и Ниномия, Ю. (2005). Вступительные замечания по исследованиям умами: возможные рецепторы и механизмы передачи сигналов на умами. Chemical Senses, 30 (Дополнение.1), Pi21 – i22.
Уолл, П. (2000). Боль: наука о страдании . Нью-Йорк, Нью-Йорк: издательство Колумбийского университета.
Цвейер, К., Велкер, Б., и Рух, В. (2004). Умеряют ли жизнерадостность, возбуждение и юмор переносимость боли? Исследование FACS. Юмор: Международный журнал исследований юмора, 17 (1-2), 85–119.
Авторство изображений
Рисунок 5.21: По материалам Murphy (1986).
нервной системы человека | Описание, развитие, анатомия и функции
Пренатальное и постнатальное развитие нервной системы человека
Почти все нервные клетки или нейроны генерируются во время пренатальной жизни, и в большинстве случаев после этого они не заменяются новыми нейронами.Морфологически нервная система впервые появляется примерно через 18 дней после зачатия в результате образования нервной пластинки. Функционально он появляется с первым признаком рефлекторной активности во втором пренатальном месяце, когда стимуляция прикосновением к верхней губе вызывает реакцию отдергивания головы. Многие рефлексы головы, туловища и конечностей могут появиться на третьем месяце.
В процессе своего развития нервная система претерпевает значительные изменения, чтобы достичь своей сложной организации.Чтобы произвести примерно 1 триллион нейронов, присутствующих в зрелом мозге, в течение всей пренатальной жизни необходимо генерировать в среднем 2,5 миллиона нейронов в минуту. Это включает формирование нейронных цепей, содержащих 100 триллионов синапсов, поскольку каждый потенциальный нейрон в конечном итоге связан либо с выбранным набором других нейронов, либо с конкретными целями, такими как сенсорные окончания. Более того, синаптические связи с другими нейронами устанавливаются в определенных местах на клеточных мембранах целевых нейронов.Совокупность этих событий не считается исключительным продуктом генетического кода, поскольку генов просто не хватает, чтобы объяснить такую сложность. Скорее, дифференцировка и последующее развитие эмбриональных клеток в зрелые нейроны и глиальные клетки достигается двумя наборами влияний: (1) специфическими подмножествами генов и (2) стимулами окружающей среды внутри и вне эмбриона. Генетические влияния имеют решающее значение для развития нервной системы в упорядоченной и временной последовательности.Клеточная дифференцировка, например, зависит от серии сигналов, регулирующих транскрипцию, процесса, в котором молекулы дезоксирибонуклеиновой кислоты (ДНК) дают начало молекулам рибонуклеиновой кислоты (РНК), которые, в свою очередь, выражают генетические сообщения, контролирующие клеточную активность. Влияния окружающей среды, происходящие от самого эмбриона, включают клеточные сигналы, которые состоят из диффундирующих молекулярных факторов ( см. Ниже Развитие нейронов). К факторам внешней среды относятся питание, сенсорный опыт, социальное взаимодействие и даже обучение.Все это важно для правильной дифференциации отдельных нейронов и для тонкой настройки синаптических связей. Таким образом, нервная система требует постоянной стимуляции на протяжении всей жизни для поддержания функциональной активности.
Развитие нейронов
На второй неделе пренатальной жизни быстро растущая бластоциста (пучок клеток, на которые делится оплодотворенная яйцеклетка) превращается в так называемый эмбриональный диск. Эмбриональный диск вскоре приобретает три слоя: эктодерму (внешний слой), мезодерму (средний слой) и энтодерму (внутренний слой).Внутри мезодермы растет хорда, осевой стержень, который служит временным позвоночником. И мезодерма, и хорда выделяют химическое вещество, которое заставляет соседние недифференцированные клетки эктодермы утолщаться вдоль того, что станет дорсальной средней линией тела, образуя нервную пластинку. Нервная пластинка состоит из нервных клеток-предшественников, известных как нейроэпителиальные клетки, которые развиваются в нервную трубку ( см. Ниже Морфологическое развитие). Затем нейроэпителиальные клетки начинают делиться, диверсифицироваться и давать незрелые нейроны и нейроглию, которые, в свою очередь, мигрируют из нервной трубки в свое окончательное местоположение.Каждый нейрон образует дендриты и аксон; аксоны удлиняются и образуют ветви, концы которых образуют синаптические связи с выбранным набором целевых нейронов или мышечных волокон.
человеческое эмбриональное развитиеРазвитие человеческого эмбриона на 18-й день, на стадии диска или щита, показано на (слева) трехчетвертном виде и (справа) в поперечном сечении.
Британская энциклопедия, Inc. Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасЗамечательные события этого раннего развития включают упорядоченную миграцию миллиардов нейронов, рост их аксонов (многие из которых широко распространяются по всему мозгу) и формирование тысяч синапсов между отдельными аксонами и их целевыми нейронами.Миграция и рост нейронов зависят, по крайней мере частично, от химических и физических воздействий. Растущие концы аксонов (называемые конусами роста), по-видимому, распознают и реагируют на различные молекулярные сигналы, которые направляют аксоны и нервные ветви к их соответствующим целям и устраняют те, которые пытаются синапсировать с неподходящими целями. Как только синаптическая связь установлена, клетка-мишень высвобождает трофический фактор (например, фактор роста нервов), который необходим для выживания нейрона, синапсирующегося с ней.Сигналы физического наведения участвуют в наведении контактов или миграции незрелых нейронов по каркасу из глиальных волокон.
В некоторых регионах развивающейся нервной системы синаптические контакты изначально не точны или стабильны, а затем следует упорядоченная реорганизация, включая устранение многих клеток и синапсов. Нестабильность некоторых синаптических связей сохраняется до тех пор, пока не наступит так называемый критический период, до которого влияние окружающей среды играет значительную роль в правильной дифференцировке нейронов и в тонкой настройке многих синаптических связей.