Виды рецепторов
Существуют несколько классификаций рецепторов:
По адекватному раздражителю
Хеморецепторы — воспринимают воздействие растворенных или летучих химических веществ.
Осморецепторы — воспринимают изменения осмотической концентрации жидкости (как правило, внутренней среды).
Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.)
Фоторецепторы — воспринимают видимый и ультрафиолетовый свет
Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) температуры
Болевые рецепторы, стимуляция которых приводит к возникновению боли. Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно.

Электрорецепторы — воспринимают изменения электрического поля
Магнитные рецепторы — воспринимают изменения магнитного поля

У
человека имеются первые шесть типов
рецепторов. На хеморецепции основаны
вкус и обоняние, на механорецепции —
осязание, слух и равновесие, а также
ощущения положения тела в пространстве,
на фоторецепции — зрение. Терморецепторы
есть в коже и некоторых внутренних
органах. Большая часть интерорецепторов
запускает непроизвольные, и в большинстве
случаев неосознаваемые, вегетативные
рефлексы.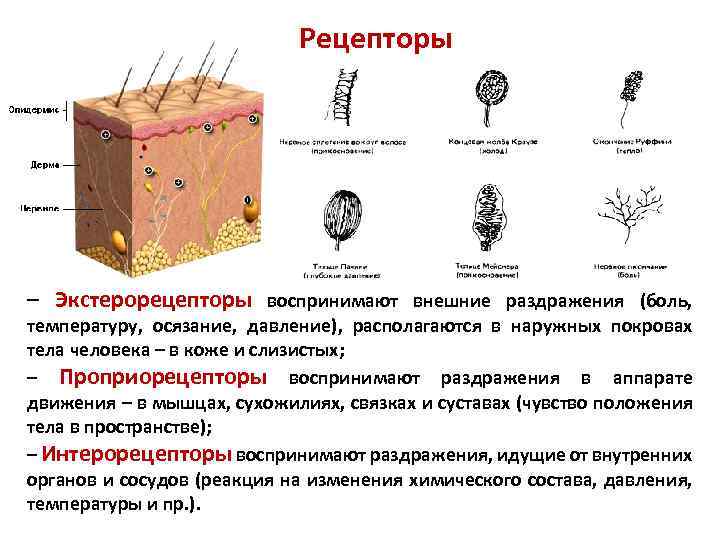
Иногда предлагается выделять группу электромагнитных рецепторов, в которую включают фото-, электро- и магниторецепторы. Магниторецепторы точно не идентифицированы ни у одной группы животных, хотя предположительно ими служат некоторые клетки сетчатки птиц, а возможно, и ряд других клеток[2].
26глаз (лат. oculus) — сенсорный орган (орган зрительной системы) человека и животных, обладающий способностью воспринимать электромагнитное излучениев световом диапазоне длин волн и обеспечивающий функцию
Глаз позвоночных
животных представляет
собой периферическую частьзрительного
анализатора,
в котором фоторецепторную функцию
выполняютнейроны —
фотосенсорные клетки («нейроциты») сетчатой
оболочки.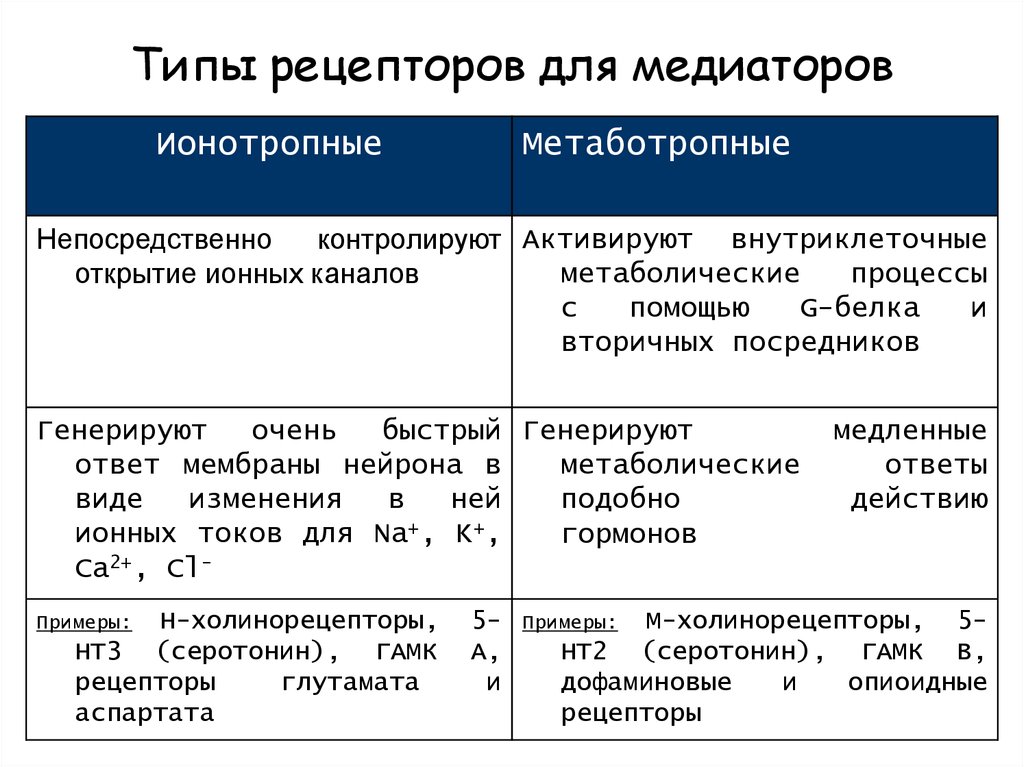 Внутреннее строение
Внутреннее строение
1. Задняя камера 2. Зубчатый край 3. Ресничная (аккомодационная) мышца 4. Ресничный (цилиарный) поясок 5. Шлеммов канал 6. Зрачок 7. Передняя камера 8. Роговица 9. Радужная оболочка 10. Кора хрусталика 11. Ядро хрусталика 12. Цилиарный отросток 13. Конъюнктива 14. Нижняя косая мышца 15. Нижняя прямая мышца 16. Медиальная прямая мышца
17. Артерии и вены сетчатки 18. Слепое пятно 19. Твердая мозговая оболочка 20. Центральная артерия сетчатки 21. Центральная вена сетчатки 22. Зрительный нерв 23. Вортикозная вена 24. Влагалище глазного яблока 25. Жёлтое пятно 26. Центральная ямка 27. Склера 28. Сосудистая оболочка глаза 29. Верхняя прямая мышца 30. Сетчатка Глазное
яблоко состоит из оболочек, которые
окружают внутреннее ядро глаза,
представляющее его прозрачное
содержимое — стекловидное
тело, хрусталик, водянистая
влага в
передней и задней камерах.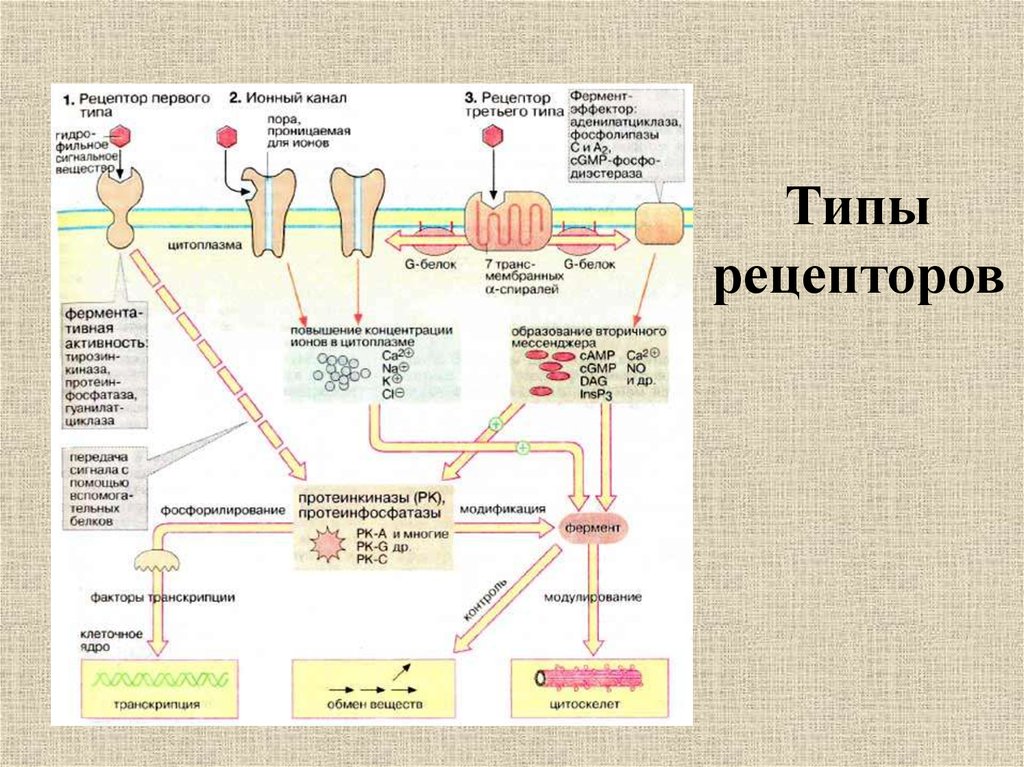
Ядро глазного яблока окружают три оболочки: наружная, средняя и внутренняя.
Наружная — очень плотная фиброзная оболочка глазного яблока (tunica fibrosa bulbi), к которой прикрепляются
Средняя, или сосудистая, оболочка глазного яблока (tunica vasculosa bulbi), играет важную роль в обменных процессах, обеспечивая питание глаза и выведение продуктов обмена. Она богата кровеносными сосудами и пигментом (богатые пигментом клетки хориоидеи препятствуют проникновению света через склеру, устраняя светорассеяние). Она образована радужкой, ресничным телом и собственно сосудистой оболочкой. В центре радужки имеется круглое отверстие — зрачок, через которое лучи света проникают внутрь глазного яблока и достигают сетчатки (величина зрачка изменяется в результате взаимодействия гладких мышечных волокон — сфинктера и дилататора, заключённых в радужке и иннервируемых
парасимпатическим и симпатическимнервами). Радужка содержит различное количество
пигмента, от которого зависит её
окраска — «цвет
глаз».Внутренняя, или сетчатая, оболочка глазного яблока (tunica interna bulbi), — сетчатка — рецепторная часть зрительного анализатора, здесь происходит непосредственное восприятие света, биохимические превращения зрительных пигментов, изменение электрических свойств нейронов и передача информации вцентральную нервную систему.
 Радужка содержит различное количество
пигмента, от которого зависит её
окраска — «цвет
глаз».
Радужка содержит различное количество
пигмента, от которого зависит её
окраска — «цвет
глаз».С функциональной точки зрения оболочки глаза и её производные подразделяют на три аппарата: рефракционный (светопреломляющий) и аккомодационный (приспособительный), формирующие оптическую систему глаза, и сенсорный (рецепторный) аппарат.
Рецепторы и их роль в организме человека
Рецепторы и их роль в организме человека
- Воробьев Антон Сергеевич
Рейтинг: 4 / 5
Пожалуйста, оцените Оценка 1Оценка 2Оценка 3Оценка 4Оценка 5Статья по анатомии и физиологии человека
Рецепторы и их роль в организме человекаВоробьев Антон Сергеевич
Рецептор
(от лат. recipere – получать) — чувствительное нервное окончание или специализированная клетка, преобразующее воспринимаемое раздражение в нервные импульсы.
Рецептор гораздо более восприимчив к внешним воздействиям, чем другие органы и нервные волокна. Чувствительность этого органа особенно высока и обратно пропорциональна порогу. То есть если говорят, что порог раздражения низкий, это значит, что чувствительность рецептора высокая.
Каждый рецептор предназначен для восприятия одного из видов раздражения.
Все рецепторы характеризуются наличием специфического участка мембраны, содержащего рецепторный белок, обусловливающий процессы рецепции.
Основной характеристикой рецепторного аппарата организма является его приспособленность к восприятию раздражений, повышенная чувствительность к ним и специализация к определенным видам воздействия.
Существуют несколькоклассификаций
рецепторов:
- По положению в организме
- Экстерорецепторы (экстероцепторы) — расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы из окружающей среды)
- Интерорецепторы (интероцепторы) — расположены во внутренних органах и воспринимают внутренние стимулы (например, информацию о состоянии внутренней среды организма)
- Проприорецепторы (проприоцепторы) — рецепторы опорно-двигательного аппарата, позволяющие определить, например, напряжение и степень растяжения мышц и сухожилий. Являются разновидностью интерорецепторов
- Проприорецепторы (проприоцепторы) — рецепторы опорно-двигательного аппарата, позволяющие определить, например, напряжение и степень растяжения мышц и сухожилий.
- По способности воспринимать разные стимулы
- Мономодальные — реагирующие только на один тип раздражителей (например, фоторецепторы — на свет)
- Полимодальные — реагирующие на несколько типов раздражителей (например, многие болевые рецепторы, а также некоторые рецепторы беспозвоночных, реагирующие одновременно на механические и химические стимулы)
- По адекватному раздражителю:
- Хеморецепторы — воспринимают воздействие растворенных или летучих химических веществ
- Осморецепторы — воспринимают изменения осмотической концентрации жидкости (как правило, внутренней среды)
- Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.)
- Фоторецепторы — воспринимают видимый и ультрафиолетовый свет
- Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) стимулы
- Болевые рецепторы, стимуляция которых приводит к возникновению боли. Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно. В действительности, они представляют собой высокопороговые сенсоры различных (химических, термических или механических) повреждающих факторов. Однако уникальная особенность ноцицепторов, которая не позволяет отнести их, например, к «высокопороговым терморецепторам», состоит в том, что многие из них полимодальны: одно и то же нервное окончание способно возбуждаться в ответ на несколько различных повреждающих стимулов.
- Электрорецепторы — воспринимают изменения электрического поля
- Магнитные рецепторы — воспринимают изменения магнитного поля
 Являются разновидностью интерорецепторов
Являются разновидностью интерорецепторов Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно. В действительности, они представляют собой высокопороговые сенсоры различных (химических, термических или механических) повреждающих факторов. Однако уникальная особенность ноцицепторов, которая не позволяет отнести их, например, к «высокопороговым терморецепторам», состоит в том, что многие из них полимодальны: одно и то же нервное окончание способно возбуждаться в ответ на несколько различных повреждающих стимулов.
Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно. В действительности, они представляют собой высокопороговые сенсоры различных (химических, термических или механических) повреждающих факторов. Однако уникальная особенность ноцицепторов, которая не позволяет отнести их, например, к «высокопороговым терморецепторам», состоит в том, что многие из них полимодальны: одно и то же нервное окончание способно возбуждаться в ответ на несколько различных повреждающих стимулов.
У человека имеются первые шесть типов рецепторов. На хеморецепции основаны вкус и обоняние, на механорецепции — осязание, слух и равновесие, а также ощущения положения тела в пространстве, на фоторецепции — зрение.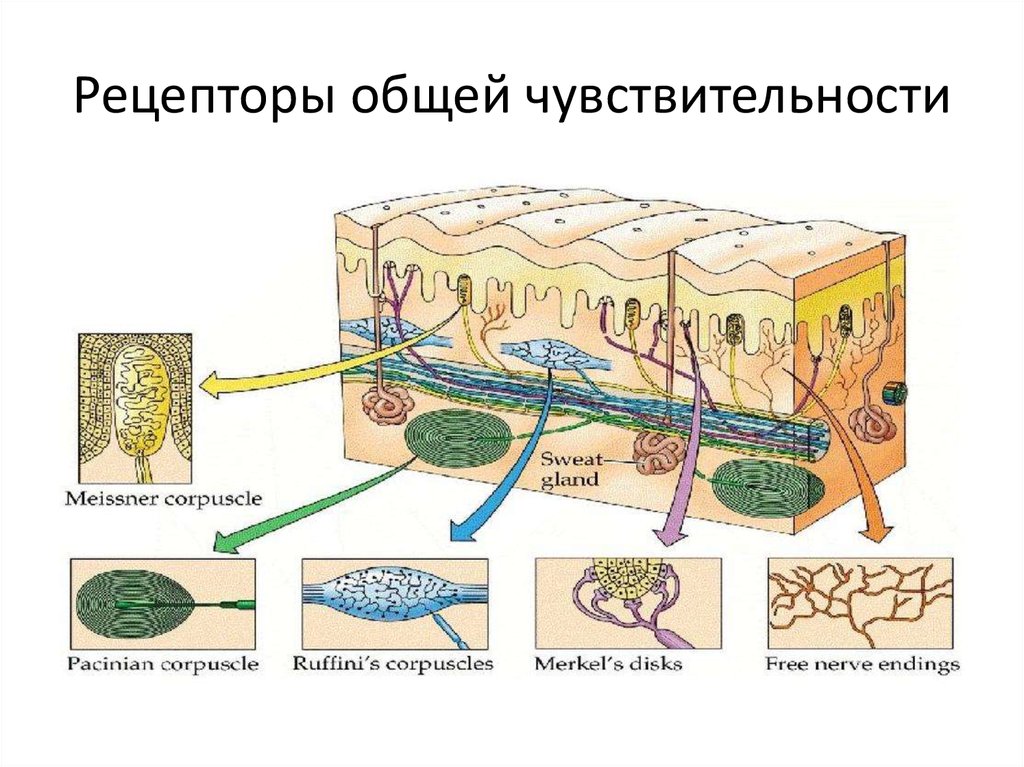 Терморецепторы есть в коже и некоторых внутренних органах. Большая часть интерорецепторов запускает непроизвольные, и в большинстве случаев неосознаваемые, вегетативные рефлексы. Так, осморецепторы включены в регуляцию деятельности почек, хеморецепторы, воспринимающие pH, концентрации углекислого газа и кислорода в крови, включены в регуляцию дыхания и т. д.
Терморецепторы есть в коже и некоторых внутренних органах. Большая часть интерорецепторов запускает непроизвольные, и в большинстве случаев неосознаваемые, вегетативные рефлексы. Так, осморецепторы включены в регуляцию деятельности почек, хеморецепторы, воспринимающие pH, концентрации углекислого газа и кислорода в крови, включены в регуляцию дыхания и т. д.
Иногда предлагается выделять группу электромагнитных рецепторов, в которую включают фото-, электро- и магниторецепторы. Магниторецепторы точно не идентифицированы ни у одной группы животных, хотя предположительно ими служат некоторые клетки сетчатки птиц, а возможно, и ряд других клеток.
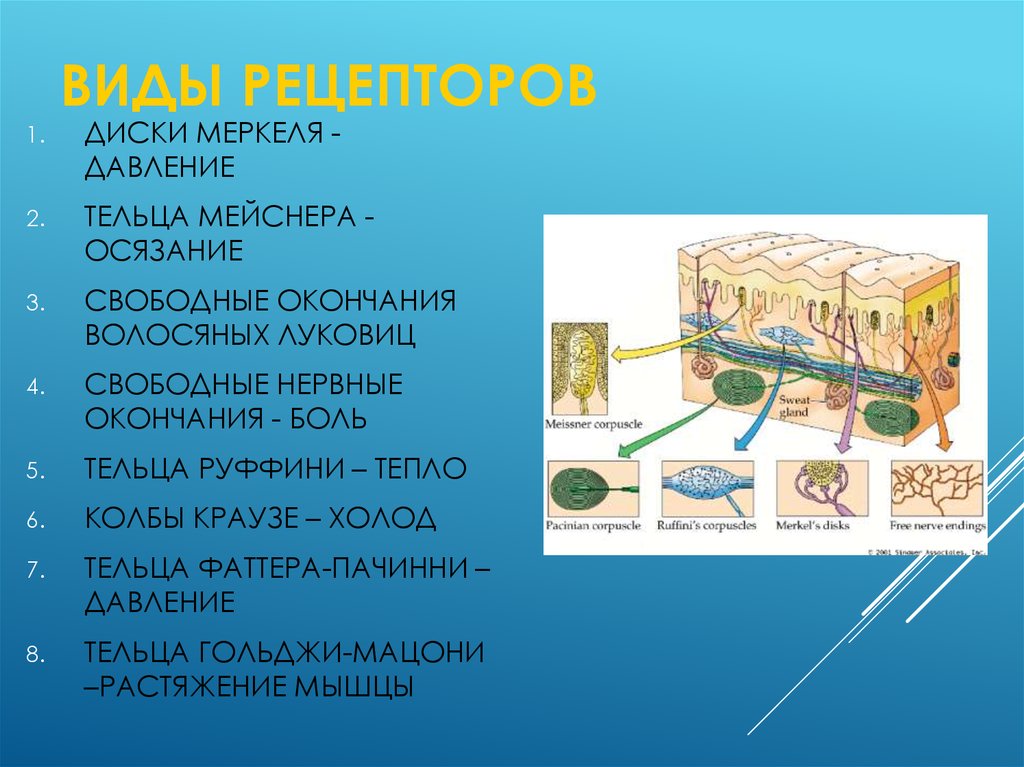
Рецепторы кожи
- Болевые рецепторы.
- Тельца Пачини — капсулированные рецепторы давления в округлой многослойной капсуле. Располагаются вподкожно-жировой клетчатке. Являются быстроадаптирующимися (реагируют только в момент началавоздействия), то есть регистрируют силу давления. Обладают большими рецептивными полями, то естьпредставляют грубую чувствительность.
- Тельца Мейснера — рецепторы давления, расположенные в дерме. Представляют собой слоистую структурус нервным окончанием, проходящим между слоями. Являются быстроадаптирующимися. Обладают малымирецептивными полями, то есть представляют тонкую чувствительность.
- Диски Меркеля — некапсулированные рецепторы давления. Являются медленноадаптирующимися (реагируют на всей продолжительности воздействия), то есть регистрируют продолжительность давления. Обладают малыми рецептивными полями.
- Рецепторы волосяных луковиц — реагируют на отклонение волоса.
- Окончания Руффини — рецепторы растяжения. Являются медленноадаптирующимися, обладают большимирецептивными полями.
Рецепторы мышц и сухожилий
- Мышечные веретена — рецепторы растяжения мышц, бывают двух типов:
- с ядерной сумкой
- с ядерной цепочкой
- Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилиерастягивается и его волокна пережимают рецепторное окончание, активируя его.
 При сокращении мышцы сухожилиерастягивается и его волокна пережимают рецепторное окончание, активируя его.
При сокращении мышцы сухожилиерастягивается и его волокна пережимают рецепторное окончание, активируя его.Рецепторы связок
В основном представляют собой свободные нервные окончания (Типы 1, 3 и 4), меньшая группа — инкапсулированные (Тип 2). Тип 1 аналогичен окончаниям Руффини, Тип 2 — тельцам Паччини.
Рецепторы сетчатки глаза
Сетчатка содержит палочковые (палочки) и колбочковые (колбочки) фоточувствительные клетки, которыесодержат светочуствительные пигменты. Палочки чуствительны к очень слабому свету, это длинные и тонкиеклетки, сориентированные по оси прохождения света. Все палочки содержат один и тот же светочуствительный пигмент. Колбочки требуют намного более яркого освещения, это короткиеконусообразные клетки, у человека колбочки делятся на три вида, каждый из которых содержит свойсветочуствительный пигмент — это и есть основа цветового зрения.
Под воздействием света в рецепторах происходит выцветание — молекула зрительного пигмента поглощает фотон и превращается в другое соединение, хуже поглощающее свет волн (этой длины волны). Практическиу всех животных (от насекомых до человека) этот пигмент состоит из белка, к которому присоединенанебольшая молекула, близкая к витамину A. Эта молекула и представляет собой химическитрансформируемую светом часть. Белковая часть выцвевшей молекулы зрительного пигмента активируетмолекулы трансдуцина, каждая из которых деактивирует сотни молекул циклического гуанозинмонофосфата, участвующих в открытии пор мембраны для ионов натрия, в результате чего поток ионов прекращается — мембрана гиперполяризуется.
Практическиу всех животных (от насекомых до человека) этот пигмент состоит из белка, к которому присоединенанебольшая молекула, близкая к витамину A. Эта молекула и представляет собой химическитрансформируемую светом часть. Белковая часть выцвевшей молекулы зрительного пигмента активируетмолекулы трансдуцина, каждая из которых деактивирует сотни молекул циклического гуанозинмонофосфата, участвующих в открытии пор мембраны для ионов натрия, в результате чего поток ионов прекращается — мембрана гиперполяризуется.
Чуствительность палочек такова, что адаптировавшийся к полной темноте человек способен увидеть вспышкусвета такую слабую, что ни один рецептор не может получить больше одного фотона. При этом палочки неспособны реагировать на изменения освещённости, когда свет настолько ярок, что все натриевые поры ужезакрыты.
Литература:
- Дэвид Хьюбел — «Глаз, мозг, зрение» перевод с англ. канд. биол. наук О. В. Левашова, канд. биол. наук Г. А. Шараева под ред. чл.-корр. АН СССР А. Л. Бызова, Москва «Мир», 1990
- http://anatomus.ru/articles/rol-retseptorov.html
 чл.-корр. АН СССР А. Л. Бызова, Москва «Мир», 1990
чл.-корр. АН СССР А. Л. Бызова, Москва «Мир», 1990
- Назад
- Вперед
You have no rights to post comments
Авторизация
Запомнить меня
Регистрация
- Забыли данные входа?
- Регистрация
Перевод сайта
Подробности и регистрация со СКИДКОЙ
типов рецепторов – принципы биологии
Клетка внутри многоклеточного организма может нуждаться в передаче сигналов другим клеткам, которые находятся на разных расстояниях от исходной клетки ( Рисунок 1 ). Не на все клетки воздействуют одни и те же сигналы.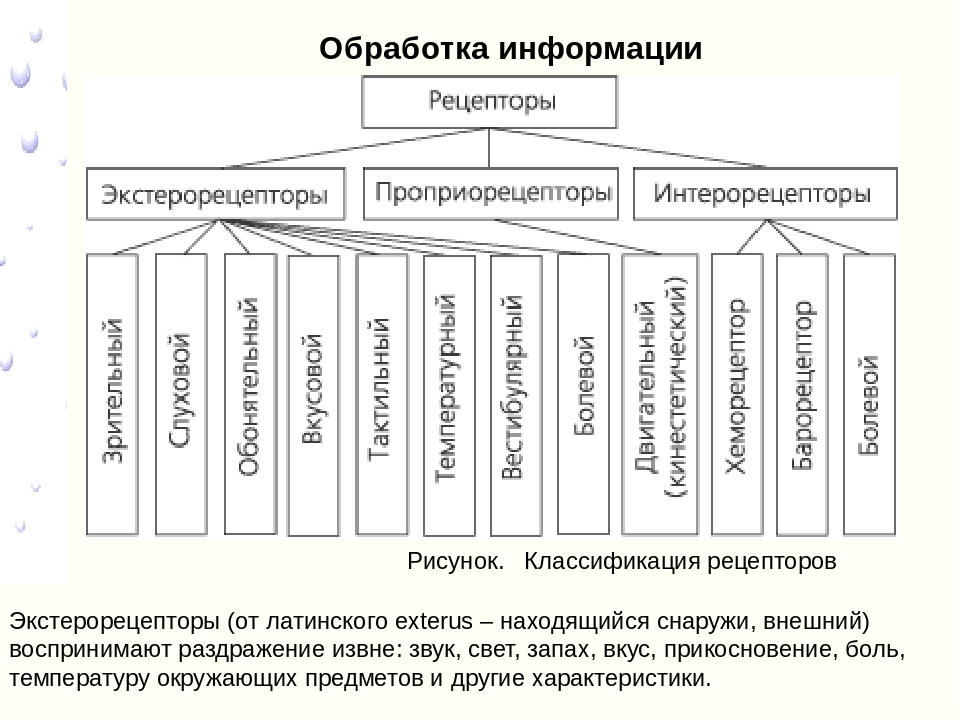 Различные типы сигнализации используются для разных целей.
Различные типы сигнализации используются для разных целей.
 Таким образом, лиганды можно рассматривать как сигнальные молекулы. Лиганды и рецепторы существуют в нескольких вариантах; однако специфический лиганд будет иметь специфический рецептор, который обычно связывает только этот лиганд.
Таким образом, лиганды можно рассматривать как сигнальные молекулы. Лиганды и рецепторы существуют в нескольких вариантах; однако специфический лиганд будет иметь специфический рецептор, который обычно связывает только этот лиганд.Внутренние рецепторы
Внутренние рецепторы , также известные как внутриклеточные или цитоплазматические рецепторы, находятся в цитоплазме клетки и реагируют на молекулы гидрофобного лиганда, способные проходить через плазматическую мембрану. Оказавшись внутри клетки, многие из этих молекул связываются с белками, которые действуют как регуляторы синтеза мРНК. Напомним, что мРНК переносит генетическую информацию от ДНК в ядре клетки к рибосоме, где собирается белок. Когда лиганд связывается с внутренним рецептором, запускается изменение формы, которое обнажает сайт связывания ДНК на белке-рецепторе. Комплекс лиганд-рецептор перемещается в ядро, затем связывается с определенными участками ДНК и способствует продукции мРНК из определенных генов (9). 0003 Рисунок 2 ). Внутренние рецепторы могут напрямую влиять на экспрессию генов (сколько определенного белка вырабатывается из гена) без необходимости передавать сигнал другим рецепторам или мессенджерам.
0003 Рисунок 2 ). Внутренние рецепторы могут напрямую влиять на экспрессию генов (сколько определенного белка вырабатывается из гена) без необходимости передавать сигнал другим рецепторам или мессенджерам.
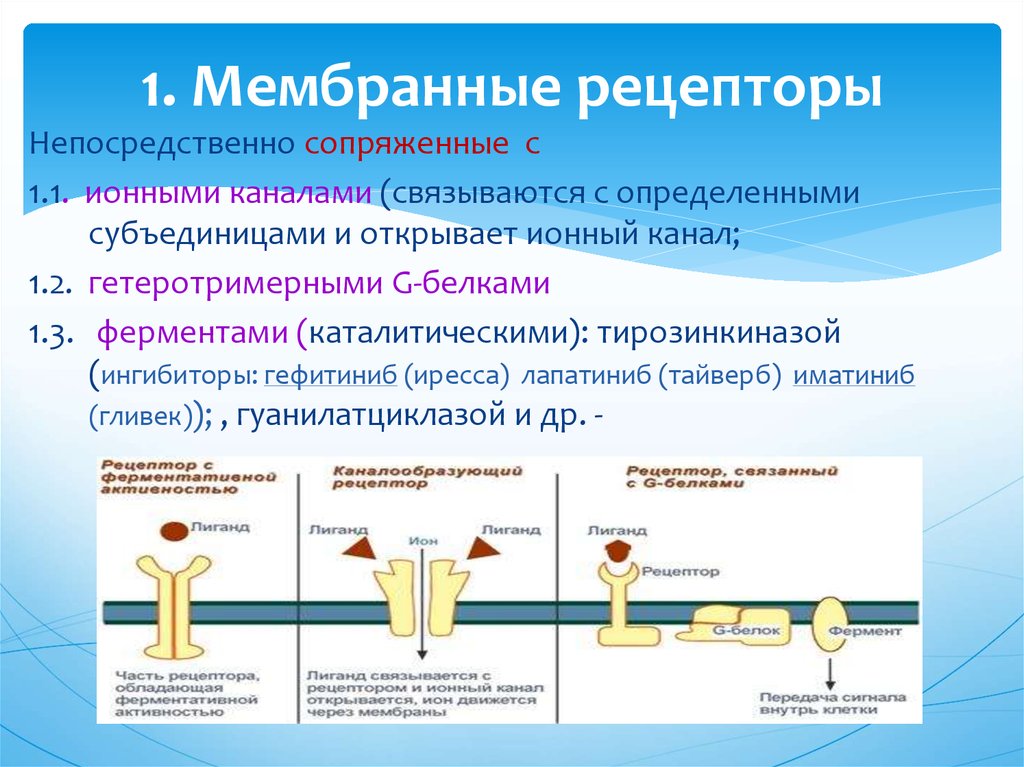
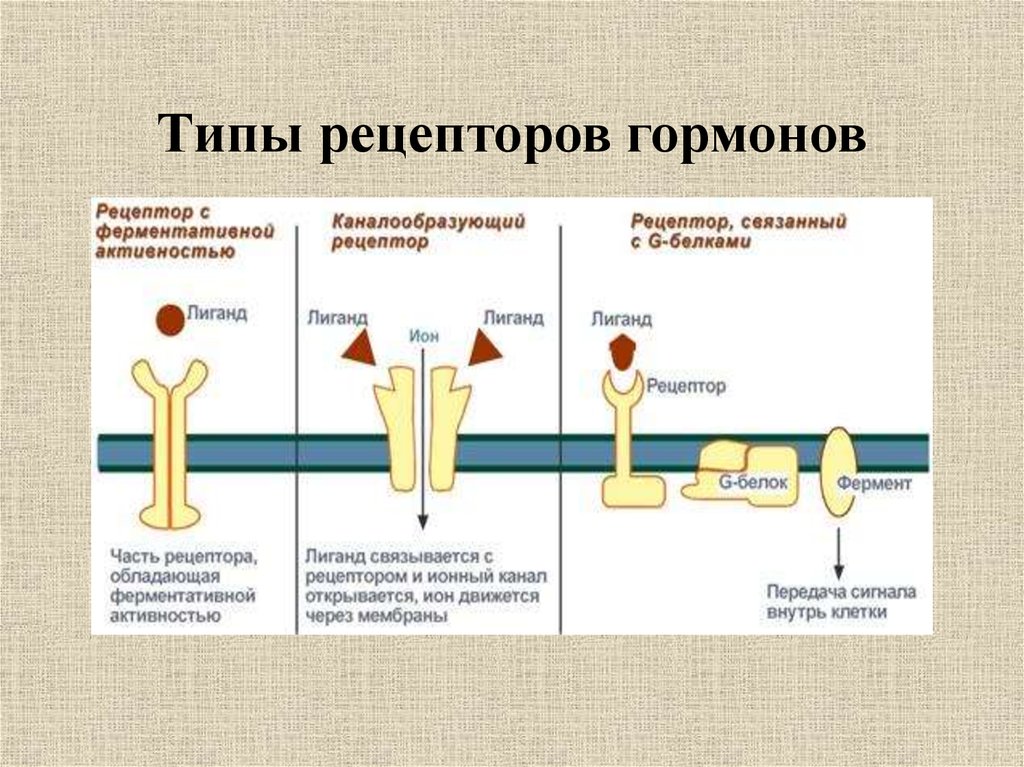
Рецепторы клеточной поверхности
Рецепторы клеточной поверхности, также известные как трансмембранные рецепторы , представляют собой белки, прикрепленные к клеточной мембране. Эти рецепторы связываются с молекулами внешнего лиганда (лиганды, которые не проходят через клеточную мембрану). Этот тип рецепторов охватывает плазматическую мембрану и выполняет передачу сигнала , при которой внеклеточный сигнал преобразуется в межклеточный сигнал. Лиганды, которые взаимодействуют с рецепторами клеточной поверхности, не должны проникать в клетку, на которую они воздействуют. Рецепторы клеточной поверхности также называют клеточно-специфическими белками или маркерами, поскольку они специфичны для отдельных типов клеток.
Лиганды, которые взаимодействуют с рецепторами клеточной поверхности, не должны проникать в клетку, на которую они воздействуют. Рецепторы клеточной поверхности также называют клеточно-специфическими белками или маркерами, поскольку они специфичны для отдельных типов клеток.
Каждый рецептор клеточной поверхности состоит из трех основных компонентов: внешнего лиганд-связывающего домена, гидрофобной трансмембранной области и внутриклеточного домена внутри клетки. Размер и протяженность каждого из этих доменов сильно различаются в зависимости от типа рецептора.
Рисунок 3 Рецепторы клеточной поверхности функционируют путем передачи сигнала через клеточную мембрану. Лиганд не попадает непосредственно в клетку. Фото предоставлено Laozhengzz; Достояние Викимедиа. Рецепторы клеточной поверхности вовлечены в большую часть передачи сигналов в многоклеточных организмах. Существует три основных категории рецепторов клеточной поверхности: рецепторы, связанные с ионными каналами, рецепторы, связанные с G-белком, и рецепторы, связанные с ферментами.
Рецепторы, связанные с ионным каналом
Рецепторы, связанные с ионным каналом , связывают лиганд и открывают канал через мембрану, который позволяет проходить определенным ионам. Чтобы сформировать канал, этот тип рецептора клеточной поверхности имеет обширную область, охватывающую мембрану. Когда лиганд связывается с внеклеточной областью канала, происходит конформационное изменение в структуре белка, что позволяет проходить через него таким ионам, как натрий, кальций, магний и водород ( рис. 4) .
Рис. 4 Закрытые ионные каналы образуют поры в плазматической мембране, которые открываются при связывании сигнальной молекулы. Затем открытая пора позволяет ионам втекать в клетку или выходить из нее.Рецепторы, связанные с G-белком
Рецепторы, связанные с G-белком связывают лиганд и активируют мембранный белок, называемый G-белком. Затем активированный G-белок взаимодействует либо с ионным каналом, либо с ферментом в мембране (, рис.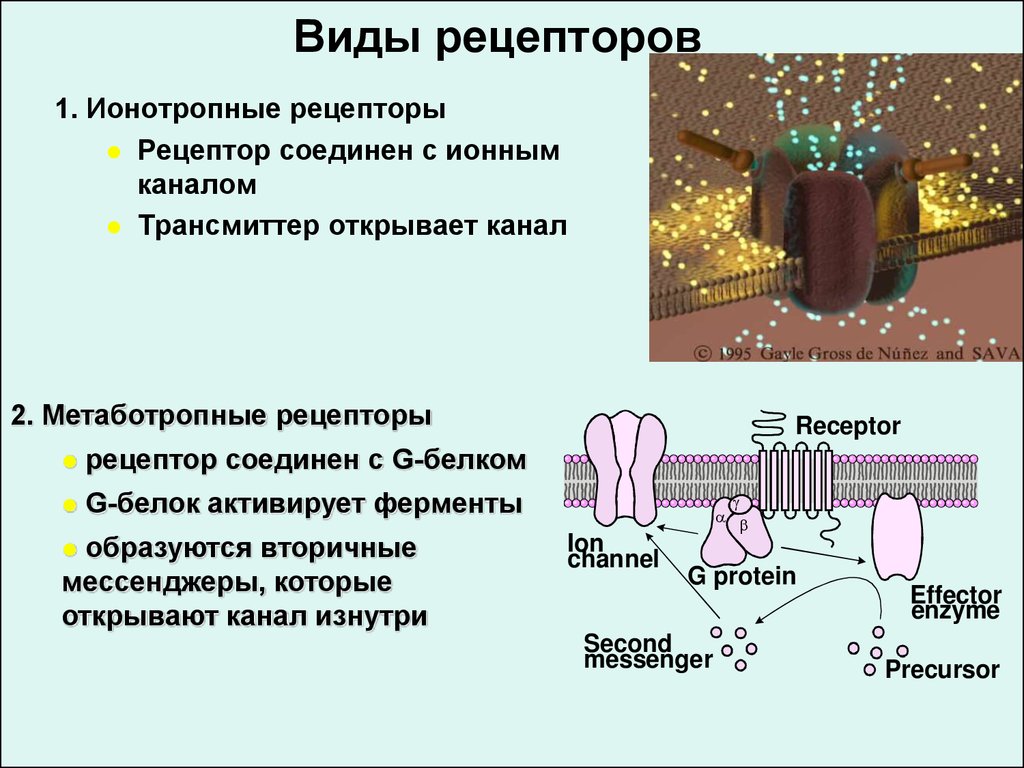 5, ). Перед связыванием лиганда неактивный G-белок может связываться с участком на конкретном рецепторе. Как только G-белок связывается с рецептором, G-белок меняет форму, становится активным и разделяется на две разные субъединицы. В результате одна или обе эти субъединицы могут активировать другие белки.
5, ). Перед связыванием лиганда неактивный G-белок может связываться с участком на конкретном рецепторе. Как только G-белок связывается с рецептором, G-белок меняет форму, становится активным и разделяется на две разные субъединицы. В результате одна или обе эти субъединицы могут активировать другие белки.
Связанные с ферментом рецепторы
Связанные с ферментом рецепторы представляют собой рецепторы клеточной поверхности с внутриклеточными доменами, которые связаны с ферментом. В некоторых случаях внутриклеточный домен самого рецептора представляет собой фермент. Другие рецепторы, связанные с ферментами, имеют небольшой внутриклеточный домен, непосредственно взаимодействующий с ферментом. Когда лиганд связывается с внеклеточным доменом, сигнал передается через мембрану, активируя фермент. Активация фермента запускает цепочку событий внутри клетки, которая в конечном итоге приводит к ответной реакции.
Когда лиганд связывается с внеклеточным доменом, сигнал передается через мембрану, активируя фермент. Активация фермента запускает цепочку событий внутри клетки, которая в конечном итоге приводит к ответной реакции.
Как вирусы узнают хозяина
В отличие от живых клеток многие вирусы не имеют плазматической мембраны или каких-либо структур, необходимых для поддержания жизни. Некоторые вирусы просто состоят из инертной белковой оболочки, содержащей ДНК или РНК. Для размножения вирусы должны проникнуть в живую клетку, которая служит хозяином, а затем захватить клеточный аппарат хозяина. Но как вирус узнает своего хозяина?
Вирусы часто связываются с рецепторами клеточной поверхности на клетке-хозяине . Например, вирус, вызывающий человеческий грипп (грипп), специфически связывается с рецепторами на мембранах клеток дыхательной системы. Химические различия в рецепторах клеточной поверхности у хозяев означают, что вирус, поражающий определенный вид (например, человека), не может заразить другой вид (например, цыплят).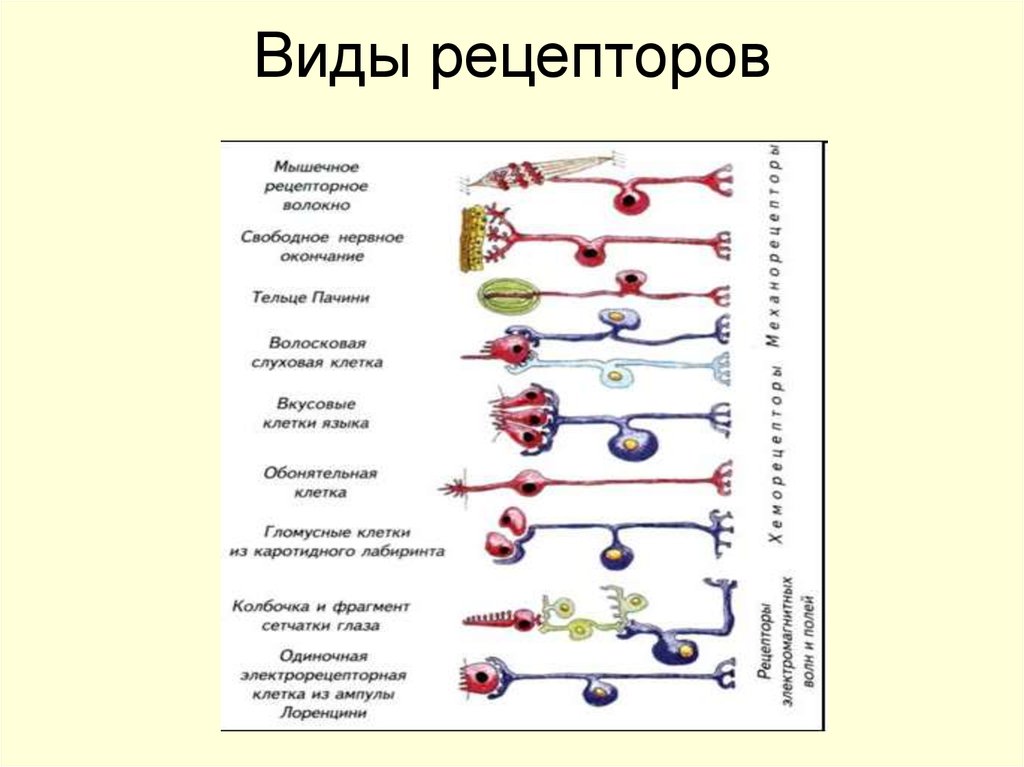
Однако вирусы имеют очень небольшое количество ДНК или РНК по сравнению с людьми, и в результате вирусное размножение может происходить быстро. Вирусное размножение неизменно приводит к ошибкам, которые могут привести к изменениям во вновь созданных вирусах; эти изменения означают, что вирусные белки, которые взаимодействуют с рецепторами клеточной поверхности, могут развиваться таким образом, что они могут связываться с рецепторами нового хозяина. Такие изменения происходят случайно и довольно часто в репродуктивном цикле вируса, но изменения имеют значение только в том случае, если вирус с новыми связывающими свойствами вступает в контакт с подходящим хозяином. В случае гриппа такая ситуация может возникнуть в условиях тесного контакта животных и людей, например, на птицефабриках и свиноводческих фермах (Сигалов, 2010). Как только вирус попадает на новый хост, он может быстро распространиться. Ученые внимательно следят за вновь появляющимися вирусами (так называемые новые вирусы) в надежде, что такой мониторинг может снизить вероятность глобальных вирусных эпидемий.
Ссылки
Текст адаптирован из: OpenStax, Biology. OpenStax CNX. 13 октября 2017 г. https://cnx.org/contents/GFy_h8cu@10.118:h5oMpCSi@8/Signaling-Molecules-and-Cellul#footnote1
.Сигалов А.Б. Школа естествознания. IV. Учимся у вирусов, Свой/Не свой 1, вып. 4 (2010): 282-298. Ю. Цао, X. Кох, Л. Донг, X. Ду, А. Ву, X. Дин, Х. Дэн, Ю. Шу, Дж. Чен, Т. Цзян, Быстрая оценка активности связывания гемагглютинина вируса гриппа с Рецепторы человека и птиц, PLoS One 6, нет. 4 (2011): e18664.
Типы рецепторов — Неврология — Книжная полка NCBI
Независимо от природы инициирующего сигнала клеточные ответы определяются наличием рецепторов, которые специфически связываются с сигнальными молекулами. Связывание сигнальных молекул вызывает конформационные изменения в рецепторе, которые затем запускают последующий сигнальный каскад. Учитывая, что химические сигналы могут действовать либо на плазматической мембране, либо внутри цитоплазмы (или ядра) клетки-мишени, неудивительно, что рецепторы обнаруживаются с обеих сторон плазматической мембраны.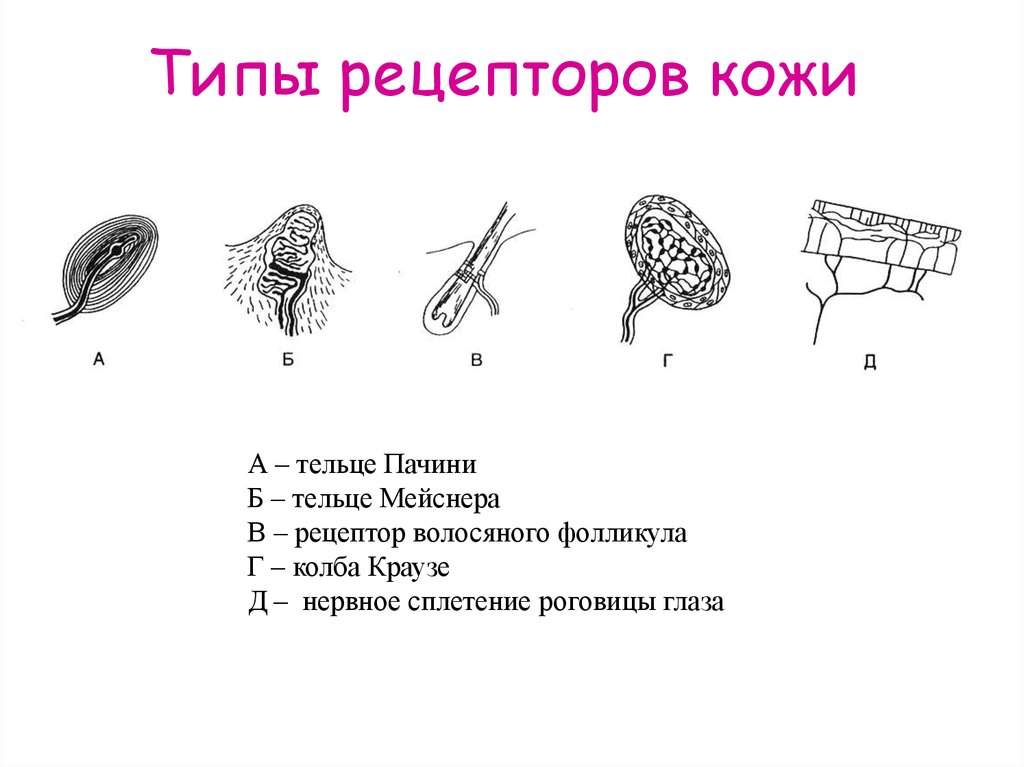
Рецепторы для непроникающих сигнальных молекул представляют собой трансмембранные белки, которые включают компоненты как снаружи, так и внутри клеточной поверхности. Внеклеточный домен таких рецепторов включает сайт связывания сигнала, в то время как внутриклеточный домен активирует внутриклеточные сигнальные каскады после связывания сигнала. Было идентифицировано большое количество этих рецепторов, которые сгруппированы в три семейства, определяемые механизмом, используемым для преобразования связывания сигнала в клеточный ответ (-).
Рисунок 8.4
Категории клеточных рецепторов. Мембранно-непроницаемые сигнальные молекулы могут связываться и активировать рецепторы, связанные с каналом (А), рецепторы, связанные с ферментами (В), или рецепторы, связанные с G-белком (С). Проникающие в мембрану сигнальные молекулы активируют внутриклеточные (подробнее…)
Связанные с каналами рецепторы (также называемые лиганд-управляемыми ионными каналами) выполняют функции рецептора и трансдукции как часть одной и той же белковой молекулы.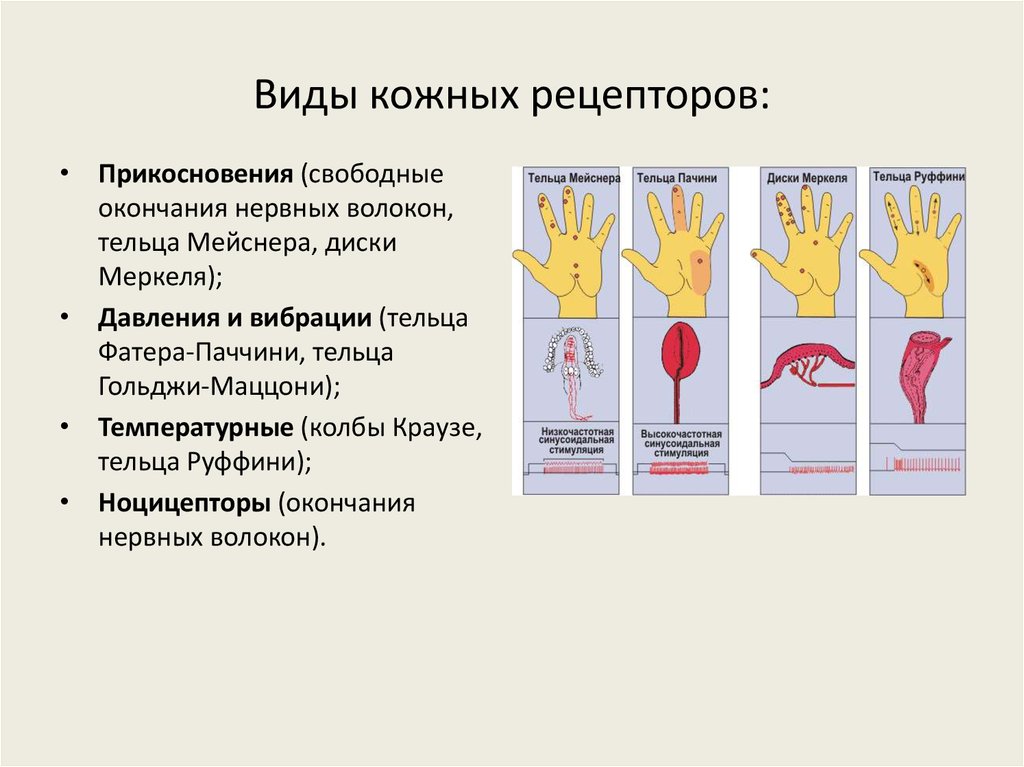 Взаимодействие химического сигнала с местом связывания рецептора вызывает открытие или закрытие поры ионного канала в другой части той же молекулы. Возникающий ионный поток изменяет мембранный потенциал клетки-мишени и в некоторых случаях также может приводить к проникновению Ca 2+ ионов, которые служат вторичным мессенджером сигнала внутри клетки. Хорошими примерами таких рецепторов являются рецепторы нейротрансмиттеров, описанные в главе 7.
Взаимодействие химического сигнала с местом связывания рецептора вызывает открытие или закрытие поры ионного канала в другой части той же молекулы. Возникающий ионный поток изменяет мембранный потенциал клетки-мишени и в некоторых случаях также может приводить к проникновению Ca 2+ ионов, которые служат вторичным мессенджером сигнала внутри клетки. Хорошими примерами таких рецепторов являются рецепторы нейротрансмиттеров, описанные в главе 7.
Связанные с ферментом рецепторы также имеют внеклеточный сайт связывания химических сигналов. Внутриклеточный домен таких рецепторов представляет собой фермент, каталитическая активность которого регулируется связыванием внеклеточного сигнала. Подавляющее большинство этих рецепторов представляют собой протеинкиназ, часто тирозинкиназ , , которые фосфорилируют внутриклеточные белки-мишени, тем самым изменяя физиологическую функцию клеток-мишеней. Заслуживающие внимания члены этой группы рецепторов включают семейство Trk рецепторов нейротрофинов (см. главу 23) и другие рецепторы факторов роста.
главу 23) и другие рецепторы факторов роста.
Связанные с G-белком рецепторы регулируют внутриклеточные реакции посредством косвенного механизма, включающего промежуточную трансдуцирующую молекулу, называемую GTP-связывающими белками (или G-белками). Поскольку все эти рецепторы обладают общей структурной особенностью семикратного пересечения плазматической мембраны, их также называют 7-трансмембранными рецепторами (или метаботропными рецепторами; см. главу 7). Были идентифицированы сотни различных рецепторов, связанных с G-белком. Хорошо известные примеры включают β-адренергический рецептор, мускарининовый тип ацетилхолинового рецептора, метаботропные глутаматные рецепторы, рецепторы одорантов в обонятельной системе и многие типы рецепторов пептидных гормонов. Родопсин, светочувствительный 7-трансмембранный белок в фоторецепторах сетчатки, представляет собой другую форму рецептора, связанного с G-белком (см. главу 11).
Внутриклеточные рецепторы активируются проникающими в клетку или липофильными сигнальными молекулами ().