22. Роль генотипа и среды в формировании фенотипа, в повышении продуктивности сельскохозяйственных растений и животных.
1. Фенотип — совокупность внешних и внутренних признаков, особенности функционирования организма. Генотип — совокупность генов, которые организм получает от родителей.
2. Зависимость проявления генотипа, влияния генов на формирование фенотипа от условий среды. Модификационная изменчивость — изменение фенотипа, не связанное с изменением генотипа. Пример: разрезанную вдоль одну половину корня одуванчика выращивали в горах, а другую на равнине. В горах из нее выросло растение с мелкими листьями, низкое, а на равнине высокое, с крупными листьями. Причины различий — влияние условий среды (при одинаковом генотипе).
3.
Пределы модификационной изменчивости
— норма реакции. Широкая норма реакции:
значительные изменения признака,
например, надоев молока в зависимости
от кормления, ухода; узкая норма реакции,
незначительные изменения признака,
например, жирности молока, окраски
шерсти. Изменения фенотипа, вызванные
изменениями окружающей среды, не ведут
к изменению генотипа.
Изменения фенотипа, вызванные
изменениями окружающей среды, не ведут
к изменению генотипа.
4. Наследование нормы реакции организмом, причина изменения нормы реакции — изменение генотипа. Формирование фенотипа — результат взаимодействия генотипа с условиями среды.
5. Приспособительное значение модификационной изменчивости для сохранения и процветания вида.
6. Применение знаний о модификационной изменчивости в сельском хозяйстве. Пример: плодородная почва, хороший уход для реализации генотипа высокопродуктивных сортов растений. Проявление признаков пород крупного рогатого скота, свиней, овец только при соблюдении рациона кормления, правил ухода за животными. Нарушение научной технологии выращивания растений и животных — причина снижения их продуктивности.
23. Разнообразие сортов растений и пород животных — результат селекционной работы ученых. Закон н. И. Вавилова о гомологических рядах в наследственной изменчивости.
1.
Селекция — наука о выведении новых
сортов растений и пород животных. Порода
(сорт) — искусственно созданная человеком
популяция, которая характеризуется
наследственными биологическими
особенностями, морфологическими и
физиологическими признаками,
продуктивностью. 2. Ч. Дарвин —
основоположник науки селекции,
обосновавший значение наследственной
изменчивости и искусственного отбора
в создании новых сортов и пород.
Порода
(сорт) — искусственно созданная человеком
популяция, которая характеризуется
наследственными биологическими
особенностями, морфологическими и
физиологическими признаками,
продуктивностью. 2. Ч. Дарвин —
основоположник науки селекции,
обосновавший значение наследственной
изменчивости и искусственного отбора
в создании новых сортов и пород.
3. Вклад Н. И. Вавилова в развитие науки селекции, в разработку ее задач. Обоснование Н. И. Вавиловым необходимости использования законов генетики в качестве научных основ селекции. Изучение и создание им коллекции сортового и видового разнообразия растений как исходного материала для селекции.
4. Закон Н. И. Вавилова о гомологических рядах в наследственной изменчивости, его значение для селекции: выявление сходных наследственных изменений у организмов близких видов.
5.
Изучение Н. И. Вавиловым видового
разнообразия. Богатство генофонда диких
видов, превышение генофонда сортов
растений и пород животных, необходимость
изучения мирового богатства видов для
селекции.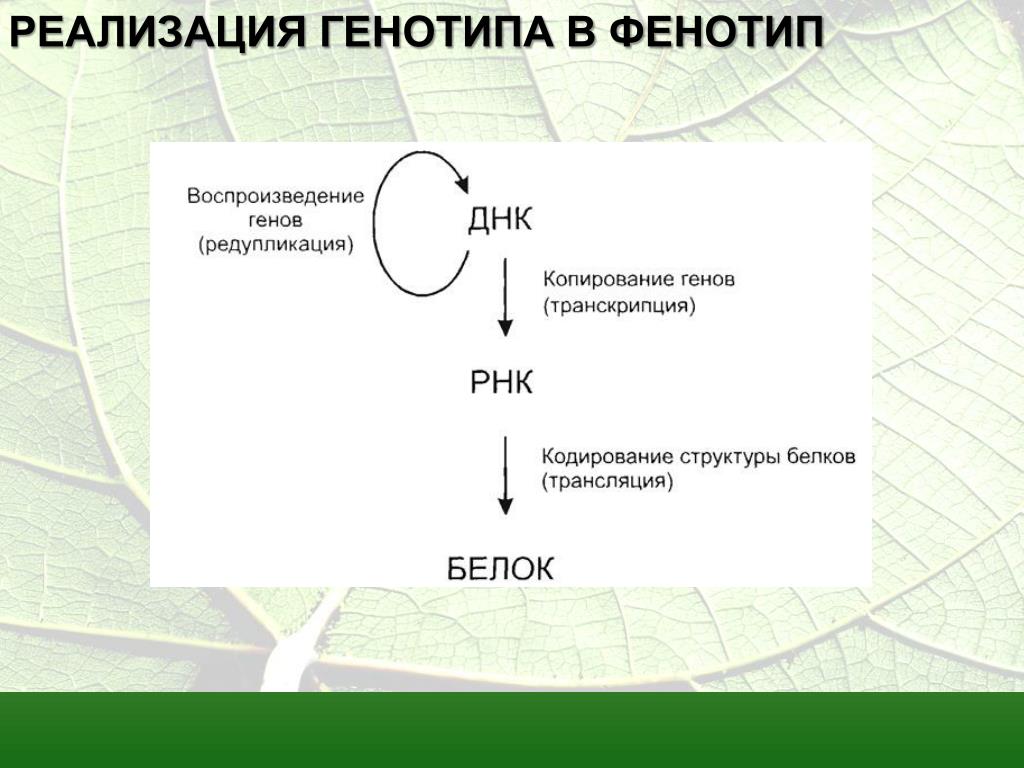
6. Учение Н. И. Вавилова о центрах многообразия и происхождения культурных растений. Центры происхождения культурных растений — в основном горные районы, древние очаги земледелия, характеризующиеся многообразием видов, разновидностей, родина сортов растений. Основные центры происхождения культурных растений.
7. Значение селекции — создание большого разнообразия высокопродуктивных сортов растений, полиплоидных форм, пригодных для выращивания в разных климатических условиях, а также пород животных, высокопродуктивных гибридных форм, бройлеров и др.
Роль генотипа и условий внешней среды в формировании фенотипа
Все многообразие всего живого и его постоянное
совершенствование были бы невозможны без изменчивости. Это
связано с тем, что генотип последовательно реализуется в
фенотип в ходе индивидуального развития организма и в
определенных условиях среды обитания, факторы которой
(колебания освещенности, температуры, влажности, условий
питания, взаимоотношений с другими организмами и др.
Если растения примулы, которые в обычных условиях имели красные цветки, перенести в оранжерею и содержать их там при температуре 30—35°С и высокой влажности, то через некоторое время все цветки у них оказываются белыми. Если эти же растения вновь поместить в обычные (комнатные) условия, то распустившиеся цветки будут красными.
Другим классическим примером, иллюстрирующим влияние внешней
среды на проявление качественных признаков, служит изменение
окраски шерсти у гималайского кролика. Обычно при 20°С у этого
кролика шерсть белая, за исключением черных ушей, лап, хвоста
и мордочки. При 30°С такие кролики вырастают полностью белыми.
Если же у гималайского кролика сбрить шерсть на боку или спине
и содержать его при температуре воздуха ниже +2°С, то вместо
белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то
в обычных условиях там снова вырастет черная шерсть.
При 30°С такие кролики вырастают полностью белыми.
Если же у гималайского кролика сбрить шерсть на боку или спине
и содержать его при температуре воздуха ниже +2°С, то вместо
белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то
в обычных условиях там снова вырастет черная шерсть.
Эти наблюдения объясняют, почему гималайские кролики рождаются белыми, без участков черной шерсти: их эмбриональное развитие происходит в условиях высокой температуры.
Известно, что все признаки и свойства организма наследственно детерминированы, однако организмы наследуют не сами признаки и свойства, а лишь возможность их развития. Для проявления и развития признака необходимы соответствующие условия внешней среды.Но даже в том случае, когда проявление и развитие того или иного признака происходит, степень его выраженности бывает
разной в зависимости от условий внешней среды: при одних она
усиливается, при других ослабляется. Пределы этих изменений
определены возможностями, заложенными в генотипе.
Пределы этих изменений
определены возможностями, заложенными в генотипе.
Пределы модификационной изменчивости признака, ограниченные действием генотипа, называют его норной реакции.
Для разных признаков и свойств организмов границы, определяемые нормой реакции, неодинаковы. Наибольшей пластичностью и изменчивостью характеризуются количественные признаки: семенная продуктивность злаков, величина удоя у крупного рогатого скота, масса животных, число и размеры листьев и колосков у растений и т. д. В то же время качественные признаки (масть животных, окраска семян, цветков и плодов, остистость и опушенность колоса у растений) мало зависят от условий среды.
Так, у крупного рогатого скота удой во многом определяется
условиями его содержания. При подборе кормов нужного качества
и количества ценная порода скота может дать 5—6 тыс. кг молока
в год, а в случае ухудшения условий содержания продуктивность
снижается до 2,5 тыс. кг и даже ниже. Что касается масти
животных, то при самых разных условиях она почти не
изменяется.
кг молока
в год, а в случае ухудшения условий содержания продуктивность
снижается до 2,5 тыс. кг и даже ниже. Что касается масти
животных, то при самых разных условиях она почти не
изменяется.
Подобную закономерность можно наблюдать и у растений. Например, растения элитного сорта картофеля, выращенные в условиях высокой культуры агротехники, дают урожай клубней 500—600 ц/га и более. Ухудшение условий (тяжелые глинистые почвы, низкая культура земледелия и ухода за растениями) может привести к тому, что урожайность прекрасного сорта картофеля снизится до 100—150 ц/га и ниже. Правда, окраска клубней и их форма при этом изменяются в очень малой степени.
Рассмотренные примеры свидетельствуют о том, что
количественные признаки обладают очень широкой нормой реакции,
а качественные — более узкой.
Таким образом, все признаки и свойства каждого сорта растений и каждой породы животных могут изменяться только в пределах нормы реакции. Поэтому попытки добиться высоких урожаев, резко повысить продуктивность сельскохозяйственных животных только за счет повышения уровня культуры земледелия, улучшения условий кормления и содержания животных несостоятельны. В первую очередь необходимо получить новые сорта растений и породы животных улучшенного генотипа, отзывчивые на хорошие условия возделывания и выращивания.
По механизму возникновения и характеру изменений признаков различают два типа изменчивости — наследственную и ненаследственную.
Под наследственной изменчивостью понимают способность к изменениям самого генетического материала, а под ненаследственной (модификационной, или фенотипической)
— способность организмов реагировать на условия окружающей среды и изменяться в пределах нормы реакции, определяемой генотипом.
Отличие генотипа-фенотипа Вильгельма Йоханнсена | Энциклопедия Embryo Project Encyclopedia
Различие генотипа и фенотипа Вильгельма Йохансена Вильгельм Йоханнсен впервые предложил различие между генотипом и фенотипом при изучении наследственности, работая в Дании в 1909 году. Различие проводится между наследственной предрасположенностью организмов (их генотипами) и способами в которых эти склонности проявляются в физических характеристиках этих организмов (их фенотипах). Это различение явилось следствием экспериментов Йоханнсена, касающихся наследуемой изменчивости растений, и повлияло на его чисто линейную теорию наследственности. Хотя значение и значение различия генотип-фенотип было предметом споров — среди современников Йоханнсена, более поздних биологических теоретиков и историков науки — многие считают это различие одним из концептуальных столпов генетики двадцатого века. Более того, некоторые использовали его для характеристики отношений между исследованиями развития, генетики и эволюции.
Йоханнсен ввел понятия генотип и фенотип в 1909 году в своем учебнике по исследованию наследственности под названием Elemente der exakten Ereblichkeitslehre (Элементы точной теории наследственности), и он разработал их более полно в статье 1911 года под названием «Генотипическая концепция наследственности». Понятия генотипа и фенотипа возникли в результате чистолинейных экспериментов Йохансена по разведению ячменя. ( Hordeum vulgare ) и фасоль обыкновенная ( Phaseolus vulgaris ).
Эксперименты Йохансена с чистыми линиями начались примерно в то время, когда он принял должность лектора ботаники и физиологии растений в Королевском ветеринарном и сельскохозяйственном колледже в Копенгагене, Дания, в 1892 году. В этих экспериментах Йохансен выращивал ячмень и бобовые растения, которые имели были самоопылены, и он измерял физические размеры семян в каждом поколении. Он обнаружил, что может разделить эти чистые линии на отдельные группы на основе характеристик семян, которые они производят.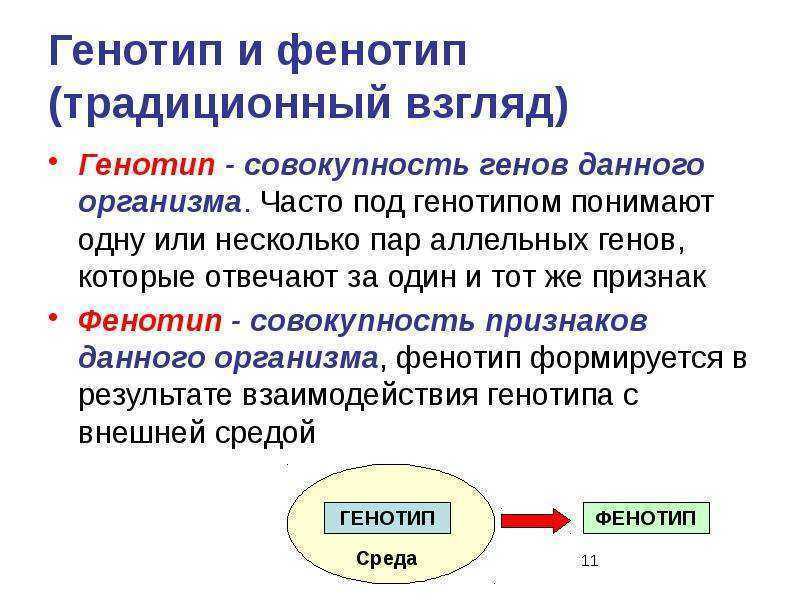 Используя комбинацию генеалогического и статистического анализа, Йоханнсен продемонстрировал, что группа, к которой принадлежало растение, была более сильным предсказателем характеристик семян, которые оно произвело, чем характеристики его материнского растения. Когда он впервые сообщил о результатах своих экспериментов в 1903, Йоханнсен назвал эту групповую идентичность ее типом, но в 1905 году он переименовал ее в термин генотип. Он противопоставлял генотип группы или организма их фенотипу, определяя последний как индивидуальные качества этих организмов.
Используя комбинацию генеалогического и статистического анализа, Йоханнсен продемонстрировал, что группа, к которой принадлежало растение, была более сильным предсказателем характеристик семян, которые оно произвело, чем характеристики его материнского растения. Когда он впервые сообщил о результатах своих экспериментов в 1903, Йоханнсен назвал эту групповую идентичность ее типом, но в 1905 году он переименовал ее в термин генотип. Он противопоставлял генотип группы или организма их фенотипу, определяя последний как индивидуальные качества этих организмов.
Эти эксперименты проводились в рамках более широкой попытки исследователей наследственности охарактеризовать виды и характер изменчивости организмов на рубеже двадцатого века. В то время биологи расходились во мнениях относительно того, как понимать взаимосвязь между изменчивостью, наследственностью и эволюционными изменениями. Последователи Дарвина утверждали, что эволюция произошла в результате действия естественного отбора на непрерывную наследственную изменчивость. Другие утверждали, что непрерывная изменчивость редко бывает наследственной или что даже если она и существует, такая изменчивость имеет пределы, так что она не может быть основой для долгосрочных эволюционных изменений. Сторонники последней позиции, воодушевленные так называемым переоткрытием теории Грегора Менделя о наследственности, утверждали, что эволюция должна происходить прерывистыми скачками. Для Йохансена различие между генотипом и фенотипом способствовало последней позиции: он считал генотип особенно неизменным, а силу естественного отбора ограничивал сортировкой ранее существовавших генотипов в гетерогенных природных популяциях.
Другие утверждали, что непрерывная изменчивость редко бывает наследственной или что даже если она и существует, такая изменчивость имеет пределы, так что она не может быть основой для долгосрочных эволюционных изменений. Сторонники последней позиции, воодушевленные так называемым переоткрытием теории Грегора Менделя о наследственности, утверждали, что эволюция должна происходить прерывистыми скачками. Для Йохансена различие между генотипом и фенотипом способствовало последней позиции: он считал генотип особенно неизменным, а силу естественного отбора ограничивал сортировкой ранее существовавших генотипов в гетерогенных природных популяциях.
Различие между генотипом и фенотипом было частью кампании Йохансена против того, что он называл концепцией передачи наследственности, согласно которой характеристики отдельных организмов передаются непосредственно их потомству. Примеры трансмиссионной концепции наследственности включают теорию пангенезиса Чарльза Дарвина, в которой изменяющиеся ткани и органы организма постоянно модифицируют его зародышевый материал, тем самым передавая характеристики этого организма его возможному потомству. Концепция генотипа Йоханнсена, напротив, была внеисторической: фенотипические характеристики матери и ее потомства возникали под влиянием одной и той же наследственной предрасположенности, передаваемой из поколения в поколение и невосприимчивой к условиям окружающей среды, в которых она проявлялась. Различие между генотипом и фенотипом Йоханнсена имеет некоторое сходство с различием Августа Вейсмана в конце девятнадцатого века между зародышем и сомой, поскольку оба считали, что причинные взаимодействия между наследственной предрасположенностью организма и его физическими характеристиками были однонаправленными. Хотя Йохансен признавал эту близость к идеям Вейсмана, он не желал участвовать в том, что он считал необоснованными рассуждениями о материальной основе генотипа, как, по его утверждению, сделал Вейсман.
Концепция генотипа Йоханнсена, напротив, была внеисторической: фенотипические характеристики матери и ее потомства возникали под влиянием одной и той же наследственной предрасположенности, передаваемой из поколения в поколение и невосприимчивой к условиям окружающей среды, в которых она проявлялась. Различие между генотипом и фенотипом Йоханнсена имеет некоторое сходство с различием Августа Вейсмана в конце девятнадцатого века между зародышем и сомой, поскольку оба считали, что причинные взаимодействия между наследственной предрасположенностью организма и его физическими характеристиками были однонаправленными. Хотя Йохансен признавал эту близость к идеям Вейсмана, он не желал участвовать в том, что он считал необоснованными рассуждениями о материальной основе генотипа, как, по его утверждению, сделал Вейсман.
Различие между генотипом и фенотипом, проведенное Иогансеном, явно запрещает связь между изучением развития и изучением наследственности. По Иогансену, генотип организма порождает фенотип организма в процессе развития под влиянием среды.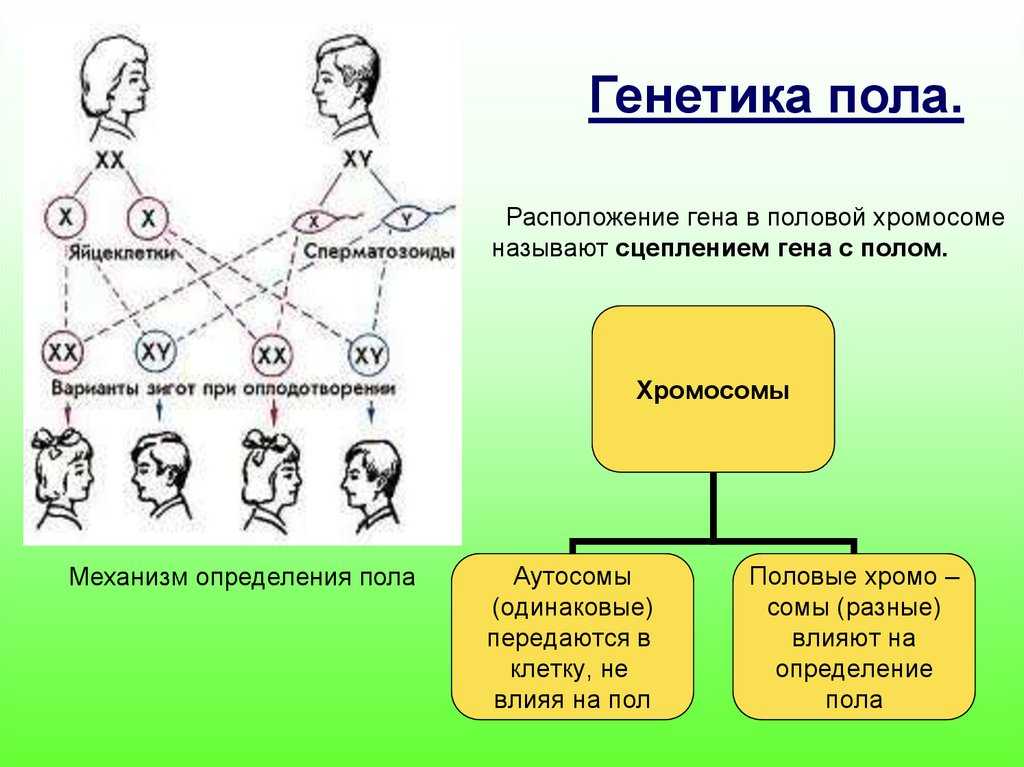 Йоханнсен чувствовал себя комфортно, отождествляя генотип с понятием Reaktionsnorm (норма реакции), предложенная Рихардом Вольтереком в Германии в 1909 году, которая для области различных возможных сред, в которых может развиваться организм, относилась к полному диапазону потенциальных вариаций во взрослом состоянии, характерных для этого организма. В соответствии со схемой различия генотипа и фенотипа биология развития была истолкована как изучение того, как генотипы порождают фенотипы. Такая интерпретация противоречила взглядам многих эмбриологов того времени, которые рассматривали наследственность как процесс или производство, механизмы которого неотделимы от механизмов самого развития.
Йоханнсен чувствовал себя комфортно, отождествляя генотип с понятием Reaktionsnorm (норма реакции), предложенная Рихардом Вольтереком в Германии в 1909 году, которая для области различных возможных сред, в которых может развиваться организм, относилась к полному диапазону потенциальных вариаций во взрослом состоянии, характерных для этого организма. В соответствии со схемой различия генотипа и фенотипа биология развития была истолкована как изучение того, как генотипы порождают фенотипы. Такая интерпретация противоречила взглядам многих эмбриологов того времени, которые рассматривали наследственность как процесс или производство, механизмы которого неотделимы от механизмов самого развития.
Правильное применение терминов генотип и фенотип было предметом некоторых споров после их введения. Зоолог Герберт Спенсер Дженнингс из США, например, интерпретировал понятия генотипа и фенотипа как непротиворечивые, утверждая в письме 1911 года в журнал Science , что, хотя абстрактный термин генотип относится к конкретной наследственной конституции организма , фенотип более конкретно относился к группе людей, имеющих одинаковые физические характеристики. Джордж Шулл, также находящийся в США, якобы консультировался с Йоханнсеном по этому вопросу, и Шулл ответил Дженнингсу в 1919 году.12. Шулл утверждал, что эти термины были в равной степени абстрактными и противоположными, причем фенотип относился не к самой группе физически сходных особей, а к общим характеристикам, которые были основой для разграничения группы. Отношения между генотипом и фенотипом — с точки зрения как его механистического, так и концептуального содержания — были постоянным предметом споров в эволюционной биологии.
Джордж Шулл, также находящийся в США, якобы консультировался с Йоханнсеном по этому вопросу, и Шулл ответил Дженнингсу в 1919 году.12. Шулл утверждал, что эти термины были в равной степени абстрактными и противоположными, причем фенотип относился не к самой группе физически сходных особей, а к общим характеристикам, которые были основой для разграничения группы. Отношения между генотипом и фенотипом — с точки зрения как его механистического, так и концептуального содержания — были постоянным предметом споров в эволюционной биологии.
По мере того как споры о роли непрерывной и прерывистой изменчивости в эволюции продолжались, а экспериментальные и теоретические достижения привели биологов к единому мнению вокруг понятия гена, язык генотипов и фенотипов получил широкое признание. Историк науки Ян Сапп утверждал, что различие между генотипом и фенотипом послужило отчуждению эмбриологов от основных исследований наследственности, создав пропасть между биологией развития и тем, что впоследствии станет популяционно-генетическим объяснением эволюционных изменений, лежащим в основе современного эволюционного синтеза. Более поздние дебаты о единицах отбора и о связи между микроэволюционными и макроэволюционными процессами в основном велись в терминах различия генотипа и фенотипа. Некоторые теоретики и философы охарактеризовали современную область эволюционной биологии развития как заполнение так называемого черного ящика между генотипом и фенотипом.
Более поздние дебаты о единицах отбора и о связи между микроэволюционными и макроэволюционными процессами в основном велись в терминах различия генотипа и фенотипа. Некоторые теоретики и философы охарактеризовали современную область эволюционной биологии развития как заполнение так называемого черного ящика между генотипом и фенотипом.
- Аллен, Гарланд Э. «Наследственность в эмбриологической парадигме: пример генетики и эмбриологии». Биологический бюллетень 168 (1985): 107–21.
- Черчилль, Фередерик Б. «Уильям Йоханнсен и концепция генотипа». Журнал истории биологии 7 (1974): 5–30.
- Данн, Лорен С. «Йоханнсен, Вильгельм Людвиг». Полный научно-биографический словарь. 7: 113–5.
- Холл, Брайан К. «Открытие черного ящика между генотипом и фенотипом: конденсация клеток как морфогенетические (модульные) единицы». Биология и философия 18 (2003): 219–47.
- Дженнингс, Герберт Спенсер. ««Генотип» и «Чистая линия».
 Наука 34 (1911): 841–2.
Наука 34 (1911): 841–2. - Йоханнсен, Вильгельм Лугвиг. «Ом Фровиден ог денс Удвиклинг хос Быг». «Об эндосперме и его развитии у ячменя». Allgemeine Zeitschrift fur Bierbrauerei und Malzfabrik 12 (1884): 625–750.
- Йоханнсен, Вильгельм Людвиг. Über Erblichkeit в Populationen und Reinen Linien. Eine Beitrag zur Beleuchtung schwebender Selektionsfragen [О наследственности в чистых линиях и популяциях. Вклад в нерешенные вопросы выбора]. Йена: Густав Фишер, 1903.
- Йоханнсен, Вильгельм Людвиг. Elemente der Exakten Erblichkeitslehre [Элементы точной теории наследственности]. Йена: Густав Фишер, 1909 г. http://dx.doi.org/10.5962/bhl.title.1060 (по состоянию на 7 декабря 2012 г.).
- Йоханнсен, Вильгельм Людвиг. «Генотипическая концепция наследственности». Американский натуралист 45 (1911): 129–59.
- Йоханнсен, Вильгельм Людвиг. Falske Analogier med henblikk paa Lighed, Slaegtskab, Arv, Tradition og Udvikling [Ложные аналогии в отношении сходства, родства, традиции и развития]. Копенгаген: Дж. Х. Шульц, 1914. .
- Ким, Кёнман. «О восприятии теории чистой линии Йохансена: к социологии научной достоверности». Социальные исследования науки 21 (1991): 649–79.
- Провайн, Уильям Б. Истоки теоретической популяционной генетики . Чикаго: Издательство Чикагского университета, 2001. .
- Райнбергер, Ханс-Йорг. «Наследственность и ее сущности около 1900 года». Исследования по истории и философии науки, часть A 39 (2008): 370–4.
- Ролл-Хансен, Нильс. «Решающий эксперимент Вильгельма Йохансена». Биология и философия 4 (1989): 303–29.
- Ролл-Хансен, Нильс. «Источники теории генотипа Вильгельма Йоханнсена». Журнал истории биологии 42 (2009): 457–93.
- Сапп, Январь. «Борьба за авторитет в области наследственности, 1900–1932: новые взгляды на рост генетики». Журнал истории биологии 16 (1983): 311–42.
- Шулл, Джордж Харрисон. 1912. «Фенотип и клон». Наука 35 (1912): 182–3. http://www.biodiversitylibrary.org/item/61279#page/349/mode/1up (по состоянию на 7 декабря 2012 г.).
- Сноррасон, Эрлинг Б. «Хансен, Эмиль Кристиан». Полный научно-биографический словарь. 6: 99–101.
- Ваншер, Йохан Хенрик. «Анализ генетического термина «генотип» Вильгельма Йохансена 1909–26». Hereditas 79 (1975): 1–4.
- Юл, Джордж. Удный. «Эксперименты профессора Йохансена в области наследственности: обзор». Новый фитолог 2 (1903): 235–42.
 Наука 34 (1911): 841–2.
Наука 34 (1911): 841–2.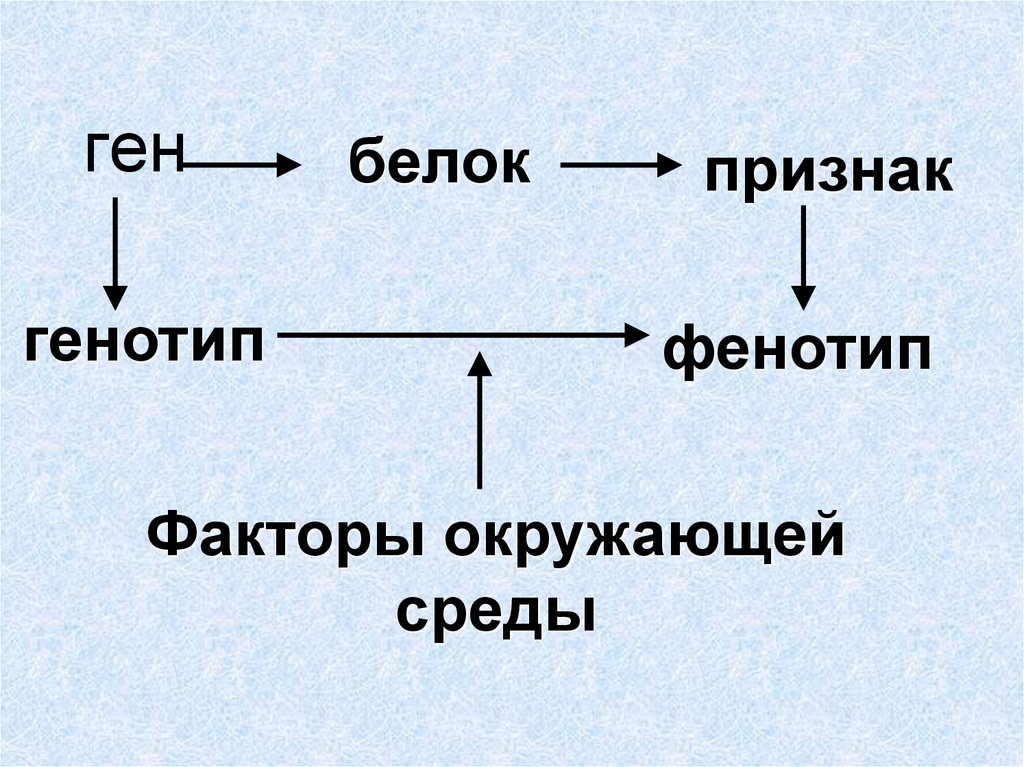 Копенгаген: Дж. Х. Шульц, 1914.
Копенгаген: Дж. Х. Шульц, 1914. http://www.biodiversitylibrary.org/item/61279#page/349/mode/1up (по состоянию на 7 декабря 2012 г.).
http://www.biodiversitylibrary.org/item/61279#page/349/mode/1up (по состоянию на 7 декабря 2012 г.).Пирсон, Б. Р. Эрик, «Различие генотипа и фенотипа Вильгельма Йоханнсена». Энциклопедия проекта «Эмбрио» (07 декабря 2012 г.). ISSN: 1940-5030 http://embryo.asu.edu/handle/10776/4206.
Университет штата Аризона. Школа наук о жизни. Центр биологии и общества. Энциклопедия проекта «Эмбрион».
Авторские права Попечительского совета штата Аризона Лицензия Creative Commons Attribution-NonCommercial-Share Alike 3. 0 Unported (http://creativecommons.org/licenses/by-nc-sa/3.0/)
0 Unported (http://creativecommons.org/licenses/by-nc-sa/3.0/)
Генетические явления; Генотип; Йоханнсен, В. (Вильгельм), 1857–1819 гг.27; Взаимодействие генотип-среда; Фенотип; Наследственность; генетика; Ботаника; физиология растений; Эволюция; Естественный отбор; Мендель, Грегор, 1822–1884 гг.; эмбриология; Дженнингс, Г. С. (Герберт Спенсер), 1868–1947; Шулл, Джордж Харрисон, 1874–1954 гг.; Концепция
Исследование под руководством UCI проливает свет на загадочные ассоциации генотип-фенотип | Медицинский факультет
Новые данные вносят свой вклад в знания о молекулярных механизмах, лежащих в основе многих сложных человеческих признаков и болезней
Ирвин, Калифорния – 1 июня 2021 г. — Новое исследование, анализирующее связь между генетикой человека (генотипом) и его наблюдаемыми характеристиками в результате взаимодействия генетики и окружающей среды (фенотип), вносит новые знания в понимание сложные человеческие качества и болезни.
Исследование под названием «Атлас локусов количественных признаков альтернативного полиаденилирования (3’aQTL), влияющих на наследуемость сложных признаков и заболеваний», было недавно опубликовано в Природа Генетика. Это новое исследование, проведенное под руководством профессора биоинформатики Калифорнийского университета в Ирвине Вей Ли, доктора философии, заведующей кафедрой биологической химии Медицинской школы UCI Грейс Б. Белл, раскрывает, насколько различия в генах людей объясняют различия в их чертах. и что можно отнести к воздействию окружающей среды.
«Наше исследование имеет особое значение, поскольку оно предлагает интерпретации, объясняющие, как естественные вариации могут формировать фенотипическое разнообразие человека и тканеспецифические заболевания», — сказал Ли. «Наше самое интересное открытие заключалось в том, что определенные события в генах человека могут объяснить значительную часть наследуемости признаков».
Хорошо известно, что 3′-UTR альтернативное полиаденилирование (APA) встречается примерно в 70 процентах генов человека и существенно влияет на клеточные процессы, такие как пролиферация, дифференцировка и онкогенез. Но до сих пор связь событий APA с риском заболевания и сложными человеческими чертами была недостаточно изучена.
Но до сих пор связь событий APA с риском заболевания и сложными человеческими чертами была недостаточно изучена.
Исследования ассоциации всего генома выявили тысячи некодирующих вариантов, связанных с человеческими чертами и заболеваниями. Однако функциональная интерпретация этих вариантов оказалась серьезной проблемой. В этом исследовании исследователи построили мультитканевой атлас локусов количественных признаков альтернативного полиаденилирования (APA) 3’UTR человека (3’aQTL), содержащий приблизительно 0,4 миллиона общих генетических вариантов, связанных с APA генов-мишеней, идентифицированных в 46 выделенных тканях. от 467 человек (Проект экспрессии генотипа ткани).
«Исходя из наших результатов, мы можем показать, что специфические молекулярные особенности, связанные с фенотипическими вариациями человека, вносят существенный вклад в молекулярные механизмы, лежащие в основе сложных черт и болезней человека», — пояснил Ли.
Исследовательская группа продолжает изучать эти молекулярные механизмы, чтобы протестировать новые гены 3’aQTL для диабета, рака предстательной железы, болезни Альцгеймера и бокового амиотрофического склероза (БАС).
Исследование было частично поддержано Научно-исследовательским институтом профилактики рака Техаса и Национальным институтом здравоохранения.
О Медицинской школе UCI
Ежегодно Медицинская школа UCI обучает более 400 студентов-медиков и около 150 докторантов и магистров. Более 700 резидентов и стипендиатов проходят обучение в Медицинском центре UCI и дочерних учреждениях. Медицинский факультет предлагает степень доктора медицины; двойная программа подготовки ученых-медиков со степенью доктора медицины и доктора наук; и докторские степени и степени магистра в области анатомии и нейробиологии, биомедицинских наук, генетического консультирования, эпидемиологии, наук о гигиене окружающей среды, патологии, фармакологии, физиологии и биофизики, а также трансляционных наук. Студенты-медики также могут получить степень доктора медицины / магистра делового администрирования, степень доктора медицины / магистра в области общественного здравоохранения или степень доктора медицины / магистра в рамках одной из трех программ, основанных на миссии: санитарное просвещение для продвижения лидеров в области интегративной медицины (HEAL-IM), лидерство Образование для продвижения разнообразия для африканцев, чернокожих и карибцев (LEAD-ABC) и Программа медицинского образования для латиноамериканского сообщества (PRIME-LC).