Зарождение и развитие ассоциативной психологии
4.05.2011
Ассоцианизм, или ассоциативная психология, появился как самостоятельное направление в XVIII в. Эта школа положила начало выделению психологии в самостоятельную, независимую от философии науку, имеющую собственный предмет и тезаурус. В русле ассоциапизма изменилась и ориентация психологии с философской на естественнонаучную методологию, а также начались поиски объективного метода исследования и становление экспериментальной психологии.
Джордж Беркли (1685-1753 гг.) положил в основу своей концепции положения Локка о ведущей роли ощущений в формировании понятий. Отбросив идею о связи ощущений с внешним миром, Беркли во главу угла поставил понятие о внутреннем опыте, став родоначальником крайней формы субъективно-идеалистического воззрения на внешний мир и психическую жизнь. Его формула познания гласила: «Быть значит быть в восприятии», т.
Психологии восприятия Беркли посвятил трактат «Новая теория зрения» (1709). В нем он использовал принцип ассоциации, чтобы объяснить восприятие видимого пространства. Отношения между осязательными и зрительными ощущениями рассмотрены и в другой работе Беркли — «Принципы человеческого познания» (1710).
Давид Юм (1711 -1776 гг.). В работах «Трактат о человеческой природе» (1734-1737) и «Исследования о человеческом разумении» (1748) устраняется противопоставление объекта и субъекта, так как объект — комплекс восприятий, а субъект- процесс восприятия. Разница между этими разными группами восприятий лишь в их комбинациях, но не в происхождении.
Беркли и Юм, разрабатывая законы ассоциаций и связывая их с особенностями психики человека, все же рассматривали эти законы как частный случай своей концепции. Основателем же ассоциативной психологии, которая просуществовала как единственное собственно психологическое направление до начала XX в., по праву считается Давид Гартли (1705-1757 гг.).
В основу своей теории Гартли положил идею Локка об опытном характере знания, а также принципы механики Ньютона.
Учение об ассоциации Гартли, изложенное им в его книге «Размышления о человеке, его строении, его долге и упованиях» (1749), базируется на учении о вибрации, так как он считал, что вибрация внешнего эфира вызывает соответствующую вибрацию органов чувств, мышц и мозга. В структуре психики человека Гартли выделил два круга -большой и малый. Большой круг проходит от органов чувств через мозг к мышцам, т.
Гартли расширил сферу душевной жизни, включив в нее не только сознание, но и бессознательные процессы, и создал первую материалистическую теорию бессознательного.
Психика, по Гартли, состоит из нескольких основных элементов — ощущений, которые являются вибрацией органов чувств, представлений (вибрацией следов в белом веществе в отсутствие реального объекта) и чувств, отражающих силу вибрации. Гартли объяснял формирование самых сложных психических процессов, в том числе мышления и воли. Он считал, что в основе мышления лежит ассоциация образов предметов со словом (сводя мышление к процессу образования понятий), а в основе воли — ассоциация слова и движения.
Гартли объяснял формирование самых сложных психических процессов, в том числе мышления и воли. Он считал, что в основе мышления лежит ассоциация образов предметов со словом (сводя мышление к процессу образования понятий), а в основе воли — ассоциация слова и движения.
Исходя из идеи о прижизненном формировании психики, Гартли полагал, что возможности воспитания, воздействия на процесс психического развития ребенка поистине безграничны.
Ключевые слова: История, Наука
Источник: Р.В. Петрунникова, И.И. Заяц, И.И. Ахременко. История психологии — Минск.: Изд-во МИУ, 2009
| Материалы по теме |
|---|
Развитие отечественной психологии после 1917 года Константинов В.В., История психологии |
Возникновение психологии Карпов А.В., Общая психология |
Этапы развития отечественной социальной психологии Челдышова Н. |
Краткая история развития психологии Столяренко Л.Д., Психология и педагогика для технических вузов |
Развитие функционализма в психологии Р.В. Петрунникова, И.И. Заяц, И.И. Ахременко. История психологии — Минск.: Изд-во МИУ, 2009… |
Развитие дифференциальной психологии Р.В. Петрунникова, И.И. Заяц, И.И. Ахременко. История психологии — Минск.: Изд-во МИУ, 2009… |
История изучения психиатрии как науки Исаев Д. Н., Психиатрия детского возраста: психопатология развития: учебник для вузов. — СПб… |
Период открытого кризиса в психологии Р.В. Петрунникова, И.И. Заяц, И.И. Ахременко. История психологии — Минск.: Изд-во МИУ, 2009… |
 Б., Шпаргалка по социальной психологии
Б., Шпаргалка по социальной психологии28.
 Становление ассоциативной психологии в XVIII веке (Беркли, Юм, Гартли).
Становление ассоциативной психологии в XVIII веке (Беркли, Юм, Гартли).Английская психология в 18 веке развивалась от эмпиризма Локка к ассоцианизму, в трудах Беркли, Юм, Гартли.
Джордж Беркли (1684 – 1753) – непосредственный последователь Локка. Создал теорию зрительного восприятия («Опыт новой теории зрения»). Считалось, что зрение дает идеи света, цветов, пространства, фигуры, движения. Нам лишь кажется будто мы непосредственно видим протяженные тела в пространстве в трех измерениях. На самом деле удаленность предметов, величина, положение друг относительно друга глазом не воспринимает. Это скорее акт суждения основанный на опыте, а не на ощущениях. Пространственные характеристики вещей даются нам посредством мышечных ощущений, возникающих от поворотов глаз, напряжения его мышц. То, что мы называем зрительными ощущениями – комбинация собственно зрительных и осязательных ощущений.
Беркли
подчеркивал условность зрительных
ощущений, их знаковую природу: зрение
стало для осязания языком, стало выражать
содержание осязательного опыта. Биологическая полезность связи языка
и зрения: образуется в опыте, обеспечивает
правильное поведение.
Биологическая полезность связи языка
и зрения: образуется в опыте, обеспечивает
правильное поведение.
Теорию Беркли развил в 19 веке А. Бэн.
Давид Юм (1711 – 1776). Последователь Беркли. «Трактат о человеческой природе», «исследования о человеческом познании». Развил понятие ассоциации. Попытался представить человеческое познание как ассоциацию идей. Различает «впечатления» (восприятия) и «идеи» (их отражения). Идеи – воспроизведенные восприятия, более слабые впечатление, используются в мышлении и рассуждении. Идеи бывают простые и сложные, сложные образуются путем ассоциации. Объясняет всю работу познания механизмом ассоциации. Виды ассоциации:
Случайные и неправильные – ассоциации по закону смежности в пространстве или во времени.
Естественные и правильные – ассоциации по закону сходства и причинности.
Причинно-следственные
отношения сводятся к привычной
последовательности явлений (есть ли
каждый раз покушав мы чувствуем сытость,
мы через некоторое время делаем о том,
что причина нашей сытости является то,
что мы покушали).
Всю душевную жизнь Юм сводил к восприятиям, чувствованиям и ассоциациями между ними. Он отрицал существование духовной субстанции, Я личности, как особых образований, отличных от восприятий. Личность – не что иное, как связка или пучок различных восприятий, следующих друг за другом с непостижимой быстротой, и находящихся в постоянном течении, в постоянном движении.
Давид Гартли (1705 – 1757) воспринял идеи Локка об опытном происхождении душевной жизни, развил его представления об ассоциациях, дал первую законченную систему ассоциативной психологии. Опирался на идеи Ньютона. Для обоснования гипотезы о физиологических механизмах душевных процессах. Главный труд — «О человеке, его строении, его обязанностях и его упованиях». Развил учения о психике, как естественных началах. Все духовные способности объясняются через обращения к органической системе мозга. Элементы душевной жизни:
сенсации – ощущения
идеации – идеи ощущений
аффекции – удовольствия – неудовольствия
Из
этих трех элементов, с помощью механизмов
ассоциации строится душевная жизнь. В
основе элементов лежат вибрации –
физиологические процессы, возникающие
в веществе нервов и мозга под влиянием
внешних воздействий. Вибрации различны.
Различиям в вибрациях соответствуют
всё разнообразие наших простых людей
ощущений, представлений и чувствований.
Ассоциации – пассивное отражение
нервных связей в мозгу. Сочетаются не
ощущения или идеи, а состояние мозга,
которые сопровождаются ими – вибрации.
На основе ассоциаций образуется все
сложные представления, явления памяти,
понятия, суждения, произвольные движения,
аффекты (Страсти) воображение. Главы о
мышлении у Гартли нет. Рассматривается
понимание слов и предложений. Понимание
слова – образование ассоциации между
словом и значением, устанавливается в
детстве. Новые мысли – только новые
комбинации старых простых идей или
разложение сложных. Большое внимание
уделяет проблеме страстей и аффектов.
Страсти – движущая сила поведения.
Аффектов во врожденном состоянии не
существует. Страсти – продукт воспитания,
возникает по механизму ассоциации между
представлением о вещи и аффекцией.
В
основе элементов лежат вибрации –
физиологические процессы, возникающие
в веществе нервов и мозга под влиянием
внешних воздействий. Вибрации различны.
Различиям в вибрациях соответствуют
всё разнообразие наших простых людей
ощущений, представлений и чувствований.
Ассоциации – пассивное отражение
нервных связей в мозгу. Сочетаются не
ощущения или идеи, а состояние мозга,
которые сопровождаются ими – вибрации.
На основе ассоциаций образуется все
сложные представления, явления памяти,
понятия, суждения, произвольные движения,
аффекты (Страсти) воображение. Главы о
мышлении у Гартли нет. Рассматривается
понимание слов и предложений. Понимание
слова – образование ассоциации между
словом и значением, устанавливается в
детстве. Новые мысли – только новые
комбинации старых простых идей или
разложение сложных. Большое внимание
уделяет проблеме страстей и аффектов.
Страсти – движущая сила поведения.
Аффектов во врожденном состоянии не
существует. Страсти – продукт воспитания,
возникает по механизму ассоциации между
представлением о вещи и аффекцией.
Объясняет возникновение произвольных движений. Виды произвольных движений.
Полупроизвольные – связи ещё не достаточно укрепились
Произвольные – движения вызываются представлениями
Вторичные автоматизмы – произвольные движения возникают безотчетливо представления о движений. Достаточно лишь желание движения.
Психика не является врожденной, врождена только способность к аффектам, а также возможность получать ощущения. Всё что есть в душевной жизни – продукт воспитания.
Теория Гартли имело большое общественное значение. Для того, чтобы сделать человека тем, Что он есть, нужно только ощущающее начало и влияние тех обстоятельств в которых действительно находится человек.
Ассоциативное обучение и гиппокамп
Ассоциативная память определяется как способность узнавать и запоминать отношения между не связанными друг с другом элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов.
Wendy A. Suzuki, PhD
Каждый день мы узнаем и запоминаем множество новой информации от имен новых людей, которых мы встречаем, до лучшего блюда, которое можно заказать в том или ином ресторане, до расположения новой велосипедной дорожки. Эта новая информация сначала приобретается, и, если она усиливается в процессе, называемом консолидацией, в конечном итоге сохраняется в долговременной памяти. Структуры медиальной височной доли необходимы для этой способности приобретать новые долговременные воспоминания о фактах и событиях. Эта форма памяти называется декларативной памятью у людей и реляционной памятью у животных. Конвергентные результаты нейропсихологических исследований у людей, а также экспериментальные исследования повреждений и нейроанатомические исследования на животных показали, что ключевые структуры медиальной височной доли, важные для декларативной/реляционной памяти, включают гиппокамп вместе с окружающей энторинальной, периринальной и парагиппокампальной корой. Хотя ясно, что структуры медиальной височной доли необходимы для приобретения новых декларативных/реляционных воспоминаний, у нас все еще есть лишь рудиментарное понимание нормальных паттернов нейронной активности, лежащих в основе этой способности. Чтобы начать рассмотрение этого вопроса, моя лаборатория зарегистрировала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной/реляционной памяти, называемой ассоциативной памятью, которая определяется как способность изучать и запоминать отношения между не связанными между собой элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов. В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению.
Хотя ясно, что структуры медиальной височной доли необходимы для приобретения новых декларативных/реляционных воспоминаний, у нас все еще есть лишь рудиментарное понимание нормальных паттернов нейронной активности, лежащих в основе этой способности. Чтобы начать рассмотрение этого вопроса, моя лаборатория зарегистрировала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной/реляционной памяти, называемой ассоциативной памятью, которая определяется как способность изучать и запоминать отношения между не связанными между собой элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов. В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению. Наша долгосрочная цель состоит в том, чтобы понять эволюцию сигналов, связанных с обучением, по всей медиальной височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
Наша долгосрочная цель состоит в том, чтобы понять эволюцию сигналов, связанных с обучением, по всей медиальной височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
Задача, требующая нового ассоциативного обучения
Чтобы изучить закономерности нейронной активности во время формирования ассоциативной памяти, мы обучили двух обезьян выполнять задание на ассоциацию места и сцены. В этом задании от животных требовалось выучить новые ассоциации между конкретными сложными визуальными «сценами» и конкретными целевыми местами с вознаграждением. Мы знаем, что медиальная височная доля играет важную роль в нормальном выполнении этой задачи, поскольку повреждение этой области у обезьян приводит к значительному ухудшению способности запоминать новые ассоциации место-сцена (Brasted et al., 2003; Brasted et al., 2002). ; Мюррей и др., 2000; Уайз и Мюррей, 19 лет.99; Мюррей и Уайз, 1996 г. ; Рупняк и Гаффан, 1987). Схематическое изображение задачи показано на рисунке 1 . В каждом испытании обезьянам сначала показывают 4 идентичных целевых стимула, наложенных на сложную визуальную сцену (обычно изображение реальной сцены на открытом воздухе). После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают сигнал сделать одно движение глаз к одной из четырех периферийных целей на экране. Для каждой визуальной сцены только одна из 4 целей связана с наградой в виде сока. Каждый день животные методом проб и ошибок усваивали 2-4 новые ассоциации место-сцена. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонной» ассоциацией, которую животные видели за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с различным целевым местоположением (т. е. с севером, югом, востоком или западом). Реакция на эталонные сцены использовалась для контроля возможной двигательной активности в клетках гиппокампа.
; Рупняк и Гаффан, 1987). Схематическое изображение задачи показано на рисунке 1 . В каждом испытании обезьянам сначала показывают 4 идентичных целевых стимула, наложенных на сложную визуальную сцену (обычно изображение реальной сцены на открытом воздухе). После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают сигнал сделать одно движение глаз к одной из четырех периферийных целей на экране. Для каждой визуальной сцены только одна из 4 целей связана с наградой в виде сока. Каждый день животные методом проб и ошибок усваивали 2-4 новые ассоциации место-сцена. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонной» ассоциацией, которую животные видели за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с различным целевым местоположением (т. е. с севером, югом, востоком или западом). Реакция на эталонные сцены использовалась для контроля возможной двигательной активности в клетках гиппокампа.
Паттерны активности гиппокампа при изучении новых ассоциаций место-сцена
В наших первоначальных исследованиях мы сосредоточились на гиппокампе, медиальной структуре височной доли, долгое время участвующей в ассоциативном обучении и памяти (Eichenbaum and Cohen, 2001; Squire and Zola , 1996; Сковилл и Милнер, 1957). Сначала мы спросили, реагируют ли клетки гиппокампа по-разному на различные визуальные сцены, используемые в задаче. Мы обнаружили, что 61% изолированных клеток гиппокампа были вовлечены в задачу, поскольку они по-разному реагировали на разные сцены (то есть визуально избирательный ответ). Более того, в соответствии с нашей рабочей гипотезой мы обнаружили, что 28% избирательно реагирующих клеток (18% всей зарегистрированной популяции клеток гиппокампа) демонстрировали изменения в нервной активности в ходе испытаний, которые в значительной степени коррелировали с поведенческой кривой обучения животного для определенного сцена. Мы назвали эти клетки «изменяющимися клетками». Наблюдались две категории изменяющихся клеток. Устойчивые изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа). Многие из этих клеток демонстрировали резкое увеличение нейронной активности, которое соответствовало кривой поведенческого обучения животного для этой связи (рис. 2А-9).0012). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что меняющаяся клетка обычно меняет свою активность только для одной конкретной изученной сцены, в то время как ответы на другие изученные сцены не меняются с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением. Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т.
Наблюдались две категории изменяющихся клеток. Устойчивые изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа). Многие из этих клеток демонстрировали резкое увеличение нейронной активности, которое соответствовало кривой поведенческого обучения животного для этой связи (рис. 2А-9).0012). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что меняющаяся клетка обычно меняет свою активность только для одной конкретной изученной сцены, в то время как ответы на другие изученные сцены не меняются с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением. Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т. Е. Обучением реагировать на север). В соответствии с этой интерпретацией ранние правильные пробы, возможно, не вызвали большой активности, если движения были сделаны предварительно, но сильная двигательная активность наблюдается, как только животное начинает последовательно реагировать на север. Если бы эта интерпретация, основанная на моторике, была верна, мы ожидали бы увидеть аналогичные уровни активности этой клетки в ответ на эталонную сцену с тем же целевым расположением на севере. Такого никогда не было. Фактически, меняющиеся клетки обычно реагировали практически без активности на эталонную сцену с тем же вознагражденным целевым местоположением. Эти результаты подтверждают идею о том, что изменение активности связано с изучением новой связи между сценой и движением цели/глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные выучили 2 последовательных набора новых ассоциаций места и сцены.
Е. Обучением реагировать на север). В соответствии с этой интерпретацией ранние правильные пробы, возможно, не вызвали большой активности, если движения были сделаны предварительно, но сильная двигательная активность наблюдается, как только животное начинает последовательно реагировать на север. Если бы эта интерпретация, основанная на моторике, была верна, мы ожидали бы увидеть аналогичные уровни активности этой клетки в ответ на эталонную сцену с тем же целевым расположением на севере. Такого никогда не было. Фактически, меняющиеся клетки обычно реагировали практически без активности на эталонную сцену с тем же вознагражденным целевым местоположением. Эти результаты подтверждают идею о том, что изменение активности связано с изучением новой связи между сценой и движением цели/глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные выучили 2 последовательных набора новых ассоциаций места и сцены. Мы обнаружили, что изменение клеток, идентифицированных в первом наборе изученных ассоциаций место-сцена, никогда не сигнализировало об обучении второму новому набору ассоциаций место-сцена, даже когда поощряемое целевое местоположение было тем же самым (т. Е. Вознаграждалась северная цель).
Мы обнаружили, что изменение клеток, идентифицированных в первом наборе изученных ассоциаций место-сцена, никогда не сигнализировало об обучении второму новому набору ассоциаций место-сцена, даже когда поощряемое целевое местоположение было тем же самым (т. Е. Вознаграждалась северная цель).
Остальные 45% изменяющихся клеток демонстрировали другой паттерн учебной активности. Эти изменяющиеся клетки начинались с избирательной реакции на сцену либо во время сцены, либо в период задержки задачи в начале сеанса, задолго до того, как животное научилось ассоциации. Эти клетки сигнализировали об обучении, возвращаясь к исходной активности, и это возвращение к исходному уровню обычно антикоррелировало с кривой обучения животного для этой конкретной сцены (рис. 2В, ). Мы назвали эти клетки базовыми устойчивыми изменяющимися клетками. Важно отметить, что изменения в нервной активности, наблюдаемые в исходно устойчивых клетках, были столь же избирательны для конкретной заученной сцены, как и в устойчивых изменяющихся клетках. Подобные паттерны активности никогда не наблюдались для эталонной сцены с соответствующим вознаграждаемым целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Подобные паттерны активности никогда не наблюдались для эталонной сцены с соответствующим вознаграждаемым целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Таким образом, как клетки с устойчивым изменением, так и клетки с устойчивым изменением базового уровня обеспечивают высокоселективный сигнал о том, когда конкретная сцена изучена. Мы предполагаем, что эти избирательные увеличения и уменьшения нейронной активности, которые происходят в популяции гиппокампа, могут представлять собой сигнал обучения сети гиппокампа. Будет важно изучить обучающие сигналы по большему количеству клеток гиппокампа, записанные одновременно, чтобы лучше понять взаимодействие между клетками во время обучения.
Время активности гиппокампа во время изучения новых ассоциаций место-сцена
Важным вопросом для любого исследования, изучающего нейронные корреляты поведения, является определение причинно-следственной связи между наблюдаемыми паттернами нейронной активности и поведенческим результатом. Является ли наблюдаемая нейронная активность движущей силой поведения или она происходит ниже по течению от критических мест происхождения? Для решения этой проблемы использовались различные подходы. Например, как упоминалось выше, исследования поражений могут вовлекать конкретную область мозга в нормальное выполнение задачи, хотя сами по себе исследования поражений не могут определить паттерны нейронной активности, лежащие в основе этой функции. Исследования с электрической стимуляцией использовались для проверки влияния прямой стимуляции на выбор животных при проблемах сенсорной дискриминации (Salzman et al., 19).92; Salzman et al., 1990), но этот подход не применялся в исследованиях функции гиппокампа. Третий метод, который использовался для исследования взаимосвязи между нейронной активностью и поведением, заключается в изучении точного времени изменений нейронной активности по отношению к поведенческому обучению. Мы предполагаем, что те избирательные нейронные изменения, которые происходят до проявления поведенческого обучения, могут быть вовлечены в изменение поведения, в то время как те, которые происходят после поведенческого обучения, могут играть роль в укреплении вновь сформированной ассоциации.
Является ли наблюдаемая нейронная активность движущей силой поведения или она происходит ниже по течению от критических мест происхождения? Для решения этой проблемы использовались различные подходы. Например, как упоминалось выше, исследования поражений могут вовлекать конкретную область мозга в нормальное выполнение задачи, хотя сами по себе исследования поражений не могут определить паттерны нейронной активности, лежащие в основе этой функции. Исследования с электрической стимуляцией использовались для проверки влияния прямой стимуляции на выбор животных при проблемах сенсорной дискриминации (Salzman et al., 19).92; Salzman et al., 1990), но этот подход не применялся в исследованиях функции гиппокампа. Третий метод, который использовался для исследования взаимосвязи между нейронной активностью и поведением, заключается в изучении точного времени изменений нейронной активности по отношению к поведенческому обучению. Мы предполагаем, что те избирательные нейронные изменения, которые происходят до проявления поведенческого обучения, могут быть вовлечены в изменение поведения, в то время как те, которые происходят после поведенческого обучения, могут играть роль в укреплении вновь сформированной ассоциации. Чтобы ответить на этот вопрос, для всех изменяющихся клеток мы рассчитали количество попыток обучения и сравнили его с рассчитанным количеством попыток нейронных изменений (см. Wirth et al., 2003 для подробного описания используемых поведенческих и нейронных алгоритмов). Мы обнаружили, что изменяющиеся клетки гиппокампа могут опережать (14 примеров) параллельно (4 примера) и отставать (19 примеров).примеры) поведенческое обучение (рис. 3 ). Эти данные свидетельствуют о том, что гиппокамп участвует во всех стадиях процесса обучения, от нескольких попыток до того, как проявляется поведенческое обучение, когда наблюдаемая активность может быть вовлечена в управление поведенческими изменениями, лежащими в основе обучения, до нескольких попыток после обучения, когда активность может быть задействована. в укреплении новообразованной ассоциации.
Чтобы ответить на этот вопрос, для всех изменяющихся клеток мы рассчитали количество попыток обучения и сравнили его с рассчитанным количеством попыток нейронных изменений (см. Wirth et al., 2003 для подробного описания используемых поведенческих и нейронных алгоритмов). Мы обнаружили, что изменяющиеся клетки гиппокампа могут опережать (14 примеров) параллельно (4 примера) и отставать (19 примеров).примеры) поведенческое обучение (рис. 3 ). Эти данные свидетельствуют о том, что гиппокамп участвует во всех стадиях процесса обучения, от нескольких попыток до того, как проявляется поведенческое обучение, когда наблюдаемая активность может быть вовлечена в управление поведенческими изменениями, лежащими в основе обучения, до нескольких попыток после обучения, когда активность может быть задействована. в укреплении новообразованной ассоциации.
Резюме и будущие направления
Мы показали, что клетки гиппокампа обеспечивают сильные связанные с обучением паттерны нейронной активности, которые участвуют в начальном формировании новых ассоциативных воспоминаний.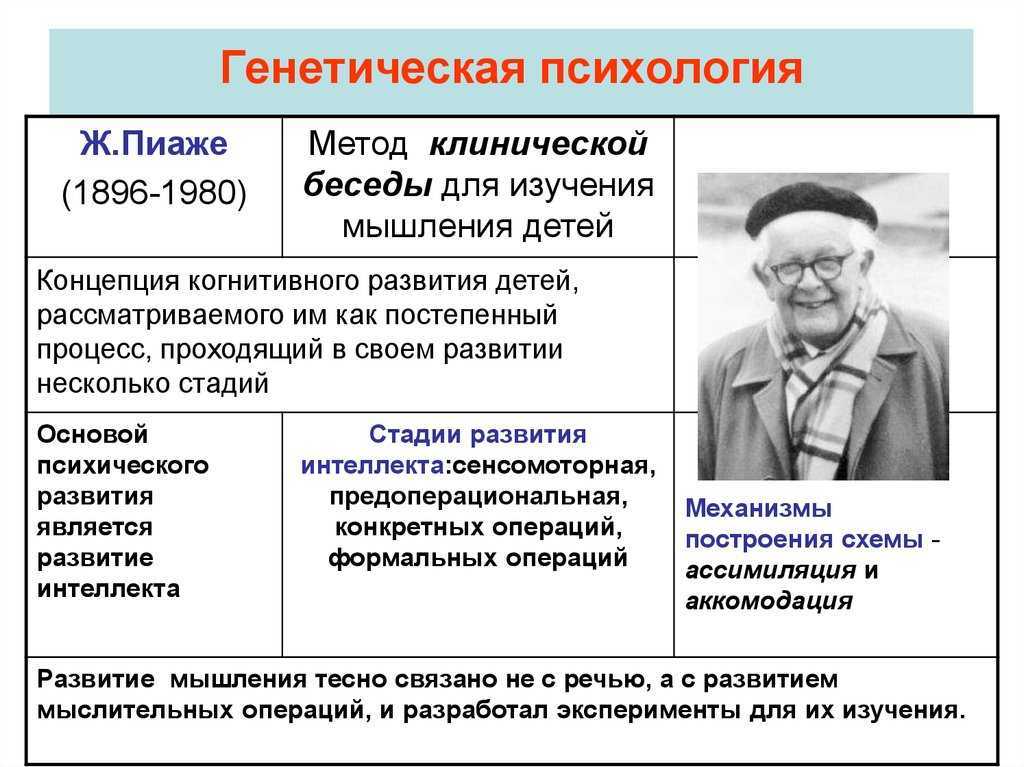 Поскольку эти изменения могут происходить до, в то же время или после обучения, эти данные позволяют предположить, что во время формирования новых ассоциативных воспоминаний может происходить постепенное вовлечение сети нейронов гиппокампа. Предыдущие исследования показали, что помимо гиппокампа клетки в нескольких других областях мозга, включая префронтальную кору (Asaad et al., 1998), лобные области, связанные с моторикой (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и стриатум (Brasted and Wise, 2004) демонстрируют схожие паттерны, связанные с обучением. активность при выполнении сходных ассоциативных учебных задач. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с моторикой областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также раннего укрепления и консолидации нового ассоциативного обучения.
Поскольку эти изменения могут происходить до, в то же время или после обучения, эти данные позволяют предположить, что во время формирования новых ассоциативных воспоминаний может происходить постепенное вовлечение сети нейронов гиппокампа. Предыдущие исследования показали, что помимо гиппокампа клетки в нескольких других областях мозга, включая префронтальную кору (Asaad et al., 1998), лобные области, связанные с моторикой (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и стриатум (Brasted and Wise, 2004) демонстрируют схожие паттерны, связанные с обучением. активность при выполнении сходных ассоциативных учебных задач. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с моторикой областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также раннего укрепления и консолидации нового ассоциативного обучения.
Ссылки
Асаад В. Ф., Райнер Г., Миллер Э. К. (1998) Нейронная активность в префронтальной коре приматов во время ассоциативного обучения. Нейрон 21: 1399-1407.
Ф., Райнер Г., Миллер Э. К. (1998) Нейронная активность в префронтальной коре приматов во время ассоциативного обучения. Нейрон 21: 1399-1407.
Brasted P.J., Bussey TJ, Murray EA, Wise SP (2002) Перерезка Fornix ухудшает условное зрительно-моторное обучение в задачах, связанных с непространственно дифференцированными ответами. J Нейрофизиол 87: 631-633.
Брастед П.Дж., Басси Т.Дж., Мюррей Э.А., Уайз С.П. (2003) Роль системы гиппокампа в ассоциативном обучении за пределами пространственной области. Мозг 126: 1202-1223.
Brasted P.J., Wise S.P. (2004) Сравнение связанной с обучением активности нейронов в дорсальной премоторной коре и стриатуме. Eur J Neurosci 19: 721-740.
Chen L.L., Wise S.P. (1995a) Активность нейронов в дополнительном поле глаза при приобретении условных глазодвигательных ассоциаций. J Нейрофиз 73: 1101-1121.
Chen L.L., Wise S.P. (1995b) Дополнительное поле глаза контрастирует с передним полем глаза во время приобретения условных глазодвигательных ассоциаций. J Нейрофиз 73: 1122-1134.
J Нейрофиз 73: 1122-1134.
Эйхенбаум Х., Коэн Н. Дж. (2001) От кондиционирования к сознательному воспоминанию. Нью-Йорк: Издательство Оксфордского университета.
Митц А. Р., Годшалк М., Уайз С. П. (1991) Зависимая от обучения активность нейронов в премоторной коре: активность при приобретении условных двигательных ассоциаций. J Neurosci 11: 1855-1872.
Мюррей Э. А., Басси Т. Дж., Уайз С. П. (2000) Роль префронтальной коры в сети произвольного зрительно-моторного картирования. Exp Br Res 133: 114-129.
Мюррей Э. А., Уайз С. П. (1996) Роль гиппокампа и нижележащей коры, но не миндалевидного тела, в зрительно-моторном условном обучении у макак-резусов. Behav Neurosci 110: 1261-1270.
Рупняк Н.М., Гаффан Д. (1987) Гиппокамп обезьяны и обучение пространственно направленным движениям. J Neurosci 7: 2331-2337.
Salzman C.D., Britten K.H., Newsome W.T. (1990) Корковая микростимуляция влияет на перцептивные суждения о направлении движения. Природа 346: 174-177.
Зальцман К. Д., Мурасуги К. М., Бриттен К. Х., Ньюсом В. Т. (1992) Микростимуляция зрительной области МТ: влияние на способность распознавания направления. J Neurosci 12: 2331-2355.
Scoville W.B., Milner B. (1957) Потеря недавней памяти после двустороннего поражения гиппокампа. J Neurol Neurosurg Psych 20: 11-21.
Сквайр Л. Р., Зола С. М. (1996) Структура и функции декларативных и недекларативных систем памяти. Proc Natl Acad Sci 93: 13515-13522.
Вирт С., Янике М., Франк Л.М., Смит А.С., Браун Е.Н., Судзуки В.А. (2003)Одиночные нейроны в гиппокампе обезьяны и изучение новых ассоциаций. Наука 300: 1578-1581.
Уайз С.П., Мюррей Э.А. (1999) Роль системы гиппокампа в условном моторном обучении: сопоставление предшественников с действием. Гиппокамп 9: 101-117.
Об авторе
Венди А. Судзуки получила докторскую степень в области неврологии в Калифорнийском университете в Сан-Диего в 1993 году и защитила докторскую диссертацию в Лаборатории нейропсихологии Национального института психического здоровья. В настоящее время она является адъюнкт-профессором нейробиологии и психологии в Нью-Йоркском университете. Ее исследования сосредоточены на понимании мозговой основы памяти. Чтобы ответить на этот вопрос, она использует поведенческую нейрофизиологию, записывая активность отдельных нейронов, когда обезьяны выполняют различные задачи, требующие памяти. Ее недавняя работа была сосредоточена на том, как клетки гиппокампа сигнализируют как о начальном формировании, так и о долговременном представлении ассоциативной памяти. Судзуки — лауреат премии Линдсли в области поведенческой неврологии (19 лет).94) и премию Troland в области экспериментальной психологии (2004).
В настоящее время она является адъюнкт-профессором нейробиологии и психологии в Нью-Йоркском университете. Ее исследования сосредоточены на понимании мозговой основы памяти. Чтобы ответить на этот вопрос, она использует поведенческую нейрофизиологию, записывая активность отдельных нейронов, когда обезьяны выполняют различные задачи, требующие памяти. Ее недавняя работа была сосредоточена на том, как клетки гиппокампа сигнализируют как о начальном формировании, так и о долговременном представлении ассоциативной памяти. Судзуки — лауреат премии Линдсли в области поведенческой неврологии (19 лет).94) и премию Troland в области экспериментальной психологии (2004).
Содержание, которое я только что прочитал:
Комментарий:
Suzuki, WA (2005, 1 февраля). Ассоциативное обучение и гиппокамп. Программа психологической науки . https://www.apa.org/science/about/psa/2005/02/suzuki
Нейробиологи открыли, как формируются ассоциативные воспоминания
- Скачать PDF Копировать
Отзыв Эмили Хендерсон, бакалавра наук 23 сентября 2021 г.
Запах свежеиспеченного шоколадного печенья когда-нибудь возвращал вас в послеобеденное время в дом вашей бабушки? Навевает ли когда-нибудь старая песня воспоминания о первом свидании? Способность запоминать отношения между не связанными между собой элементами (запах и место, песня и событие) известна как ассоциативная память.
Психологи начали изучать ассоциативную память в 1800-х годах, когда Уильям Джеймс описал это явление в своей книге «189».0 классический Основы психологии. Современные ученые согласны с тем, что структуры, ответственные за формирование ассоциативной памяти, находятся в медиальной височной доле, или в знаменитом «центре памяти» мозга, но конкретные задействованные клетки и то, как эти клетки контролируются, остаются загадкой. до настоящего времени.
Нейробиологи из Калифорнийского университета в Ирвайне обнаружили особые типы нейронов в центре памяти мозга, которые отвечают за приобретение новых ассоциативных воспоминаний. Кроме того, они обнаружили, как контролируются эти нейроны ассоциативной памяти. Мы полагаемся на ассоциативные воспоминания в повседневной жизни, и это исследование является важным шагом в понимании подробного механизма того, как эти типы воспоминаний формируются в мозгу.
Мы полагаемся на ассоциативные воспоминания в повседневной жизни, и это исследование является важным шагом в понимании подробного механизма того, как эти типы воспоминаний формируются в мозгу.
Хотя ассоциативная память является одной из самых основных форм памяти в нашей повседневной жизни, механизмы, лежащие в основе ассоциативной памяти, остаются неясными».
Кей Игараси, ведущий научный сотрудник, научный сотрудник Центра нейробиологии обучения и памяти и доцент кафедры анатомии и нейробиологии Медицинского факультета Калифорнийского университета в Ирвине
В исследовании, опубликованном сегодня в журнале Nature , впервые сообщается, что для приобретения новых ассоциативных воспоминаний необходимы специфические клетки латеральной энторинальной коры медиальной височной доли, называемые веерными клетками. контролируются дофамином, химическим веществом мозга, которое, как известно, участвует в нашем опыте удовольствия или вознаграждения.
В ходе исследования исследователи использовали электрофизиологические записи и оптогенетику для записи и контроля активности веерных клеток у мышей, когда они учатся связывать определенные запахи с вознаграждением. Этот подход привел исследователей к открытию, что веерные клетки вычисляют и представляют ассоциацию двух новых несвязанных элементов (запах и награда). Эти веерные клетки необходимы для успешного приобретения новых ассоциативных воспоминаний. Без этих ячеек можно восстановить предварительно изученные ассоциации, но нельзя получить новые ассоциации. Кроме того, для приобретения новых ассоциаций также требуется дофамин.
«Мы никогда не ожидали, что дофамин участвует в цепи памяти. Однако, когда накопились доказательства, постепенно стало ясно, что дофамин задействован», — сказал Игараши. «Эти эксперименты были для нас как детективная история, и мы в восторге от результатов».
Это открытие является важной частью головоломки понимания того, как в мозгу формируются воспоминания, и закладывает фундамент, на котором другие исследователи могут продолжать работу.