Фенотипы сосудистого старения по данным VaSera-скрининга у молодых людей с наличием артериальной гипертензии. Место дисплазии соединительной ткани | Евсевьева
1. Ben-Shlomo Y, Spears M, Boustred C, May M, Anderson SG, Benjamin EJ et al. Aortic pulse wave velocity improves cardio-vascular event prediction: an individual participant metaanalysis of prospective observational data from 17,635 subjects. J Am Coll Cardiol. 2014;63(7):636–646. doi:10.1016/j.jacc.2013.09.063
2. Mitchell GF, Hwang SJ, Vasan RS, Larson MG, Pencina MJ, Hamburg NM et al. Arterial stiffness and cardiovascular events: the Framingham Heart Study. Circulation. 2010;121(4):505–511. doi:10.1161/CIRCULATIONAHA.109.886655
3. Gielen S, Backer GDe, Piepoli M, Albus C, Astin F, Benlian P et al. The ESC Textbook of preventive cardiology. Oxford: University Press. 2016. p. 616 ISBN: 9780198795049.
4. Бурко Н. В., Авдеева И. В., Олейников В. Э., Бойцов С. А. Концепция раннего сосудистого старения. Рациональная фармакотерапия в кардиологии. 2019;15(5):742–749. doi:10.20996/1819-6446-2019-15-5-742-749
2019;15(5):742–749. doi:10.20996/1819-6446-2019-15-5-742-749
5. Неудахин Е. В., Морено И. Г. К вопросу о патогенезе атеросклероза и коррекции атерогенных нарушений у детей. Российский медицинский журнал. 2018;26(9):62–68.
6. Хоффман Дж. Атеросклероз. Детская кардиология. Пер. с англ. М.: Практика, 2006. C. 467–475.
7. Nilsson P. Early vascular ageing — a concept in development. Eur Endocrinology. 2015;11(1):26–31. doi:10.17925/EE.2015.11.01.26
8. Botto F, Obregon S, Rubinstein F, Scuteri A, Nilsson Р, Kotliar С. Frequency of early vascular aging and associated risk factors among an adult population in Latin America: the OPTIMO study. J Hum Hypertens. 2018;32(3):219–227. doi:10.1038/s41371-018-0038-1
9. Laurent S, Boutouyrie P, Cunha P, Lacolley C, Nilsson P. Concept of extremes in vascular aging from early vascular aging to supernormal vascular aging. Hypertension. 2019;74(2):218–228. doi:10.1161/HYPERTENSIONAHA.119
10. Suzuki K. Implications of CAVI in helth checkup. CAVI Now & Future. The Best Solution. 2015;(4):18–25.
CAVI Now & Future. The Best Solution. 2015;(4):18–25.
11. Mattace-Raso FUS, Hofman A, Verwoert GC, Witteman JCM, Wilkinson I, Cockcroft J. Reference values for carotidfemoral pulse wave velocity in the reference values for arterial stiffness’ collaboration database. Eur Heart J. 2010;31(19):2338–2350. URL: https://www.researchgate.net/publication/303074010_Reference_values_for_carotidfemoral_pulse_wave_velocity_in_the_reference_values_for_arterial_stiffness’_collaboration_database
12. Nilsson PM, Laurent S, Cunha PG, Olsen MH, Rietzschel E, Franco OH et al. Characteristics of healthy vascular ageing in pooled population-based cohort studies: the global Metabolic syndrome and Artery REsearch Consortium. J Hypertens. 2018;36(12):2340–2349. doi:10.1097/HJH.0000000000001824
13. Teemu JN, Bindu K, Gary FM, Ramachandran SV. Relative contributions of pulse pressure and arterial stiffnessto cardiovascular disease. The Framingham Heart Study. Hypertension. 2019;73(3):712–717. doi:10.1161/HYPERTENSIONAHA.118.12289
doi:10.1161/HYPERTENSIONAHA.118.12289
14. Ротарь О. П., Алиева А. С., Бояринова М. А., Толкунова К. М., Конради А. О. Концепция сосудистого возраста: какой инструмент для оценки выбрать в клинической практике? Кардиология. 2019;59(2):45–53.doi:10.18087/cardio.2019.2.10229
15. Chen W, Li S, Fernandez C, Sun D, Lai CC, Zhang T et al. Temporal relationship between elevated blood pressure and arterial stiffening among middle-aged black and white adults: The Bogalusa Heart Study. Am J Epidemiol. 2016;183:599–608. doi:10.1093/aje/kwv274
16. Cecelja M, Chowienczyk P. Dissociation of aortic pulse wave velocity with risk factors for cardiovascular disease other than hypertension: a systematic review. Hypertension. 2009;54(6):1328–1336. doi:10.1161/HYPERTENSIONAHA.109.137653
17. Mitchell GF. Arterial stiffness and hypertension: chicken or egg? Hypertension. 2014;64(2):210–214. doi.org/10.1161/HYPERTENSIONAHA.114.03449
18. Mitchell G, Powell J. Arteriosclerosis. A primer for “in focus” reviews on arterial stiffness. Arterioscler Thromb, Vasc Biol. 2020;40(5):1025–1027. doi:10.1161/ATVBAHA.120.314208
Arterioscler Thromb, Vasc Biol. 2020;40(5):1025–1027. doi:10.1161/ATVBAHA.120.314208
19. Евсевьева М. Е., Мириджанян Э. М., Бабунц И. В., Первушин Ю. В. Особенности липидного спектра крови и наследственная отягощенность по сердечно-сосудистым заболеваниям у лиц молодого возраста в зависимости от уровня здоровья. Кардиоваскулярная терапия и профилактика. 2005;4(6–2):77–81.
20. Евсевьева М. Е., Кумукова З. В. Особенности психологического статуса у лиц молодого возраста с признаками артериальной гипертензии. Российский психиатрический журнал. 2007;3:53–57.
21. Васюк Ю. А., Иванова С. В., Школьник Е. Л., Котовская Ю. В., Милягин В. А., Олейников В. Э. и др. Согласованное мнение российских экспертов по оценке артериальной жесткости в клинической практике. Кардиоваскулярная терапия и профилактика. 2016;15(2):4–19.
22. Rogoza AN, Konradi AO, Orlov AV, Soldatenkova NA, Rotar OP, Alieva AS et al. Еarly vascular aging: prevalence in Russian population according to different assessment methods. Gazzetta Medica Italiana Archivio per le Scienze Mediche. 2019;178(6):380–385.
Gazzetta Medica Italiana Archivio per le Scienze Mediche. 2019;178(6):380–385.
23. 2018 ЕОК/ЕОАГ Рекомендации по лечению больных с артериальной гипертензией. Российский кардиологический журнал. 2018;23(12):143–228. doi:10.15829/1560-4071-2018-12-143-228.2018
24. Диагностика, лечение, профилактика ожирения и ассоциированных с ним заболеваний (национальные клинические рекомендации), Санкт-Петербург, 2017. URL: https://scardio.ru/content/Guidelines/project/Ozhirenie_klin_rek_proekt.pdf
25. Golubnitschaja O. Flammer Syndrome in the Global Context — The “U-Shape” of Health Risks. In: Flammer Syndrome: From Phenotype to Associated Pathologies, Prediction, Prevention and Personalisation. Series: Advances in Predictive, Preventive and Personalised Medicine. 2019;11:1–8. Series Editor: O. Golubnitschaja. Springer Nature Switzerland AG. 2019; 375 p.
26. Bhaskaran K, Dos-Santos-Silva I, Leon DA, Douglas IJ, Smeeth L. Association of BMI with overall and cause-specific mortality: a population-based cohort study of 3·6 million adults in the UK. Lancet Diabetes Endocrinol. 2018:6(12):944–953. doi:10.1016/S2213-8587(18)30288-2
Lancet Diabetes Endocrinol. 2018:6(12):944–953. doi:10.1016/S2213-8587(18)30288-2
27. Musher DM, Abers MS, Corrales-Medina VF. Acute infection and myocardial infarction. N Engl J Med. 2019;380(2):171–176.
28. Qanitha А, Ajm de Mol B, Pabittei DR, Mappangara I, Van der Graaf Y, Dalmeijer GW et al. Infections in early life and premature acute coronary syndrome: a case-control Study. Eur J Prev Card. 2016;23(15):1640–1648.
29. Dratva J, Caviezel S, Schaffner E, Bettschart R, Kuenzli N, Schindler C et al. Infectious diseases are associated with carotid intima media thickness in adolescence. Atherosclerosis. 2015; 243(2):609–615.
30. Недифференцированные дисплазии соединительной ткани (проект клинических рекомендаций РНМОТ). Терапия. 2019;33(7):9–42.
31. Нарушения соединительной ткани в кардиологии. Диагностика и лечение. Российские рекомендации (I пересмотр). Российский кардиологический журнал. 2013;18(1):1–32.
32. Mancia G, Grassi G, Parati G, Zanchetti A. White coat hypertension: an unresolved diagnostic and therapeutic problem. Sprin Int Publish. 2015. 147 p. doi:10.1007/978-3-319-07410-8
White coat hypertension: an unresolved diagnostic and therapeutic problem. Sprin Int Publish. 2015. 147 p. doi:10.1007/978-3-319-07410-8
33. Кобалава Ж. Д., Котовская Ю. В., Кобзев Р. Ю. Характеристики центральной пульсовой волны у молодых мужчин с разными фенотипами артериального давления. Кардиология. 2010;50(2):36–40.
34. Иваненко В. В., Ротарь О. П., Конради А. О. Взаимосвязь показателей жесткости сосудов и центрального давления с поражением органов-мишеней. Сердечная недостаточность. 2012;13(2):101–104.
35. Кобалава Ж. Д., Котовская Ю. В., Ахметов Р. Е. Артериальная ригидность и центральное давление: новые патофизиологические и лечебные концепции. Артериальная гипертензия. 2010;16(2):126–133.
36. Милягин В. А., Милягина И. В., Абраменкова Н. Ю., Отрохова Е. В., Грекова М. В., Коптева В. В. и др. Неинвазивные методы исследования магистральных сосудов. Монография. Смоленск. 2012. 224 с.
37. Стражеско И. Д., Ткачева О. Н., Акашева Д. У., Дудинская Е. Н., Агальцов М.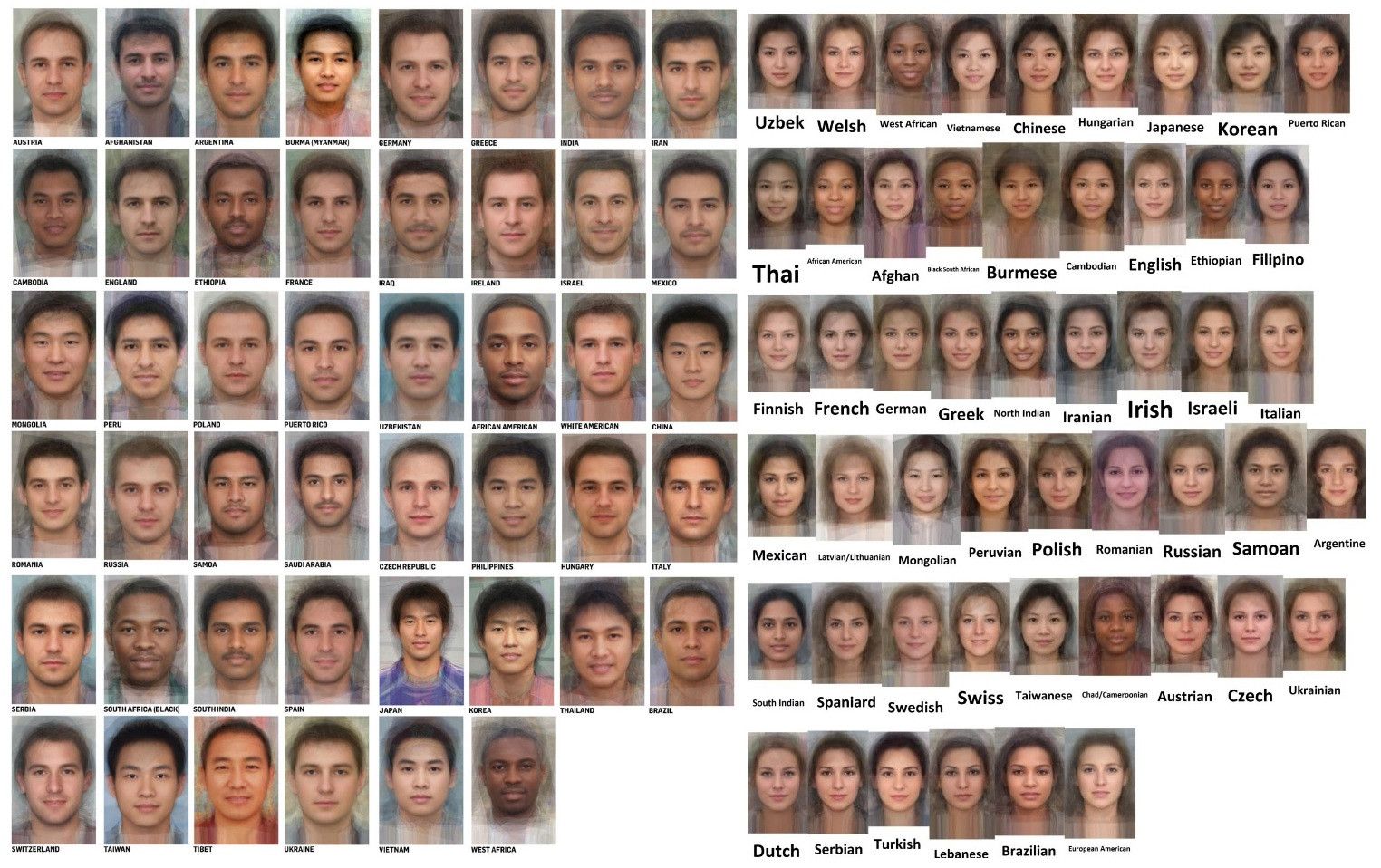 В., Кругликова А. С. и др. Взаимосвязь между различными структурно-функциональными характеристиками состояния артериальной стенки и традиционными факторами кардиоваскулярного риска у здоровых людей разного возраста. Часть 2. Рациональная фармакотерапия в кардиологии. 2016;12(3):244–252.
В., Кругликова А. С. и др. Взаимосвязь между различными структурно-функциональными характеристиками состояния артериальной стенки и традиционными факторами кардиоваскулярного риска у здоровых людей разного возраста. Часть 2. Рациональная фармакотерапия в кардиологии. 2016;12(3):244–252.
38. Rotar OP, Boyarinova MA, Moguchaia EV, Kolesova EP, Erina AM, Solntsev VN et al. Subclinical target organ damage in subjects with different components of metabolic syndrome. Clin Exp Hypertens. 2018;40(5):421–426. doi:10.1080/10641963.2017.1384488
39. Phillips R, Alpert B, Schwingshackl А, Huang X, Blakely D, Rovnaghi CR et al. Inverse relationship between cardio-ankle vascular index and body mass index in healthy children. J Pediatr. 2015;167(2):361–365.e1. doi:10.1016/j.jpeds.2015.04.042
40. Dangardt F, Osika W, Volkmann R, Gan LM, Friberg P. Obese children show increased intimal wall thickness and decreased pulse wave velocity. Clin Physiol Funct Imaging. 2008;28(5):287–293. doi:10. 1111/j.1475-097X.2008.00806.x
1111/j.1475-097X.2008.00806.x
41. Smirnova MI, Gorbunov VM, Volkov DA, Koshelyaevskaya YN, Deev AD, Furman NV et al. Characteristics of patients with reproducible masked hypertension. Eur Heart J. 2019;40(S 1): ehz747.0407. doi:10.1093/eurheartj/ehz747.0407
42. Franklin SS, O’Brien E, Staessen JA. Masked hypertension: understanding its complexity. Eur Heart J. 2017;38(15):1112–1118. doi:10.1093/eurheartj/ehw502
43. Mallion J, Ormezzano O, Barone-Rochette G, Neuder Y, Salvat M, Baguet JP. Hypertension artérielle masquée: myrhe ou réalité. La Presse Med. 2008;37(6 Pt 2):1034–1037. doi:10.1016/j.lpm.2008.01.022
44. Евсевьева М. Е., Коновалова Н. М., Ростовцева М. В., Слеткова Д. А., Щербакова Л. Е. Скрытая гипертензия и индекс аугментации центрального давления у юношей-студентов. Российский кардиологический журнал. 2016;21(S 4–1):29–30.
45. Милягин В. А., Ковалева О. А., Пунин Д. А. Определение центрального артериального давления — перспективное направление выявления начальной стадии артериальной гипертензии Терапия. 2021;7(2):39–49. doi:10.18565/therapy.2021.2.39-49
2021;7(2):39–49. doi:10.18565/therapy.2021.2.39-49
46. Семенкин А. А., Нечаева Г. И., Дрокина О. В., Махрова В. Н., Калиничева С. А., Живилова Л. А., Чиндарёва О. И. Возрастные аспекты структурно-функциональных изменений артерий у лиц с дисплазией соединительной ткани. Архив внутренней медицины. 2013;(3):46–50.
47. Семенкин А. А., Дрокина О. В., Нечаева Г. И., Живилова Л. А., Женатов А. Б. Недифференцированная дисплазия соединительной ткани как независимый предиктор структурно-функциональных изменений артерий. Кардиоваскулярная терапия и профилактика. 2013;12(3):29–34.
48. Евсевьева М. Е., Кошель В. И., Еремин М. В., Галькова И. Ю., Русиди А. В., Чудновский Е. В. и др. Скрининг ресурсов здоровья студентов и формирование внутривузовской профилактической среды: клинические, образовательные и воспитательно-педагогические аспекты. Медицинский вестник Северного Кавказа. 2015;10(1):64–69.
49. Aatola H, Koivistoinen T, Tuominen H, Juonala M, Lehtimäki T, Viikari JSA et al. Influence of child and adult elevated blood pressure on adult arterial stiffness: the cardiovascular risk in Young Finns Study. Hypertension. 2017;70(3):531–536.
Influence of child and adult elevated blood pressure on adult arterial stiffness: the cardiovascular risk in Young Finns Study. Hypertension. 2017;70(3):531–536.
Как гены влияют на внешность?
Общество вращается вокруг идеи привлекательности. Бренды зарабатывают на всеобщем желании быть красивыми и вместе со своими товарами продают мечту о красивой внешности. В то же время ученые бьются над загадками ДНК, которые определяют саму природу нашего облика. Хотя многое пока остается скрыто от пристального взгляда исследователей, часть загадок им уже удалось разгадать.
Содержание
- Какую информацию о внешности содержит ДНК: генотип
- Как на внешность влияет окружающий мир: фенотип
- ДНК-фенотипирование: мумии и преступники
- Все человечество состоит из больших семей: гаплогруппы
- Красота по наследству
- Внешние факторы
- Красота в уникальности
Какую информацию о внешности содержит ДНК: генотип
ДНК — инструкция, скрытая в каждом организме и одновременно объединяющая всех людей на планете. Каждая клетка, орган и часть тела строится согласно этой инструкции.
Каждая клетка, орган и часть тела строится согласно этой инструкции.
Но в то же время ДНК — это и уникальный код, который делает людей совершенно разными. Цвет кожи и глаз, рост, размер носа, длина рук и ног, расстояние между глазами. Какими они будут?
Облик человека практически напрямую связан с внешностью его родителей. Вернее, с особым миксом их хромосом. Всего в норме хромосом 23 пары, половину их них человек получает от отца, а половину — от матери. Внутри хромосом существуют определенные участки, определяющие различные физические особенности.
Участки ДНК, в которых закодированы определяющие признаки человека, в том числе его физические особенности, называются генами. Гены составляют генотип.
Вариации одного и того же гена называются аллелями. Они также переходят нам от матери и отца. Аллели могут быть идентичными или разными. А их точное расположение на хромосоме известно как локус.
Принцип влияния аллелей, которые определяют облик ребенка, особенно легко разобрать на классической задаче с геном, кодирующим цвет глаз (на самом деле за цвет глаз отвечает больше одного гена, однако данный пример удобен для объяснения). Например, у отца карие глаза, в то время как у матери — голубые.
Например, у отца карие глаза, в то время как у матери — голубые.
Аллель карего цвета является доминантным, а аллель голубого – рецессивным. Если ребенок унаследует два разных аллеля, у него будут карие глаза. Чтобы у ребенка был шанс унаследовать голубые глаза, оба его родителя должны быть носителями аллеля голубых глаз (при этом они оба могут быть кареглазыми).
Кроме цвета глаз, генотип определяет цвет волос, рост, размер обуви. Казалось бы, теперь можно посмотреть на внешность человека и определить его генотип? Но нет, это будет похоже на игру в наперстки: определить конкретные аллели может лишь генетический тест. В противном случае вы не сможете знать ответ наверняка. Дело в том, что доминантный аллель может скрывать наличие рецессивного.
Вернемся к примеру с глазами: генетики обозначают аллель, отвечающий за карие глаза, буквой B, а за голубые — буквой b. Кареглазый человек может иметь либо две коричневые версии гена OCA2 (BB), либо коричневую и голубую версии (Bb). Оба генотипа BB и Bb дают карие глаза. Поэтому, если у вас карие глаза, вы не можете знать точно, носителем какого из двух генотипов вы являетесь на самом деле.
Оба генотипа BB и Bb дают карие глаза. Поэтому, если у вас карие глаза, вы не можете знать точно, носителем какого из двух генотипов вы являетесь на самом деле.
Генетический тест Атлас поможет узнать больше о генах, которые повлияли на цвет и строение радужки глаз. Мы анализируем генетические варианты, связанные с пигментацией, бороздами и криптами.
В вопросе изучения генотипов ученые сходятся в том, что они еще только подступают к началу исследований. Пока что генетики с осторожностью смотрят на возможность создания точного физического образа человека, основываясь на его ДНК. И все же за последние десятилетия ученым удалось ощутимо продвинуться в своих исследованиях. Поэтому сегодня они говорят о том, что могут с 75% вероятностью указать происхождение, цвет глаз и волос человека на основе его ДНК.
Как на внешность влияет окружающий мир: фенотип
Внешность — это нечто большее, чем генетически запрограммированный рост, цвет глаз или волос. Каждый человек является носителем уникального набора внешних и внутренних характеристик — фенотипа. Фенотип формируется под воздействием не только генетики, но и факторов окружающей среды.
Фенотип формируется под воздействием не только генетики, но и факторов окружающей среды.
К факторам окружающей среды можно отнести питание, климат, в том числе температуру и влажность, экологические условия и многое другое.
Так, цвет кожи контролируется генами, которые определяют количество и тип меланина. Этот пигмент отвечает за оттенок кожи, глаз, волос, а также участвует в некоторых обменных процессах.
Воздействие ультрафиолета в регионе проживания может вызвать потемнение существующего меланина и способствовать усилению меланогенеза и, следовательно, формированию более темной кожи.
Фенотип включает в себя набор наблюдаемых характеристик или черт, таких как рост, вес, форма носа, цвет волос, а также наше поведение. При этом ключевое различие между фенотипом и генотипом заключается в том, что генотип наследуется от родителей, а фенотип формируется в процессе индивидуального развития под влиянием внешней среды.
К примеру, представьте, что согласно генотипу человек должен был иметь высокий рост, однако переболел в детстве рахитом и не достиг ожидаемых значений. В данном случае его фенотип не совпал с генотипом.
В данном случае его фенотип не совпал с генотипом.
Предполагается, что, выбирая себе партнера, мы в первую очередь обращаем внимание на фенотипические признаки. Например, симметрию черт лица и тела. Однако исследователям до сих пор неясно, являются ли они основным признаком при оценке потенциальных кандидатов для создания пары. Или же симметрия коррелирует с другими чертами, влияющими на физическую привлекательность.
ДНК-фенотипирование: мумии и преступники
Фенотипирование открывает множество новых возможностей в работе судебно-медицинских экспертов. Оно делает доступными новые методы прогнозирования внешности человека на основе найденной на месте преступления ДНК.
Фенотипирование ДНК позволяет следователям в некоторых странах предсказывать внешность человека: от определения круга подозреваемых на основе образцов ДНК, найденных на месте преступления, до идентификации неизвестных человеческих останков и раскрытия дел о пропавших без вести.
Сегодня даже небольших образцов ДНК может оказаться достаточно для получения информации о чертах лица и других внешних признаках: цвете волос, глаз, кожи. Однако результаты фенотипирования не считаются точным доказательством и их нельзя использовать для идентификации людей. Поэтому главная цель судебно-медицинского фенотипирования ДНК в том, чтобы сузить число потенциальных подозреваемых.
Однако результаты фенотипирования не считаются точным доказательством и их нельзя использовать для идентификации людей. Поэтому главная цель судебно-медицинского фенотипирования ДНК в том, чтобы сузить число потенциальных подозреваемых.
В настоящее время на пути исследователей, занимающихся фенотипированием, стоят две преграды:
- нехватка данных, которые позволили бы соотнести генетические варианты с определенными фенотипическими признаками;
- необходимость установить значение этих вариантов в фенотипическом предсказании, чтобы в дальнейшем определить точность предсказания внешних признаков на основе ДНК.
Понимание каждой генетической детали и механизма не обязательно для предсказания того, как будет выглядеть лицо. Не нужно знать, как устроен двигатель автомобиля, чтобы вывести статистическую закономерность движения автомобилей по проспекту.
Поэтому ученые активно используют уже существующие наработки не только для того, чтобы искать людей, но и для того, чтобы путешествовать в прошлое. Фенотипирование активно используется для изображения представителей древнего мира: людей и животных. Например, для воссоздания облика до мумификации.
Фенотипирование активно используется для изображения представителей древнего мира: людей и животных. Например, для воссоздания облика до мумификации.
Генетическое исследование древних образцов очень похоже на судебно-медицинский анализ. В обоих случаях невозможно сравнить генетическую информацию с другими членами семьи и практически невозможно добиться полной индивидуальной идентификации.
Поэтому весь процесс похож скорее на прогнозирование внешних характеристик человека. При этом его биогеографическое происхождение, скрытое в ДНК, безусловно, может стать решающим. И так мы подходим к миру гаплогрупп.
Все человечество состоит из больших семей: гаплогруппы
Гаплогруппы — это сообщество людей или гаплотипов (групп аллелей из одной хромосомы, обычно наследуемых вместе), которых объединяет общий предок, передавший всем одну и ту же мутацию (полиморфизм).
Изучение гаплогрупп помогает нам понять свое происхождение.
По результатам Генетического теста Атлас можно узнать, насколько близок ваш геном к геному неандертальцев — людей, живших десятки тысяч лет назад на территории Европы и Азии и не являющихся нашими прямыми предками.

Галпрогруппы также важны для понимания истории. Данные о гаплотипах хранятся в биобанках и помогают понимать и даже дописывать генетический ландшафт стран и наций. Так, одно из последних таких исследований помогло изучить степень изоляции древних европейцев. В результате стало ясно, что число предков современных британцев, живущих за пределами острова, крайне широко.
Анализ гаплогрупп помогает и современным археологам. Например, при изучении останков, обнаруженных в Мезмайской пещере на Северном Кавказе (она находится на границе, разделяющей Европу и Азию). Анализ генома костей древней женщины показал тесные генетические связи ранних неандертальцев среднего палеолита, которых позднее сменила генетически отдаленная более поздняя группа, обитавшая в тех же географических районах.
Кроме того, изучая гаплогруппы, ученые могут определить закономерности генетической изменчивости, связанные со здоровьем. Например, если полиморфизм связан с определенным заболеванием, ученые могут исследовать связанные участки ДНК, чтобы попытаться идентифицировать ген или гены, ответственные за возникновение заболевания.
Красота по наследству
Несмотря на важное значение, которое общество придает красоте, с научной точки зрения мы знаем довольно мало о том, почему одни лица кажутся «красивее» других. Исследовательская команда из Университета Вашингтона попыталась найти ответ на важный вопрос: «передается ли красота по наследству»? А если это так, то какой ген отвечает за внешнюю привлекательность?
Для проведения исследования была использована генетическая выборка из 4383 европейцев, включая мужчин и женщин. Для начала добровольцев попросили оценить фотографии по уровню привлекательности запечатленных на них людей.
Затем ученые проанализировали геном «красавчиков» и смогли идентифицировать два значимых локуса привлекательности для всего генома. В результате они выделили несколько генов-кандидатов, которые могли бы стать ответственными за передачу «привлекательности».
Интересно, что эти гены отличались для мужчин и женщин. Женская красота оказалась связана с генами, влияющими на массу тела. А вот варианты, связанные с привлекательностью мужчин, были ассоциированы с генами, влияющими на уровень холестерина в крови.
А вот варианты, связанные с привлекательностью мужчин, были ассоциированы с генами, влияющими на уровень холестерина в крови.
Исследователи пришли к выводу, что не существует единого «главного гена», влияющего на привлекательность. Вместо этого, привлекательность, скорее всего, контролируется большим количеством слабых генетических ассоциаций со сложными регулирующими эффектами.
Более того, когда люди смотрят друг на друга, они анализируют фенотип и неосознанно «считывают» генетические различия между популяциями. Поэтому существует вероятность, что отбор по внешним признакам сыграл важную роль в эволюции нашего вида. При этом в вопросах выживания, вполне вероятно, была важна не только красота, но и уникальность.
Внешние факторы
Гены — это лишь часть того, что определяет внешний вид человека. Мы живем в социуме и постоянно контактируем с внешним миром. Потому окружающая среда и образ жизни играют не менее важную роль.
Хотя ученым и неизвестно, насколько эта роль велика. Но мы точно знаем, что образ жизни может повлиять на активацию определенных генов, а те, в свою очередь, сыграют свою роль в формировании внешности человека.
Но мы точно знаем, что образ жизни может повлиять на активацию определенных генов, а те, в свою очередь, сыграют свою роль в формировании внешности человека.
Возьмем двух подруг: Анну и Ольгу. У обеих есть генетическая предрасположенность к ожирению. Предположим, что Анна живет в более богатой семье. Ее родители могут обеспечить дочери разнообразные и питательные блюда. В их меню меньше соли, жиров и добавленного сахара. Анна с детства учится любить овощи и фрукты.
У родителей Анны есть свободное время, которое они проводят вместе с дочерью: занимаются спортом и много гуляют. Сбалансированное питание и физическая активность не позволят предрасположенности к ожирению проявиться.
В семье Ольги другая ситуация. Ее воспитывают родители с низким доходом, которые живут в районе, где свежая, здоровая пища дефицитна и дорога. Поскольку ее родители не могут позволить себе фрукты и овощи, Ольга часто ест замороженные, готовые блюда и продукты быстрого приготовления, в которых много сахара, жира и соли.
Родители Ольги работают на нескольких работах, и девочка предоставлена сама себе. Как правило, она проводит время в одиночестве перед телевизором. Привычки Ольги в еде и дефицит физической активности могут способствовать проявлению генетической склонности. И вероятно, что она вырастет с лишним весом.
Генотип и фенотип, с которыми мы рождаемся, важны. Они словно наш багаж, который нужно аккуратно пронести через всю жизнь. Состав багажа может повлиять на путешествие, но и маршрут может отразиться на состоянии багажа.
Жизненный опыт Анны и Ольги влияет на то, когда и как проявят себя их гены. Взаимосвязь между нашими генами и поведением может со временем меняться по мере того, как мы получаем новый опыт.
Красота в уникальности
Внешний вид имеет отчасти практическую и даже финансовую ценность. Так, по некоторым данным, красивые люди получают больше внимания, денег, уважения.
Люди хотят быть привлекательными. По статистике пластический хирург в 2021 году в среднем провел 320 хирургических операций.Для сравнения в 2020 году их число было почти на треть меньше — 220.
 Для сравнения в 2020 году их число было почти на треть меньше — 220.
Для сравнения в 2020 году их число было почти на треть меньше — 220.Но красота — нечто большее, чем симметрия и размер черт лица. На самом деле это сложное сочетание внешней картины, ума и обаяния.
Представления о канонической, маркетинговой красоте постоянно меняются. Сегодня мы живем в открытом социуме, готовым принимать красоту во всем ее разнообразии больше чем когда-либо. Те, кого мы считаем красивыми, на самом деле отражают наши субъективные представления о красоте. Учитывая, что эти представления у разных людей отличаются, мы можем сказать, что живем в мире, где прекрасны все. И уникальны. Потому что именно такими нас делает ДНК.
Больше о внешности и ДНК в блоге Атласа:
- Как гены влияют на цвет глаз
- Как ДНК определяет цвет и форму волос
- Как гены влияют на качество зубов
The Center for Nanoscale Science. Penn State University
Jim Dawson. Defining a Face: What Can DNA Phenotyping Really Tell Us About An Unknown Sample?
J.
E. Scheib, S.W. Gangestad, R. Thornhill. Facial attractiveness, symmetry and cues of good genes.University College London. The genetics of human physical appearance.
Peter M. Schneider, Barbara Prainsack, Manfred Kayser. The Use of Forensic DNA Phenotyping in Predicting Appearance and Biogeographic Ancestry.
Cláudia Gomesa, Manuel Fondevilab, Sara Palomo-Díeza, Antonio F.Pardiñasc, CésarLópez-Matayoshiad, Carlos Baeza-Richera, Ana María López-Parraa, María Victoria Lareub, Belén Lópeze, Eduardo Arroyo-Pardo. Phenotyping the ancient world: The physical appearance and ancestry of very degraded samples from a chalcolithic human remains.
Edmund Gilbert, Ashwini Shanmugam, Gianpiero L. Cavalleri. Revealing the recent demographic history of Europe via haplotype sharing in the UK Biobank.
Tatiana V. Andreeva, Andrey D. Manakhov, Fedor E. Gusev, Anton D. Patrikeev, Lyubov V. Golovanova, Vladimir B. Doronichev, Ivan G.
Bowen Hu, Ning Shen, James J. Li, Hyunseung Kang, Jinkuk Hong, Jason Fletcher, Jan Greenberg, Marsha R. Mailick, Qiongshi Lu. Genome-wide association study reveals sex-specific genetic architecture of facial attractiveness.
The Aesthetic Society. Aesthetic Plastic Surgery Statistics.
 E. Scheib, S.W. Gangestad, R. Thornhill. Facial attractiveness, symmetry and cues of good genes.
E. Scheib, S.W. Gangestad, R. Thornhill. Facial attractiveness, symmetry and cues of good genes.
| СПИНА |
Методы
Методы НАЗАД Антропологические типы, представленные на этом сайте, получены в результате качественного анализа множества различных антропологических исследований, в основном написанных в 20-м веке, в основном на немецком, английском, французском, итальянском, русском, шведском, венгерском или польском языках. Классические антропологические типологии в одних случаях были похожи друг на друга, а в других сильно отличались друг от друга. Для получения типов, изображенных на этом сайте, сравнивались типовые описания разных авторов, и в случае сходства несколько типов объединялись в один, а в случае различия сохранялись исходные типы. Чтобы понять, почему был выбран именно этот подход, какие существуют альтернативы, как классические исследования 20-го века определили их типы и как они соотносятся с современными, часто филогенетическими исследованиями 21-го века, здесь дается методологический обзор. Классические антропологические типологии в одних случаях были похожи друг на друга, а в других сильно отличались друг от друга. Для получения типов, изображенных на этом сайте, сравнивались типовые описания разных авторов, и в случае сходства несколько типов объединялись в один, а в случае различия сохранялись исходные типы. Чтобы понять, почему был выбран именно этот подход, какие существуют альтернативы, как классические исследования 20-го века определили их типы и как они соотносятся с современными, часто филогенетическими исследованиями 21-го века, здесь дается методологический обзор. В большинстве случаев на этом сайте приоритет отдается интегральному подходу. Поскольку колонизация и глобализация быстро изменили антропологический состав многих регионов мира, все карты и описания относятся к 1500 году , до того, как эти процессы начались в больших масштабах. Иллюстрировать нынешнее современное состояние разнообразия было бы противоречиво, поскольку в настоящее время любой тип может появиться в любой точке мира. Далее рассматривается, откуда берутся определения антропологических типов, являются ли они четкими биологическими подразделениями или же они «падают с неба» и являются совершенно случайными, рукотворными, даже политическими определениями. В конце концов, решается вопрос: «Почему важны фенотипы?». Вся информация является результатом частного хобби-проекта и не основана на финансировании каких-либо исследований. Хотя сайт старается быть максимально точным, некоторые результаты носят спекулятивный характер и могут содержать ошибки. Если вы найдете ошибки, пожалуйста, свяжитесь со мной. |
 Кроме того, многие смешанные группы еще не стабилизированы, т.е. члены одной семьи могут сильно различаться. Стабилизация обычно занимает несколько столетий.
Кроме того, многие смешанные группы еще не стабилизированы, т.е. члены одной семьи могут сильно различаться. Стабилизация обычно занимает несколько столетий.1.) Численный подход
Численный подход анализирует комбинации антропометрических признаков на индивидуальном уровне (в отличие от популяционного подхода). Эта методология также является основой интегрального подхода и популяционного подхода и была частью анализа для большинства антропологов. Однако, в отличие от, например, интегральный подход, числовой подход только использует метрики для определения типа. Это имеет то преимущество, что никакие «человеческие предубеждения» не изменяют результаты, например. субъективное впечатление, которое некоторые фенотипы производят на человека-наблюдателя. Кроме того, анализ данных может выявить взаимосвязи, которые иначе невозможно наблюдать. Таким образом, числовые типы являются исключительно статистическим результатом. Базовая схема классификации европейских типов основана на числовом подходе: Эта методология также является основой интегрального подхода и популяционного подхода и была частью анализа для большинства антропологов. Однако, в отличие от, например, интегральный подход, числовой подход только использует метрики для определения типа. Это имеет то преимущество, что никакие «человеческие предубеждения» не изменяют результаты, например. субъективное впечатление, которое некоторые фенотипы производят на человека-наблюдателя. Кроме того, анализ данных может выявить взаимосвязи, которые иначе невозможно наблюдать. Таким образом, числовые типы являются исключительно статистическим результатом. Базовая схема классификации европейских типов основана на числовом подходе:Таблица 1: Пример для европейских типов на основе численного подхода по комбинации трех антропометрических признаков (Lebzelter, 1929):
Головной указатель не менее 81 считается короткоголовым, поэтому любой высокий светловолосый человек с головным указателем 80,9 будет классифицирован как нордид с использованием этого подхода, основанного на трех признаках. 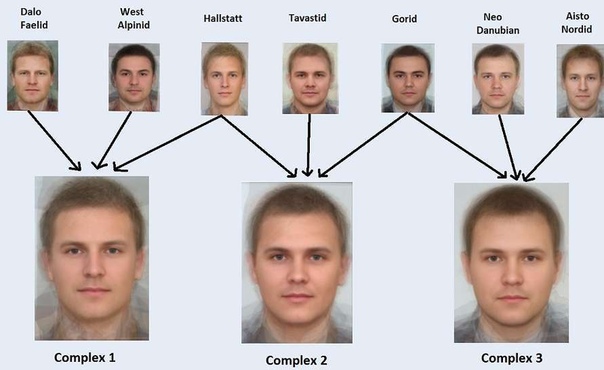 С индексом 81 он был бы Норидом, если не учитывать другие антропометрические особенности.
Численный подход был популярен, например, в польской и югославской школах антропологии, которые часто использовали более трех метрик, но все же лишь ограниченное число. Численными методами иногда выявлялись лаппоидные или южномонголоидные типы среди коренных американцев или австралоидные и эскимоидные типы в Африке, хотя эти люди имеют мало родства с коренными американцами и африканцами соответственно.
Это обнажает слабое место использования исключительно численного подхода: если принимать во внимание лишь несколько антропометрических признаков, огромные различия в других признаках не будут учитываться и, следовательно, игнорироваться. Таким образом, числовой антрополог сильно зависит от своих данных и их качества. Кроме того, с каждым метрическим признаком количество комбинаций быстро возрастает до огромных чисел (см. рис. 1). Таким образом, необходимо выбрать относительно небольшое количество признаков. С индексом 81 он был бы Норидом, если не учитывать другие антропометрические особенности.
Численный подход был популярен, например, в польской и югославской школах антропологии, которые часто использовали более трех метрик, но все же лишь ограниченное число. Численными методами иногда выявлялись лаппоидные или южномонголоидные типы среди коренных американцев или австралоидные и эскимоидные типы в Африке, хотя эти люди имеют мало родства с коренными американцами и африканцами соответственно.
Это обнажает слабое место использования исключительно численного подхода: если принимать во внимание лишь несколько антропометрических признаков, огромные различия в других признаках не будут учитываться и, следовательно, игнорироваться. Таким образом, числовой антрополог сильно зависит от своих данных и их качества. Кроме того, с каждым метрическим признаком количество комбинаций быстро возрастает до огромных чисел (см. рис. 1). Таким образом, необходимо выбрать относительно небольшое количество признаков. Однако рассмотрение только 5, 10 или даже 50 антропометрических признаков будет недостаточным для описания такого сложного организма, как человек, они могут отразить лишь небольшую часть фенотипа. Кроме того, погрешность измерения или разные способы измерения (например, существует несколько методов получения носового индекса) или метрические признаки, измененные окружающей средой, легко могут повлиять на классификацию. Человеческая предвзятость также легко вносится при выборе данных, полученных для анализа.
Таким образом, разумные результаты при численном подходе достигаются только при высоком качестве данных. Сегодня, например, можно выполнить 3D-сканирование тела и получить тысячи антропометрических признаков и добавить генетическую информацию. Это было невозможно для классических антропологов, и у них не было подходящих компьютеров для обработки таких больших объемов данных. В любом случае, несмотря на недостатки численного подхода, рассмотрение антропометрических признаков полезно при любом типологическом подходе, следует проявлять осторожность только в том случае, если не учитывается никакая другая информация. Однако рассмотрение только 5, 10 или даже 50 антропометрических признаков будет недостаточным для описания такого сложного организма, как человек, они могут отразить лишь небольшую часть фенотипа. Кроме того, погрешность измерения или разные способы измерения (например, существует несколько методов получения носового индекса) или метрические признаки, измененные окружающей средой, легко могут повлиять на классификацию. Человеческая предвзятость также легко вносится при выборе данных, полученных для анализа.
Таким образом, разумные результаты при численном подходе достигаются только при высоком качестве данных. Сегодня, например, можно выполнить 3D-сканирование тела и получить тысячи антропометрических признаков и добавить генетическую информацию. Это было невозможно для классических антропологов, и у них не было подходящих компьютеров для обработки таких больших объемов данных. В любом случае, несмотря на недостатки численного подхода, рассмотрение антропометрических признаков полезно при любом типологическом подходе, следует проявлять осторожность только в том случае, если не учитывается никакая другая информация. Рисунок 1: Количество возможных комбинаций типов дает определенное количество антропометрических признаков, которые имеют два или три проявления признака: |
2.) Интегральный подход
| Интегральный подход является предпочтительным для типов, представленных на этом сайте. Только в том случае, если у антропологов, использующих интегральный подход, было мало информации о конкретном месте в мире, для региона включались типы других подходов. Такие подходы были распространены в немецкой, итальянской и французской антропологических школах. Интегральный подход родственен численному, точнее, строится на нем. Он также использует статистические комбинации антропометрических признаков (см. таблицу 1), но не полностью полагается на них. Вместо этого он добавляет большое количество дополнительной качественной информации. Это включает в себя качественную оценку физических признаков и того, как они выглядят для глаза человека-наблюдателя, особенно особенностей мягких тканей человека, которые трудно измерить. Рисунок 2: Динарид или Нордид? (сравните таблицу 1) Для любого определенного набора идеальных типов (типологии) люди будут группироваться в три категории: 1.) Типичные представители идеального типа, проявляющие ~80-100% определенных признаков. 2.) «Смешанные» особи, которые соответствуют (линейной) комбинации двух или более типов. 3.) Лица, которые не могут быть охвачены типологией. Чтобы найти набор типов, интегральный антрополог, например, может попытаться максимизировать количество людей в группе 1 и минимизировать количество людей в группе 3. В идеале определенный тип должен доминировать в определенных популяциях, социальных группах или семьях. и быть характерным для них. Интегральный подход сталкивается с рядом проблем. Большой объем информации, которую необходимо рассмотреть, может быть слишком сложным, чтобы найти набор возможных типов, который лучше всего описывает человеческое разнообразие. В целом интегральный подход является очень мощным инструментом для демонстрации разнообразия. Поэтому он был выбран для этого сайта. В отличие от некоторых классических антропологов, которые продвигали свои выводы как определенные по разным причинам, здесь они рассматриваются просто как фенотипическая модель. |
 Интегральный подход также учитывает исторические факты, т.е. о старых миграциях, семейной истории человека и окружающей среде, в которой живет человек, потому что они могут изменить фенотипы. Это также имеет значение, если тип сохраняется в семьях, социальных группах или популяциях, например. семейный или деревенский. Только после учета всей возможной информации производится классификация.
При учете такого большого количества информации интегральный подход сталкивается с той же проблемой, что и численный подход, поскольку количество возможных комбинаций быстро увеличивается с каждым рассматриваемым признаком (см. рис. 1). Однако у интегрального подхода есть обходной путь, поскольку он ослабляет критерии сортировки. т.е. он определяет набор «идеальных» типов, но допускает отклонение небольшого числа признаков (например, двух или трех). Пример показан ниже на рисунке 2. Высокий брахицефальный темноволосый человек может быть норд(о)идом 9.0541 1 при интегральном подходе, даже если два антропометрических признака отклоняются, если остальные десятки признаков (и, например, семейный анамнез) являются типично нордскими.
Интегральный подход также учитывает исторические факты, т.е. о старых миграциях, семейной истории человека и окружающей среде, в которой живет человек, потому что они могут изменить фенотипы. Это также имеет значение, если тип сохраняется в семьях, социальных группах или популяциях, например. семейный или деревенский. Только после учета всей возможной информации производится классификация.
При учете такого большого количества информации интегральный подход сталкивается с той же проблемой, что и численный подход, поскольку количество возможных комбинаций быстро увеличивается с каждым рассматриваемым признаком (см. рис. 1). Однако у интегрального подхода есть обходной путь, поскольку он ослабляет критерии сортировки. т.е. он определяет набор «идеальных» типов, но допускает отклонение небольшого числа признаков (например, двух или трех). Пример показан ниже на рисунке 2. Высокий брахицефальный темноволосый человек может быть норд(о)идом 9.0541 1 при интегральном подходе, даже если два антропометрических признака отклоняются, если остальные десятки признаков (и, например, семейный анамнез) являются типично нордскими. Такой человек будет классифицирован как динарид при строгом числовом подходе, основанном на таблице 1, в которой используются три черты. На рисунке ясно видны значительные различия между Nordoid и Dinarid, хотя они были бы одинаковыми в приведенном выше примере численного подхода. Это связано с тем, что трех признаков обычно слишком мало, чтобы зафиксировать человеческий фенотип.
Некоторые могут возразить, что человек в середине рисунка 2 приближается к тому, что многие авторы называют альпинидами, но в интегральном подходе альпиниды определяются как невысокие и коренастые, с более круглым лбом и более слабым подбородком, меньшим черепом, более короткими ногами, более широким носом и т. д. Некоторые можно считать смешанным. Пример иллюстрирует, как разные типологические подходы могут приводить к разным результатам в их классификации из-за различий в методологии. Кроме того, для интегральных антропологов мягкие ткани обычно играли относительно большую роль (например, при дифференциации средиземноморцев от ориенталидов).
Такой человек будет классифицирован как динарид при строгом числовом подходе, основанном на таблице 1, в которой используются три черты. На рисунке ясно видны значительные различия между Nordoid и Dinarid, хотя они были бы одинаковыми в приведенном выше примере численного подхода. Это связано с тем, что трех признаков обычно слишком мало, чтобы зафиксировать человеческий фенотип.
Некоторые могут возразить, что человек в середине рисунка 2 приближается к тому, что многие авторы называют альпинидами, но в интегральном подходе альпиниды определяются как невысокие и коренастые, с более круглым лбом и более слабым подбородком, меньшим черепом, более короткими ногами, более широким носом и т. д. Некоторые можно считать смешанным. Пример иллюстрирует, как разные типологические подходы могут приводить к разным результатам в их классификации из-за различий в методологии. Кроме того, для интегральных антропологов мягкие ткани обычно играли относительно большую роль (например, при дифференциации средиземноморцев от ориенталидов).
 В целом число теоретически возможных определений типов, которые могут быть включены в типологию, равно 9.0350 почти бесконечное , хотя лишь немногие из потенциальных типологий действительно хорошо отражают человеческое разнообразие. Могут существовать аналогичные типологии аналогичного значения, которые также описывают разнообразие. Таким образом, определения типов интегральных подходов иногда различаются, особенно в деталях. Таким образом, любая типология больше похожа на модель, которая существует в большинстве наук в качестве полезных инструментов — как упрощение реальности, которое никогда не может охватить каждую деталь реального мира. Кроме того, впечатление, которое человек может произвести на человека-наблюдателя для качественной оценки, может быть предвзятым, поскольку такая оценка зависит от навыков, личности, мотивации и опыта наблюдателя. Другая проблема заключается в том, что исторические данные и антропометрические измерения могут содержать ошибки. В настоящее время можно было бы частично заменить информацию из исторических документов и сказок (например, о старых миграциях) генетической информацией и иметь больше и более качественные антропометрические данные.
В целом число теоретически возможных определений типов, которые могут быть включены в типологию, равно 9.0350 почти бесконечное , хотя лишь немногие из потенциальных типологий действительно хорошо отражают человеческое разнообразие. Могут существовать аналогичные типологии аналогичного значения, которые также описывают разнообразие. Таким образом, определения типов интегральных подходов иногда различаются, особенно в деталях. Таким образом, любая типология больше похожа на модель, которая существует в большинстве наук в качестве полезных инструментов — как упрощение реальности, которое никогда не может охватить каждую деталь реального мира. Кроме того, впечатление, которое человек может произвести на человека-наблюдателя для качественной оценки, может быть предвзятым, поскольку такая оценка зависит от навыков, личности, мотивации и опыта наблюдателя. Другая проблема заключается в том, что исторические данные и антропометрические измерения могут содержать ошибки. В настоящее время можно было бы частично заменить информацию из исторических документов и сказок (например, о старых миграциях) генетической информацией и иметь больше и более качественные антропометрические данные. Такая генетически расширенная интегральная типология может быть разработана путем максимизации группы 1 и минимизации группы 3 после сбора данных. В качестве альтернативы количество переменных может быть уменьшено с помощью анализа основных компонентов и аналогичных методов. Поскольку у классических антропологов такой информации не было, вполне возможно, что некоторые из их типов устарели бы при современном подходе.
Такая генетически расширенная интегральная типология может быть разработана путем максимизации группы 1 и минимизации группы 3 после сбора данных. В качестве альтернативы количество переменных может быть уменьшено с помощью анализа основных компонентов и аналогичных методов. Поскольку у классических антропологов такой информации не было, вполне возможно, что некоторые из их типов устарели бы при современном подходе.3.) Население
Проблемы неоднозначности других типологических подходов можно преодолеть с помощью популяционного подхода. Этот подход был особенно распространен во второй половине 20-го века и обычно использовался русской антропологической школой, а также некоторыми французскими, немецкими и другими исследователями. Вместо того, чтобы рассматривать отдельных лиц, популяционный подход рассматривает средние показатели комплексов населения или языковых групп. Эти группы могут содержать широкий спектр фенотипов. Явным преимуществом популяционного подхода является то, что популяции или языковые группы часто могут быть четко определены, следовательно, существует уникальное, четкое решение. Кроме того, его применимость очень высока. Может быть включено множество различных метрик, и для результирующей системы типов не существует ограничений, как в двух подходах, показанных выше. Теоретически каждый популяционный комплекс хорошо узнаваем даже при оценке 1000 антропометрических признаков. Первые компьютеры 20-го века могли обрабатывать усредненные показатели населения гораздо проще, чем отдельные наборы данных. Таким образом, он добавил много новых выводов в исследование. Образовавшиеся комплексы населения, т.е. Балтийский, Западно-Балтийский и т. д. обычно не идентичны типам других подходов, хотя могут быть и схожими. Вместо того, чтобы рассматривать отдельных лиц, популяционный подход рассматривает средние показатели комплексов населения или языковых групп. Эти группы могут содержать широкий спектр фенотипов. Явным преимуществом популяционного подхода является то, что популяции или языковые группы часто могут быть четко определены, следовательно, существует уникальное, четкое решение. Кроме того, его применимость очень высока. Может быть включено множество различных метрик, и для результирующей системы типов не существует ограничений, как в двух подходах, показанных выше. Теоретически каждый популяционный комплекс хорошо узнаваем даже при оценке 1000 антропометрических признаков. Первые компьютеры 20-го века могли обрабатывать усредненные показатели населения гораздо проще, чем отдельные наборы данных. Таким образом, он добавил много новых выводов в исследование. Образовавшиеся комплексы населения, т.е. Балтийский, Западно-Балтийский и т. д. обычно не идентичны типам других подходов, хотя могут быть и схожими. Обычно внутри каждого популяционного комплекса выделяют несколько типов. На рис. 3 приведен пример. В верхнем ряду показаны локальные разновидности, выделенные с помощью интегрального подхода, во втором ряду приведены примеры популяционных комплексов, которые эти типы могли образовать. Обычно внутри каждого популяционного комплекса выделяют несколько типов. На рис. 3 приведен пример. В верхнем ряду показаны локальные разновидности, выделенные с помощью интегрального подхода, во втором ряду приведены примеры популяционных комплексов, которые эти типы могли образовать.Рисунок 3: Соотношение популяционных комплексов и антропологических типов интегрального подхода: Рисунок 3 также показывает один недостаток популяционного подхода: все три изображенных комплекса выглядят одинаково. Это связано с тем, что по мере агрегирования разнообразия фенотипов внутри популяции может быть потеряно много информации. Популяции могут быть разнообразными, социальные группы или группы предков внутри популяции могут существенно различаться, и это не всегда известно исследователю. Определенная социальная группа может быть больше похожа на жителей другого региона, чем на сожительствующие социальные группы. Если взять только среднее значение и дисперсию нескольких показателей по популяциям, возможно, будет отброшено большое количество информации, доступной только на индивидуальном уровне. Рисунок 4: Функция плотности вероятности предполагаемого роста мужчин для места с нордами и лаппидами в Северной Скандинавии: |
 В качестве примера на рисунке 4 показана оценочная функция плотности вероятности одного антропометрического признака, мужского роста, для места в Северной Скандинавии, населенного нордскими и саамскими людьми. Нордиды в среднем значительно выше саамов. Таким образом, функция имеет два пика. Подобные явления обнаруживаются во многих местах мира и во многих различных антропометрических характеристиках. Эта информация не может быть получена комплексом населения, если различные группы внутри населения не разделены. Это особенно невыгодно в очень разных странах, таких как Индия, где кастовая система порождала большое разнообразие внешности.
В качестве примера на рисунке 4 показана оценочная функция плотности вероятности одного антропометрического признака, мужского роста, для места в Северной Скандинавии, населенного нордскими и саамскими людьми. Нордиды в среднем значительно выше саамов. Таким образом, функция имеет два пика. Подобные явления обнаруживаются во многих местах мира и во многих различных антропометрических характеристиках. Эта информация не может быть получена комплексом населения, если различные группы внутри населения не разделены. Это особенно невыгодно в очень разных странах, таких как Индия, где кастовая система порождала большое разнообразие внешности. Однако популяционный подход остается наиболее надежным в практическом смысле.
Однако популяционный подход остается наиболее надежным в практическом смысле.4.) Раса как социальный конструкт
Другой подход, очень распространенный сегодня, состоит в том, чтобы определить антропологические группы или «расы» как социальную конструкцию. Это имеет мало общего с тремя подходами, проиллюстрированными выше, и еще меньше с филогенетическими отношениями. Критерии классификации различаются от страны к стране и меняются со временем. Человек, считающийся «черным» в Соединенных Штатах, может не рассматриваться как черный в других частях мира, человек может считаться «белым» в Уганде или даже в Австралии. В подходе социальных конструктов классификация людей часто связана с историей страны. Типичными примерами являются «правила одной капли». Например. если американец имеет 1/8 африканского происхождения, он будет классифицирован как афроамериканец, даже если фенотипически и генетически он может быть в основном европейцем. И наоборот, человек с 1/8 европейского происхождения не будет считаться в этой системе «белым» или «европейцем». Большую роль играет и то, как человек себя ведет, где он вырос и как себя идентифицирует. У некоторых людей может быть происхождение, при котором они могут выбирать свою «расу», а у людей с другим происхождением — нет. Этот подход также применялся к коренным американцам в США, коренным австралийцам или даже айнам в Японии. Другой характеристикой подхода социальных конструктов является чрезмерный акцент на единственной антропометрической характеристике, часто на цвете кожи. Например. Суданиды. Эфиопидов, индо-меланидов, негритидов и австралидов можно охарактеризовать просто как «черных», хотя они сильно различаются генетически, исторически и даже антропометрически. У них просто есть хотя бы одна общая антропометрическая черта. Эквивалентным было бы объединение всех широколицых людей в одну «расу» (чего никогда не делалось). В некоторых случаях политические идеологии играют большую роль в подходе социальных конструктов, несколько десятилетий назад аналогичные методы в антисемитских странах Европы использовались для идентификации евреев. И наоборот, человек с 1/8 европейского происхождения не будет считаться в этой системе «белым» или «европейцем». Большую роль играет и то, как человек себя ведет, где он вырос и как себя идентифицирует. У некоторых людей может быть происхождение, при котором они могут выбирать свою «расу», а у людей с другим происхождением — нет. Этот подход также применялся к коренным американцам в США, коренным австралийцам или даже айнам в Японии. Другой характеристикой подхода социальных конструктов является чрезмерный акцент на единственной антропометрической характеристике, часто на цвете кожи. Например. Суданиды. Эфиопидов, индо-меланидов, негритидов и австралидов можно охарактеризовать просто как «черных», хотя они сильно различаются генетически, исторически и даже антропометрически. У них просто есть хотя бы одна общая антропометрическая черта. Эквивалентным было бы объединение всех широколицых людей в одну «расу» (чего никогда не делалось). В некоторых случаях политические идеологии играют большую роль в подходе социальных конструктов, несколько десятилетий назад аналогичные методы в антисемитских странах Европы использовались для идентификации евреев. Подход социальных конструктов не используется на этом сайте, потому что он не может описать биологическое разнообразие человека научным способом. Подход социальных конструктов не используется на этом сайте, потому что он не может описать биологическое разнообразие человека научным способом. |
5.) Филогенетический подход
Очень мощным научным подходом к изучению человеческого разнообразия является филогенетический подход. Вместо рассмотрения фенотипических и антропометрических признаков он изучает генетический состав людей, часто на основе данных молекулярного секвенирования. Термин «филогенетика» происходит от греческих терминов «фил» и «филон», обозначающих «племя», «клан», «расу». С развитием генетических методов в конце 20-го и начале 21-го века этот подход стал распространенным методом исследования биологического разнообразия человека, хотя 100 лет назад уже существовали филогенетические деревья, которые имели сходство с современными деревьями. Как правило, разнообразие генетических признаков оценивается, а затем с помощью статистического анализа объединяется в основные компоненты или аналогичные факторы. Филогенетический подход лучше всего подходит для изучения древних миграций, времени расхождения человеческих популяций и степени их смешения. Однако он сталкивается с теми же статистическими проблемами, что и численный подход, когда речь идет о сопоставимости и качестве данных. В большинстве случаев собранная генетическая информация будет описывать только часть всего биологического организма, данные могут содержать ошибки, а методы измерения не всегда сопоставимы. Это может привести к необъективным результатам, поскольку сложный организм не рассматривается полностью, а многие ненаблюдаемые характеристики и гены игнорируются. Кроме того, применяемые статистические методы имеют ограничения и особые требования к данным, которые не всегда могут быть удовлетворены. Другая слабость заключается в том, что филогенетический подход не учитывает фенотипические различия (см. ниже). Однако, несмотря на эти недостатки, возможность использования филогенетического подхода для изучения древних миграций человека остается большой. Филогенетический подход лучше всего подходит для изучения древних миграций, времени расхождения человеческих популяций и степени их смешения. Однако он сталкивается с теми же статистическими проблемами, что и численный подход, когда речь идет о сопоставимости и качестве данных. В большинстве случаев собранная генетическая информация будет описывать только часть всего биологического организма, данные могут содержать ошибки, а методы измерения не всегда сопоставимы. Это может привести к необъективным результатам, поскольку сложный организм не рассматривается полностью, а многие ненаблюдаемые характеристики и гены игнорируются. Кроме того, применяемые статистические методы имеют ограничения и особые требования к данным, которые не всегда могут быть удовлетворены. Другая слабость заключается в том, что филогенетический подход не учитывает фенотипические различия (см. ниже). Однако, несмотря на эти недостатки, возможность использования филогенетического подхода для изучения древних миграций человека остается большой. На рис. 5 показано сильно упрощенное филогенетическое древо современного человека. На рис. 5 показано сильно упрощенное филогенетическое древо современного человека. |
Филогенетическое древо показывает общего предка Homo sapiens и неандертальцев, живших около полумиллиона лет назад в Африке. Неандертальцы, покинувшие Африку за несколько сотен тысяч лет до Homo sapiens, приспособились к прохладному климату ледникового периода Европы и Азии, развив коренастое тело, очень большие носы и более светлую кожу. Homo sapiens начал расходиться на современные разновидности около 150 000 лет до н. э. в Африке. Ранние Homo sapiens, вероятно, жили в сухом тропическом климате Африки (во время ледникового периода тропики были менее влажными, чем сегодня). Таким образом, носы, вероятно, были лишь умеренно широкими (см. взаимосвязь между шириной носа и влажностью), а кожа, вероятно, была (темно) коричневой. Койсан отделился первым от всех остальных людей. Общий предок всех некойсанских групп развил разные разновидности: протосуданиды, протонилотиды и протоэтиопиды. Последняя группа начала мигрировать из Африки около 60 000 лет назад и смешалась с неандертальцами и, возможно, другими людьми, такими как денисовцы. Когда люди мигрировали на север, их кожа стала светлее, часто достигая светлого или среднего коричневого тона. Фенотипы быстро начали меняться в новых условиях из-за адаптивного давления, естественного отбора и культурных изменений, которые привнесли новые условия. Северо-восточные азиаты приспособились к континентальному климату с ледяными зимами, европейцы к земледельческой жизни в облачном умеренном климате. Обе адаптации привели от скромной к сильной депигментации, более узким носам и более коротким конечностям. Некоторые группы мигрировали через Южную Азию в Австралию и Меланезию (например, предки букаидов). Хотя букаиды генетически ближе к европейцам и выходцам из Восточной Азии, они выглядят относительно похожими на бантуидов и многих других африканцев к югу от Сахары из-за их адаптации к аналогичному влажному и тропическому климату. Койсан отделился первым от всех остальных людей. Общий предок всех некойсанских групп развил разные разновидности: протосуданиды, протонилотиды и протоэтиопиды. Последняя группа начала мигрировать из Африки около 60 000 лет назад и смешалась с неандертальцами и, возможно, другими людьми, такими как денисовцы. Когда люди мигрировали на север, их кожа стала светлее, часто достигая светлого или среднего коричневого тона. Фенотипы быстро начали меняться в новых условиях из-за адаптивного давления, естественного отбора и культурных изменений, которые привнесли новые условия. Северо-восточные азиаты приспособились к континентальному климату с ледяными зимами, европейцы к земледельческой жизни в облачном умеренном климате. Обе адаптации привели от скромной к сильной депигментации, более узким носам и более коротким конечностям. Некоторые группы мигрировали через Южную Азию в Австралию и Меланезию (например, предки букаидов). Хотя букаиды генетически ближе к европейцам и выходцам из Восточной Азии, они выглядят относительно похожими на бантуидов и многих других африканцев к югу от Сахары из-за их адаптации к аналогичному влажному и тропическому климату. Рисунок 5 показывает, что при относительно постоянных условиях в доисторической Африке фенотипы менялись с умеренной скоростью, в то время как они быстро менялись в различных новых условиях за пределами Африки. Рисунок 5 показывает, что при относительно постоянных условиях в доисторической Африке фенотипы менялись с умеренной скоростью, в то время как они быстро менялись в различных новых условиях за пределами Африки.По крайней мере два разных процесса определяют человеческое разнообразие: генетическая дивергенция и фенотипическая дивергенция. Основной движущей силой генетической дивергенции являются стохастические генетические мутации, которые происходят с течением времени, в то время как основной движущей силой фенотипической дивергенции является адаптивное давление в новых условиях, культуре и возникающих в результате процессах естественного отбора. Разделенные популяции при сходных обстоятельствах долгое время будут оставаться похожими из-за одинакового адаптивного давления и, возможно, даже конвергентной эволюции. Для различий между людьми важны оба процесса. Это показано ниже. |
Почему важны фенотипы?
В предыдущем разделе было показано, что филогенетический подход очень полезен для изучения человеческих миграций и генетической дивергенции. Однако различия у людей обусловлены не только генетической дивергенцией, но и фенотипической дивергенцией, вызванной адаптивным давлением. В ходе эволюции новый вид может быть создан любым из этих процессов, в большинстве случаев комбинацией обоих. Если не будет фенотипической дивергенции, а будет длительный процесс генетической дивергенции, в разных популяциях будет развиваться слишком много генетических различий, чтобы продолжать производить жизнеспособное потомство и развиваться в новые виды. Если не будет генетической дивергенции, а будет сильный естественный отбор и адаптивное давление на одну или обе популяции, их анатомия станет настолько другой, что они также перестанут скрещиваться. До видообразования обычно развиваются разные подвиды или «расы». Не существует единого определения, с какого момента разные фенотипы и/или генотипы могут рассматриваться как разные подвиды. Это иллюстрируется «серой зоной» на рис. 6, которая изображает два процесса (гораздо более подробное описание фенотипической и генетической дивергенции в эволюции см. Однако различия у людей обусловлены не только генетической дивергенцией, но и фенотипической дивергенцией, вызванной адаптивным давлением. В ходе эволюции новый вид может быть создан любым из этих процессов, в большинстве случаев комбинацией обоих. Если не будет фенотипической дивергенции, а будет длительный процесс генетической дивергенции, в разных популяциях будет развиваться слишком много генетических различий, чтобы продолжать производить жизнеспособное потомство и развиваться в новые виды. Если не будет генетической дивергенции, а будет сильный естественный отбор и адаптивное давление на одну или обе популяции, их анатомия станет настолько другой, что они также перестанут скрещиваться. До видообразования обычно развиваются разные подвиды или «расы». Не существует единого определения, с какого момента разные фенотипы и/или генотипы могут рассматриваться как разные подвиды. Это иллюстрируется «серой зоной» на рис. 6, которая изображает два процесса (гораздо более подробное описание фенотипической и генетической дивергенции в эволюции см. , например, Winker, 2009).). , например, Winker, 2009).). Рисунок 6: Фенотипическая и генетическая дивергенция: Рисунок 7: Фенотипическая и генетическая дивергенция для выбранных пар типов с рисунка 5: Таким образом, фенотипические, а также генетические различия объясняют разнообразие людей. Человеческое разнообразие может иметь значение для здоровья, питания, общества, спорта и навыков. Текущее исследование добавляет много информации о генетических различиях, но мало о фенотипических различиях. Однако на протяжении большей части палеолита, когда основные современные ветви Homo sapiens расходились, люди были тесно связаны с климатом и природой, в которых они жили, их фенотипы были зеркалом окружающей их среды. Таким образом, Humanphenotypes пытается закрыть этот пробел, предоставляя типологическую модель, которая обобщает доступную информацию о фенотипических различиях между людьми. Данные и типы в основном изучались в течение 20 века. About the AuthorRelated Posts |
 рис. 5), но все же оба типа похожи друг на друга. Это связано с тем, что оба живут в относительно похожем климате, и адаптивные нагрузки были одинаковыми. Таким образом, хотя генетические различия между букаидами и бантуидами больше, фенотипическая разница невелика.
рис. 5), но все же оба типа похожи друг на друга. Это связано с тем, что оба живут в относительно похожем климате, и адаптивные нагрузки были одинаковыми. Таким образом, хотя генетические различия между букаидами и бантуидами больше, фенотипическая разница невелика.