самоорганизация и целенаправленное поведение / Хабр
Введение
В данном топике я хочу рассказать о модели необычной нейронной сети, в разработке которой мне посчастливилось участвовать. Данная модель была разработана около года назад (про авторство написано в последнем разделе), однако исследование ее после этого прекратились из-за совершенной нехватки времени (занятости в своих собственных проектах). Тем не менее я опишу ее здесь в надежде на то, что некоторые мысли покажутся читателям интересными и дадут ростки возможных дальнейших исследований в данном направлении.
Сразу оговорюсь, что эта модель не претендует никоим образом на прототип ИИ. Мы скорее хотели исследовать возможность самоорганизации и перспективы возникновения целостного целенаправленного поведения в динамической системе «эгоистичных» (гомеостатических) нейронов.
Как мне кажется, для более полного понимания логики работы модели будет полезно прочитать мой топик про Теорию функциональных систем, однако опять же это на ваше усмотрение.
Поехали!
Биологическое основание
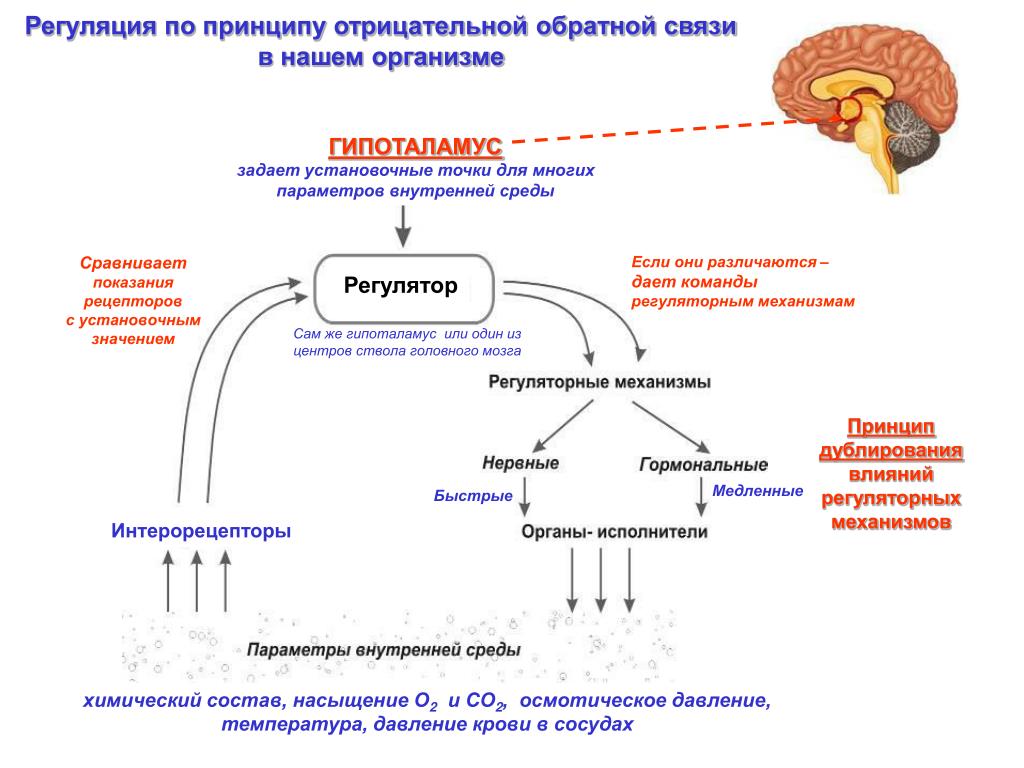
Данная модель строилось на основании теории функциональных систем П.К. Анохина и теории гомеостаза.
Гомеостаз — саморегуляция, способность открытой системы сохранять постоянство своего внутреннего состояния посредством скоординированных реакций, направленных на поддержание динамического равновесия. Уолтер Кэнон, который изначально ввел этот термин, называл гомеостаз «мудростью тела» [1].
Простейшие биологические мотивации возникают при нарушении гомеостатического равновесия в нейронах соответствующих физиологических зон (нарушение запасов энергии, кислорода или осмотический дисбаланс). Тем не менее, существует некая универсальная единица для оценки биологической мотивации [2]. Существование этой единицы связано с тем, что удовлетворение любой мотивации вызывает удовольствие и, соответственно, удаление дисбаланса в гомеостатической системе, который и порождает данную мотивацию. Поэтому моделирование гомеостаза – это прямой путь к созданию искусственной системы, обладающей некоторым подобием мотивации, т.

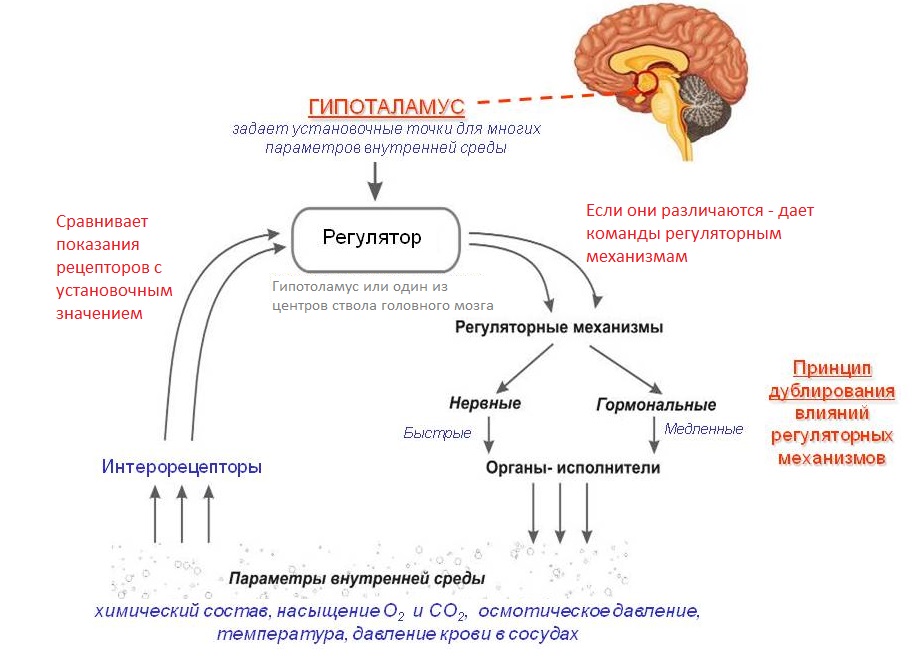
Рис. 1. Гомеостаз нейрона
(LoR – гомеостаз низкого уровня, HoR – гомеостаз высокого уровня, LS – локальный сенсор рассогласования, R – награда снижающая уровень рассогласования)
В литературе известен набор моделей гомеостаза [3,4], но они не ориентированы на самоорганизацию нейронных сетей и целенаправленное поведение. Моделирование целенаправленного поведения, в том числе на основе нейронных сетей и обучения с подкреплением, также имеет богатую историю и широко освещена в литературе. Однако в рамках этих моделей не рассматривался гомеостаз нейронов и сетей в целом.
Собственно, исходя из всего вышесказанного, была сформулировано иерархия моделей, которые составят полную систему:
— модель гомеостатического нейрона, объединяющая ключевые характеристики известных моделей и основные принципы рассматриваемой парадигмы;
— модели гомеостатической нейронной сети, основанной на гомеостатических нейронах;
— модель простейшего организма, управляемого гомеостатической нейронной сетью, и его взаимодействия со средой;
— моделирование эволюционного развития простейших организмов в условиях изменяющейся внешней среды.
Модель гомеостатического нейрона и нейронной сети
Так как основной задачей является анализ парадигмы гомеостатического нейрона как возможной основы механизма самоорганизации нейронной сети и ее способности к автономному поведению, то особенное внимание необходимо уделить именно этой ключевой первичной модели. Модель гомеостатического нейрона подразумевает, что каждый нейрон стремится к поддержанию своего оптимального внутреннего состояния – гомеостаза, причем при рассогласовании текущего состояния с оптимальным, нейрон формирует ответ, направленный на возвращение к оптимальному состоянию. Как уже упоминалось ранее изменение множества факторов может привести к нарушению равновесия нейрона, однако в модельной ситуации введем некоторую единую

Стремление нейрона к гомеостазу выражается в активности, направленной на устранение рассогласования
qopt-q(t), тем более выраженной, чем больше величина рассогласования. Активность нейрона требует энергетических затрат, поэтому может принимать разные формы в зависимости от количества имеющейся энергии e(t).Таким образом, мы сформулировали некоторые требования к модели «эгоистического» нейрона, т.е. такого нейрона который имеет одну цель – поддерживать свое состояние близкое к оптимальному. Однако, каждый нейрон не может существовать изолировано, а обязан функционировать в целостной нейронной сети. Таким образом, мы можем ввести понятие вектора воздействия на конкретный нейрон со стороны остальной сети:
Это воздействие меняет эндогенную оценку нейрона на следующий такт времени:
Данная функция может выглядеть по разному, однако самый просто ее случай – это
Получается, что выше мы ввели все характеристики нейрона, а также методику определения воздейтсвия на нейрон со стороны остальных нейронов.
Теперь определим действие нейрона, которое он выбирает на основе текущего рассогласования:
На этом этапе необходимо остановится подробнее, так как с одной стороны он является одним из ключевых во всей модели, а с другой стороны само понятие действия аккумулирует в себе несколько биологических принципов и наших размышлений, о которых было сказано выше.
Первое, что необходимо заметить – это то, что любое воздействие на нейрон со стороны других нейронов влечет к отклонению от его гомеостатического равновесию. Соответственно все действия нашего «эгоистичного» нейрона должны быть направлены на возвращение к этому равновесию. В этом заключается
принцип гомеостаза низкого уровня. Тут возможны несколько случаев, которые собственно и были рассмотрены при составлении функции действия.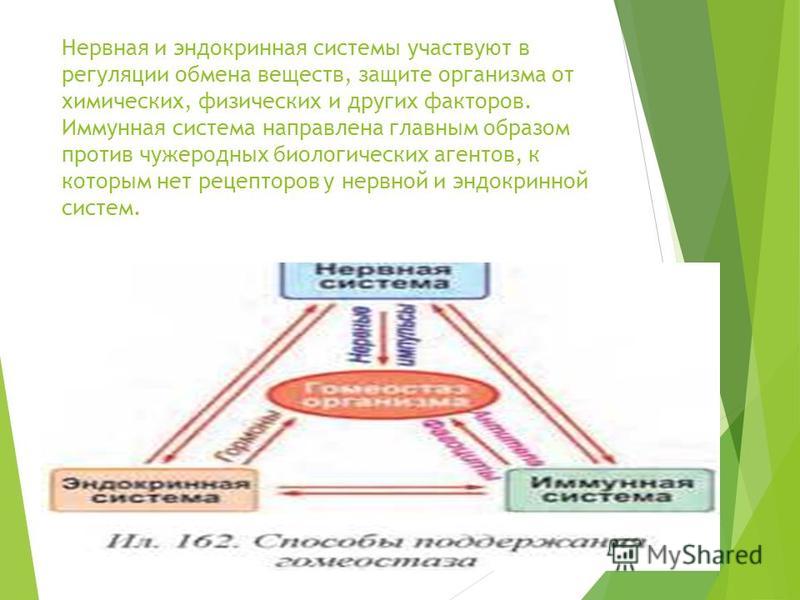
Действие нейрона в первую очередь зависит от степени его рассогласованности. Если нейрон находится далеко от точки гомеостатического равновесия, то он осуществляет действие, которое немедленно возвращает его к равновесию – генерацию спайка. Если же он близок к равновесию, то включается другой механизм гомеостаза – медленное восстановление. На каждое действие нейрон тратит энергию: на спайк – большую, а на восстановление – не очень большую. Про пополнение энергии нейрона будет рассказано далее.
Кроме этого энергии нейрона может не хватить на генерацию спайка, даже когда он необходим, что означает выбор восстановление до того момента, пока энергия не пополнится.
Таким образом, действие нейрона является функцией от входного сигнала, эндогенной оценки состояния и запаса энергии. Собственно на этом заканчивается формальная модель нейрона (в том числе гомеостаза нижнего уровня) и сети в целом. На рис. 2 приведено схематическое изображение «эгоистического» нейрона.
Рис. 2. «Эгоистичный» нейрон
2. «Эгоистичный» нейрон
Модель организма
Нейронная сеть является частью целого организма, однако для его полного определения необходимо ввести еще несколько дополнительных понятий. Первое из них – это общий пул энергии Е(t), из которого восстанавливаются энергии всех нейронов сети. Если e(t)<emin (оптимальный запас энергии нейрона), то
Теперь нам необходимо определить понятие гомеостаза высшего уровня.
Введем понятие эндогенной оценки состояния всей сети:
При это будем считать, что есть некоторое оптимальное состояние Qopt:
Тогда определимся, что гомеостаз высшего уровня вступает в действие в двух различных случаях:
— после установления стабильности (в случае если в сети наблюдается циклический процесс, но состояние не достигает оптимального значения)
— при «возникновении потребности» (в случае большого рассогласования эндогенной оценки состояния сети)
В этом случае происходит изменение параметров qiopt случайным образом, при этом величина изменения каждого значения пропорциональна, как рассогласованию эндогенной оценки состояния сети, так и рассогласованию эндогенной оценки конкретного нейрона. Таким образом, при невозможности системы достичь оптимального состояния и при угрозе жизни всей сети, включается механизм гомеостаза высшего уровня. Cхематично такая модель организма показана на рис. 3.
Таким образом, при невозможности системы достичь оптимального состояния и при угрозе жизни всей сети, включается механизм гомеостаза высшего уровня. Cхематично такая модель организма показана на рис. 3.
Рис. 3. Модель организма
Здесь, наверное необходимо привести некоторые экспериментальные данные, по которым можно будет судить о качественной динамики такой модели (без гомеостаза высшего уровня). Мы составили сеть из 10 нейронов со случайно распределенными весами по распределению Wij~N(0,1) и с различными оптимальными значениями эндогенной оценки состояния и запаса энергии. При этом на момент инициализации 9 нейронов находились в состоянии равновесия, а один из нейронов был рассогласован. На рис. 4-5 приведены зависимости от времени эндогенной оценки состояния и энергии каждого нейрона в такой системе.
Рис. 4. График динамики эндогенной оценки состояния нейронов
Рис. 5. График динамики запаса энергии нейронов
По рисункам видно, что в итоге вся система приходит в положение гомеостатического равновесия, однако наши опыты показывают, что существует область параметров, при которых достижение гомеостатического равновесия невозможно и в системе начинается автоколебательный процесс, что свидетельствует о необходимости включения гомеостаза высшего уровня.
Взаимодействие со средой
Представим себе, что мы помещаем такой организм в среду. Предположим, что у нашего организма всего одна потребность – потребность в поддержании достаточного уровня общей энергии, которая расходуется на гомеостаз и на метаболические процессы. В таком случае необходимо, чтобы организм мог принимать решение в нужный момент о восполнении уровня энергии. То есть, например нажимать на кнопку, чтобы ему подавалась еда.
Мы думали о введение макропараметров организма, изменение которых определяет его потребности (их может быть несколько), а также о рассмотрение набора эффекторов, позволяющих организму изменять значения своих макропараметров за счет взаимодействия с внешней средой. В основе предлагаемого подхода к проблеме лежит передача информации о значениях макропараметров непосредственно гомеостатическим нейронам, что связывает внутренний гомеостаз нейрона с удовлетворением соответствующей потребности. Таким образом, гомеостаз второго уровня отвечает за работоспособность системы в целом, а первого уровня – за эффективную линию поведения организма при взаимодействии с окружающей средой.
Поясню на примере. Для этого предположим, что в структуре нейронной сети присутствует два специализированных нейрона.
Первый нейрон специализирован относительно потребности в восполнении энергии, то есть он вступает в рассогласование в случае, если уровень общей энергии недостаточен. То есть у него есть дополнительный вход, на который поступает уровень недостатка общей энергии ( E-Emin). Второй нейрон специализирован относительно выполнения действия восполнения энергии, то есть в тот момент, когда он генерирует спайк происходит восполнение общей энергии из внешней среды. Данная система изображена на рис. 6.
Рис. 6. Взаимодействие организма со средой
красный нейрон специализирован относительно потребности в восполнении энергии, синий — относительно выполнения действия восполнения энергии)
Таким образом, рассогласование в организм вносится в тот момент, когда он испытывает потребность в еде. В свою очередь нейроны пытаются устранить это рассогласование.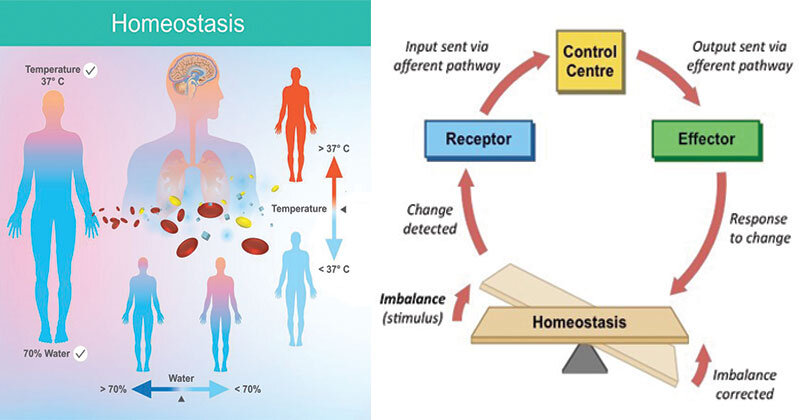 Пока не будет наблюдаться спайк на нейроне специализированном относительно выполнения действия, восполнение энергии не произойдет, и сигнал о недостатке энергии будет продолжать поступать в систему. Таким образом, в какой-то момент спайк генерируется на втором специализированном нейроне и запас общей энергии пополняется, из-за чего нейрон, который специализирован относительно потребности в восполнении энергии перестает быть рассогласованным.
Пока не будет наблюдаться спайк на нейроне специализированном относительно выполнения действия, восполнение энергии не произойдет, и сигнал о недостатке энергии будет продолжать поступать в систему. Таким образом, в какой-то момент спайк генерируется на втором специализированном нейроне и запас общей энергии пополняется, из-за чего нейрон, который специализирован относительно потребности в восполнении энергии перестает быть рассогласованным.
Безусловно это очень модельная ситуация, однако она показывает по какой логике может функционировать такой организм в среде, где требуется принимать некоторые решения.
Непрерывная модель
Исключительно для тех, кто задумается о том, как можно эту дискретную во времени модель сделать непрерывной, привожу наш вариант, основанный на модели нейронов ФитцХью-Нагумо.
Заключение
Безусловно данная модель не может претендовать на хоть какую-то законченность. Однако, мне кажется, что некоторые мысли в нее заложенные, достаточно интересны и могут стать объектом исследования читателей.
 В частности идея рассогласования нейрона, как мотивации для совершения действия, определение рассогласования – как соотношения внутренних параметров нейрона с микросредой, а также гомеостаз – как основа для устранения рассогласования и для самоорганизуещего эффекта.
В частности идея рассогласования нейрона, как мотивации для совершения действия, определение рассогласования – как соотношения внутренних параметров нейрона с микросредой, а также гомеостаз – как основа для устранения рассогласования и для самоорганизуещего эффекта.Также этим топиком я хотел обратить внимание читателей на то, что даже в достаточно простых моделях могут возникать интересные эффекты самоорганизации, которые в перспективе могут приводит к целостному поведению.
Отмечу, что в данном обзоре я не касался нетривиального вопроса введения процесса обучения в эту модель, а также собственно эволюции. Все это остается предметом дальнейших возможных исследований.
Авторство
В разработке данной модели принимали участие Максим Комаров (Нижегородский университет), Даниил Каневский (ВМиК МГУ), Сергей Куливец (ИПУ РАН), а также ваш покорный слуга. Также огромной благодарности заслуживают Лев Ефимович Цитоловский (Бар-Иланский Университет, Израиль) и Владимир Георгиевич Редько (НИИСИ РАН) за их поддержку и слежение за направлением наших мыслей, когда нас заносило.
 Каждый из нас привнес в эту модель свои мысли и соображения из областей, которыми мы занимаемся.
Каждый из нас привнес в эту модель свои мысли и соображения из областей, которыми мы занимаемся.Список литературы
[1]. Cannon W. B. Physical Review, 1928, 9:399-431.
[2]. Cabanac M. Journal of Theoretical Biology, 1992, 155:173-200.
[3]. Davis G.W. Annual Review of Neuroscience, 29:307-323, 2006.
[4]. Marder A., Prinz A. A. BioEssays, 24: 1145-1154, 2002
Гомеостаз
Гомеостаз на уровне организма или клетки — это частный случай наблюдаемого в природных открытых системах поддержания стационарного состояния. В простых физико-химических системах стационарное состояние определяется постоянством внешних условий. Биологические системы способны активно противостоять изменениям окружающей среды и стационарное состояние у них — следствие функционирования механизмов регуляции.[ …]
Гомеостаз на уровне экосистем. Многообразные формы связей, существующие между популяциями разных видов, объединяют биоценоз в целостную биологическую систему. Как все биологические системы любого уровня, биоценоз существует в определенных абиотических условиях того или иного участка географической среды. Эта среда никогда не является абсолютно стабильной, а устойчивое выполнение главенствующей функции экосистем — поддержания биогенного круговорота — должно обеспечиваться биоценотическими адаптивными механизмами. Эти механизмы при рассмотрении их на уровне экосистем носят исторический характер, так как складываются на протяжении всего существования и формирования экосистем. Между элементами экосистемы устанавливаются общие стабильные взаимоотношения, которые адаптированы не только к общим факторам существования данной экосистемы, но и к степени взаимосвязей, определяющих устойчивое поддержание этих отношений в колеблющихся условиях данной конкретной среды.[ …]
Как все биологические системы любого уровня, биоценоз существует в определенных абиотических условиях того или иного участка географической среды. Эта среда никогда не является абсолютно стабильной, а устойчивое выполнение главенствующей функции экосистем — поддержания биогенного круговорота — должно обеспечиваться биоценотическими адаптивными механизмами. Эти механизмы при рассмотрении их на уровне экосистем носят исторический характер, так как складываются на протяжении всего существования и формирования экосистем. Между элементами экосистемы устанавливаются общие стабильные взаимоотношения, которые адаптированы не только к общим факторам существования данной экосистемы, но и к степени взаимосвязей, определяющих устойчивое поддержание этих отношений в колеблющихся условиях данной конкретной среды.[ …]
Гомеостаз популяции — поддержание определенной численности (плотности). Изменение численности зависит от целого ряда факторов среды — абиотических, биотических и антропогенных. Однако всегда можно выделить ключевой фактор, наиболее сильно влияющий на рождаемость, смертность, миграцию особей и т. д. На рис. 15 представлена схема поддержания гомеостаза популяции животных, где ключевым регулирующим факторов является доступность пищевых ресурсов. При повышении плотности популяции усиливаются механизмы, снижающие ее плотность, и наоборот.[ …]
д. На рис. 15 представлена схема поддержания гомеостаза популяции животных, где ключевым регулирующим факторов является доступность пищевых ресурсов. При повышении плотности популяции усиливаются механизмы, снижающие ее плотность, и наоборот.[ …]
Гомеостаз организма, материальные потоки в котором управляются когерентными электромагнитными полями ферми-систем надмолекулярных структур, определяется их информационным взаимодействием с окружающим волновым пространством. Физической аналогией информационных процессов в белковых структурах может быть резонансный электромагнитный контур, изменения добротности и сдвиги частоты которого, происходящие под воздействием различных факторов, качественно описывают процессы активизации тех или иных генов белковых структур. По этой причине все виды внешних воздействий, включая космические и социальные, вносящие десинхронизацию приобретенных при рождении организма информационных кодов, являются для него деструктивными.[ …]
Гомеостаз (ис) — способность организма или системы организмов поддерживать устойчивое (динамическое) равновесие в изменяющихся условиях среды. [ …]
[ …]
Гомеостаз — способность биологических систем — организма, популяции и экосистем — противостоять изменениям и сохранять равновесие. Исходя из кибернетической природы экосистем — гомеостатический механизм — это обратная связь. Например, у пойкилотермных животных изменение температуры тела регулируется специальным центром в мозге, куда постоянно поступает сигнал обратной связи, содержащий данные об отклонении от нормы, а от центра поступает сигнал, возвращающий температуру к норме. В механических системах аналогичный механизм называют сервомеханизмом, например, термостат управляет печью.[ …]
Гомеостаз обеспечивается работой механизмов регулирования, действующих по принципу отрицательной обратной связи. Тогда нарушения в функционировании живой системы, используя кибернетические термины, следует констатировать как появление в канале обратной связи «помех» или «шумов».[ …]
Гомеостаз поддержание относительного постоянства внутренних свойств, состава, функций внутренней среды биологической системы, организма при изменяющихся внешних условиях. [ …]
[ …]
Понятие «гомеостаз» широко используется в экологии для характеристики устойчивости различных систем. Гомеостаз клетки определяется специфическими физико-химическими условиями, отличными от условий внешней среды; гомеостаз многоклеточного организма — поддержанием постоянства внутренней среды. Константами гомеостаза животных являются объем, состав крови и других жидкостей организма.[ …]
По уровню гомеостаза предлагают судить о потенциальной устойчивости организма и клетки. Поэтому малость отклонения в живой системе по сравнению с величиной воздействия его вызывающего должна свидетельствовать о большей устойчивости системы (Левич, 1976). По нашему мнению, в последнем случае правильнее было бы говорить не об устойчивости живой системы, а о ее «робастности» (robust), т.е. об устойчивости к возмущениям. Явление биологической устойчивости много сложнее понятия стабильности гомеостатического состояния. Например, «реакция сверхчувствительности» — важный компонент устойчивости у растения, обеспечивает его выживаемость именно из-за высокой реактивности клеток организма к внешнему воздействию. [ …]
[ …]
Понятие о гомеостазе существует с древних времен, но оно больше известно под названием «равновесие в природе». Как отмечает П. Митчел (2001), в XX в. взгляды на равновесие в природе изменялись от полного их отрицания и до того, что виды сами поддерживают равновесие «ради своего блага». Последняя идея породила теорию о том, что все сообщества и даже Земля (Гея) представляют собой сверхорганизмы. Ч. Эльтон считал, что равновесие в природе не существует. И, на самом деле, оно не есть навсегда данное состояние, а изменяется, что и определяет ход эволюции биосферы.[ …]
Поддержание гомеостаза экосистемы возможно лишь в определенных пределах. Вне сферы действия отрицательной обратной связи вступает в силу положительная обратная связь. Область действия отрицательной обратной связи можно изобразить в виде гомеостатического плато (рис. 8.6). Оно состоит из ступенек; в пределах каждой ступеньки действует отрицательная обратная связь. Переход со ступеньки на ступеньку может произойти в результате изменения в «датчике». Так, увеличение или уменьшение количества пищевых ресурсов переводит гомеостаз на другой уровень.[ …]
Так, увеличение или уменьшение количества пищевых ресурсов переводит гомеостаз на другой уровень.[ …]
Считается, что гомеостаз является механизмом, регулирующим, упорядочивающим изменение свойств экосистемы. Постулируется, что совокупность оценок, характеризующих процессы, регулируемые в системе гомеостатическими механизмами, при нормальном состоянии подчиняется гауссовскому распределению [48]. Такое положение, подтвержденное экспериментально, дает возможность отделить нормальные состояния от состояний, выходящих за пределы нормы. Например, оценки, укладывающиеся при этом в интервал у±а, свидетельствуют о хорошем состоянии системы, а выходящие за эти пределы — о плохом.[ …]
На уровне экосистем гомеостаз проявляется в наиболее устойчивых формах взаимодействия между видами, что выражается в приспособленности к особенностям среды и поддержании циклов круговорота биогенов. Можно рассматривать даже гомеостаз биосферы, в которой взаимодействие разнообразных организмов поддерживает постоянство газового состава атмосферы, состав почв, состава и концентрации солей мирового океана и др. [ …]
[ …]
Таким образом, всеобщий гомеостаз биосферы зависит от стабильности биогеохимического круговорота веществ в природе. Но являясь планетарной экосистемой, она состоит из экосистем всех уровней, поэтому первоочередное значение для ее гомеостаза имеют целостность и устойчивость природных экосистем.[ …]
Однако обеспечение дыхательного гомеостаза является важнейшей, но не единственной функцией жабр. Наряду с почками жабры играют активную роль в поддержании водно-солевого гомеостаза, обеспечивая ионное и осмотическое постоянство внутренней среды [39, 140]. Обитая в резко различающихся по концентрации неорганических ионов морских и пресных водах, рыбы вынуждены с помощью различных морфофизиологических и физиолого-биохимических механизмов постоянно решать проблему стабилизации химического состава внутренней среды, т. е. изолировать внутриклеточные и внеклеточные жидкости от прямого влияния повышенной, или пониженной, солености окружающей среды. Основой решения этой проблемы служит непроницаемость для воды и ионов поверхность тела рыб.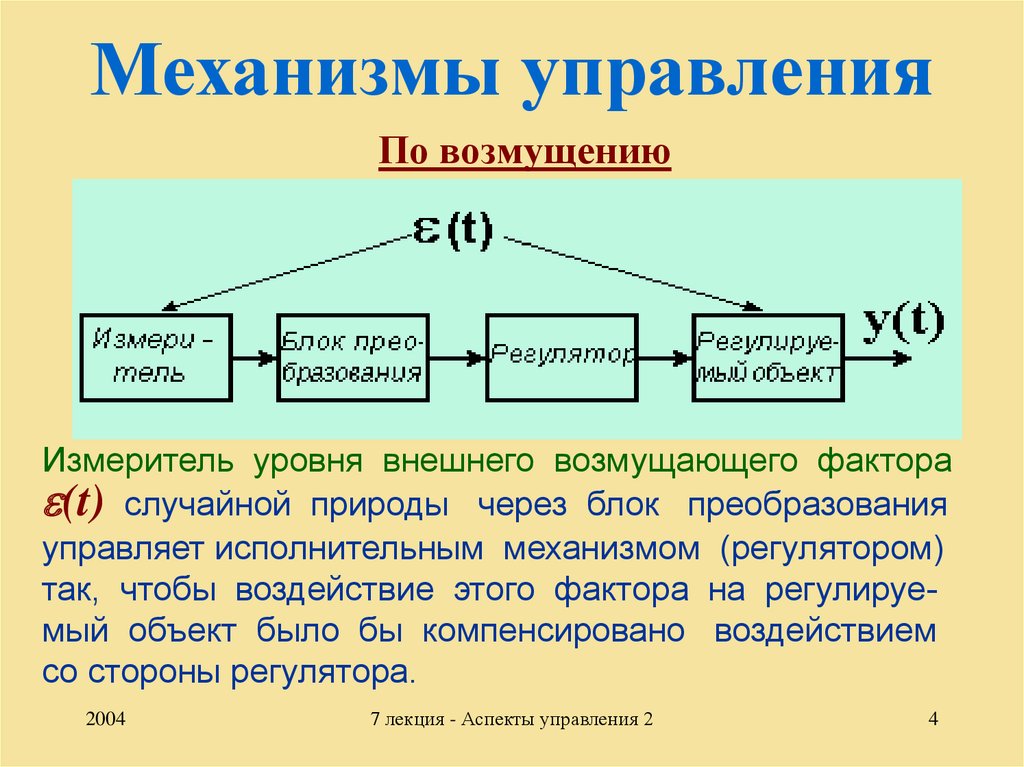 Исключение составляют именно жабры, непроницаемость которых сделала бы невозможной обеспечение рыб кислородом. Между тем через жабры идет постоянная «утечка» ионов из организма пресноводных рыб. По современным данным экстраренальные потери достигают 75-90% солей [504,681].[ …]
Исключение составляют именно жабры, непроницаемость которых сделала бы невозможной обеспечение рыб кислородом. Между тем через жабры идет постоянная «утечка» ионов из организма пресноводных рыб. По современным данным экстраренальные потери достигают 75-90% солей [504,681].[ …]
Но происходило обратное. В эпохи дестабилизации гомеостаз или функционирование природных систем в соответствии с принципом Ле Шателье — Брауна не были главными в биосфере и значительно подавлялись другими, более мощными, силами. Будучи порождением свойств космической среды, а также планетарных особенностей Земли, явления дестабилизации, тем не менее, никогда не достигали масштабов, достаточных для уничтожения жизни, хотя сильно меняли эволюцию органического мира, а отчасти и неорганической природы.[ …]
Кроме вышеуказанных «общеорганизменных» функций наличие гомеостаза организма существует еще одна очень важная особенность: живое вещество как бы создает еще одну среду обитания, а именно возможность заселения организма другими живыми существами для постоянного или временного обитания.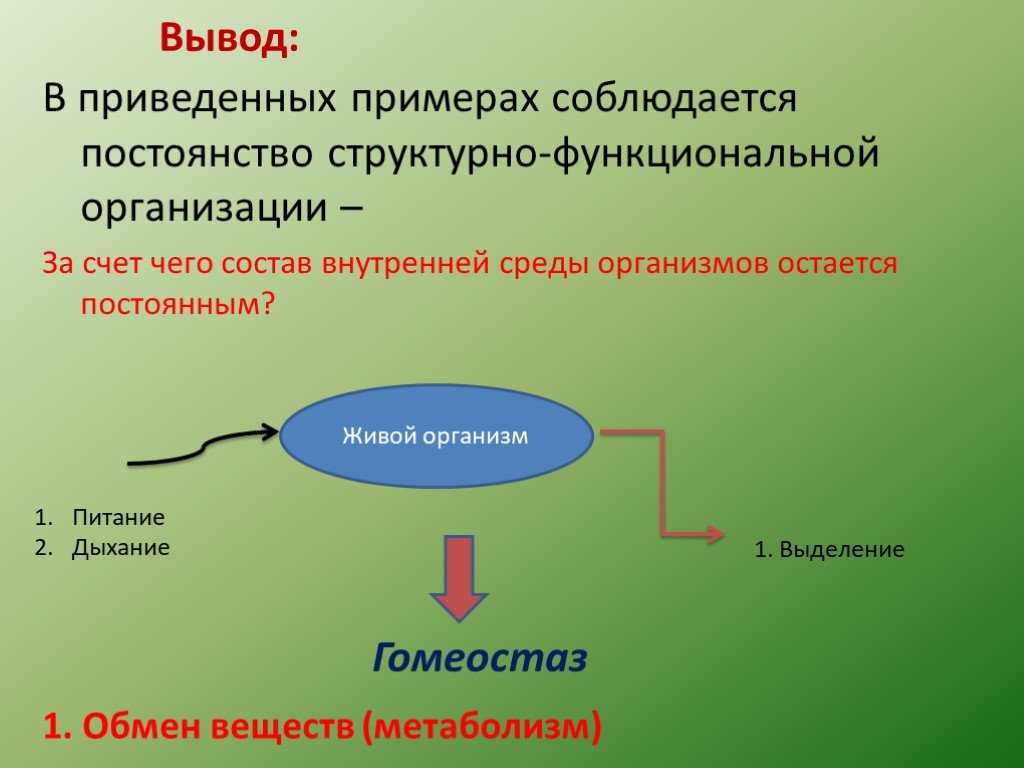 Это созданная жизнью новая биотическая среда обитания. К существам, которые заселяют эту среду, многие специалисты относят вирусы. Так, И.А. Шилов (2000) считает, что исключительная простота их устройства является вторичным явлением, даже скорее это вновь возникшая форма живых существ, полностью осврившая внутриклеточную среду в организмах других уровней. Вторым подтверждением этого тезиса является то, что вирусы обладают высокой степенью сложности и разнообразия генетической системы. Упрощение строения, ставшее возможным благодаря обязательным безусловным связям вирусов с хозяином-организмом, обеспечивающим стабильные условия жизни, затронуло даже фундаментальные свойства, присущие подавляющему большинству форм жизни: вирусы не обладают раздражимостью и лишены собственного аппарата синтеза белка. Вирусы не способны к самостоятельному существованию, и их связь с клеткой —это не только пространственная, но и жесткая функциональная связь, с которой клетка и вирус представляют некое единство.
Это созданная жизнью новая биотическая среда обитания. К существам, которые заселяют эту среду, многие специалисты относят вирусы. Так, И.А. Шилов (2000) считает, что исключительная простота их устройства является вторичным явлением, даже скорее это вновь возникшая форма живых существ, полностью осврившая внутриклеточную среду в организмах других уровней. Вторым подтверждением этого тезиса является то, что вирусы обладают высокой степенью сложности и разнообразия генетической системы. Упрощение строения, ставшее возможным благодаря обязательным безусловным связям вирусов с хозяином-организмом, обеспечивающим стабильные условия жизни, затронуло даже фундаментальные свойства, присущие подавляющему большинству форм жизни: вирусы не обладают раздражимостью и лишены собственного аппарата синтеза белка. Вирусы не способны к самостоятельному существованию, и их связь с клеткой —это не только пространственная, но и жесткая функциональная связь, с которой клетка и вирус представляют некое единство. [ …]
[ …]
Поддержание численности, оптимальной в данных условиях, называется гомеостазом популяции. Гомеостатические возможности популяций различны и осуществляются они через взаимоотношения особей между собой и с окружающей средой.[ …]
Поддержание определенной численности или равновесное состояние получило название гомеостаза популяций. Рост, снижение или постоянство численности популяций и зависит от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей), рис. 10.24.[ …]
Наступило совершенно особое третье состояние биосферы, характеризующееся нарушением гомеостаза антропогенными факторами тогда, когда все природные предпосылки благоприятствуют его поддержанию. Это состояние — время третьей природы, оцениваемое часто как недопустимо критическое, в действительности выглядит гораздо предпочтительнее целого ряда биосферных кризисов прошлого, в частности, последней ледниковой эпохи. Тогда силы неорганической субстанции в глобальном масштабе очень сильно ослабляли мощность биологического круговорота. Можно наверняка утверждать, что редуцирующее воздействие на биокруговорот современной цивилизации как и ущерб от нее биоразнообразию еще далеко не достигли уровня времени вюрма-максимума. Это внушает оптимизм, ибо позволяет с научных позиций оценивать упругость глобальной экосистемы как очень высокую.[ …]
Можно наверняка утверждать, что редуцирующее воздействие на биокруговорот современной цивилизации как и ущерб от нее биоразнообразию еще далеко не достигли уровня времени вюрма-максимума. Это внушает оптимизм, ибо позволяет с научных позиций оценивать упругость глобальной экосистемы как очень высокую.[ …]
Способность популяции поддерживать определенную численность своих особей называется гомеостазом популяции. В основе этого важнейшего, эволюционно приобретенного свойства лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой эта особь принадлежит.[ …]
Болезнью называют особое состояние организма, характеризующееся серьезным нарушением гомеостаза и развитием на этом фоне специфических приспособительных реакций (например, воспалительного процесса), направленных на его восстановление.[ …]
Основным критерием отнесения того или иного вещества к токсинам служит его способность нарушать гомеостаз какого-либо организма. При этом одно и то же вещество может быть токсйчно по отношению к одним организмам, но не токсично по отношению к другим. С другой стороны, появление токсичных веществ в пищевых цепях различных групп организмов может сложным образом сказываться на разных «звеньях» этой цепи. Какова действительная роль многих ксенобиотиков или малотоксичных веществ в сложных пищевых цепях организмов и различных экосистемах — это во многом остается пока неизвестным.[ …]
Под постоянным наблюдением находились общее состояние организма (динамика массы тела, показатели водного гомеостаза — суточное водопотребление, морфологический состав крови и др.).[ …]
В.Г.Горшковым принимается в качестве основного критерия антропогенной дестабилизации биосферы — нарушения гомеостаза в ней.[ …]
Для управления экосистемами не требуется регуляция извне — это саморегулирующаяся система. Саморегулирующий гомеостаз на экосистемном уровне обеспечен множеством управляющих механизмов. Один из них — субсистема «хищник—жертва» (рис. 5.3). Между условно выделенными кибернетическими блоками управление осуществляется посредством положительных и отрицательных связей. Положительная обратная связь «усиливает отклонение», например увеличивает чрезмерно популяцию жертвы. Отрицательная обратная связь «уменьшает отклонение», например, ограничивает рост популяции жертвы за счет увеличения численности популяции Хищников. Эта кибернетическая схема (рис. 5.3а) отлично иллюстрирует процесс коэволюции в системе «хищник—жертва», так как в этой «связке» развиваются и взаимные адаптационные процессы (см. рис. 3.5). Если в эту систему яе вмешиваются другие факторы (например, человек.уничтожил хищника), то результат саморегуляции будет описываться гомеостатическим плато (рис. 5.3 б) — областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы.[ …]
Правило затухания процессов: насыщающиеся системы с увеличением степени равновесности с окружающей их средой или внутреннего гомеостаза характеризуются затуханием в них динамических процессов. Например, темпы размножения акклиматизированных организмов по мере насыщенности сообщества затухают.[ …]
Например, темпы размножения акклиматизированных организмов по мере насыщенности сообщества затухают.[ …]
Поскольку единственной формой геологически длительного бесконфликтного существования (коэволюции) человечества и биосферы является гомеостаз (устойчивое равновесие в условиях изменяющейся среды) общества с дикой природой, сохранение которой является необходимым условием выживания человечества, то, отмечает В.Зубаков, единственная реальная форма вхождения человечества в этот гомеостаз — сокращение численности населения Земли к концу XXI в. не менее чем в 4 раза. Регулирование людской численности в биосфере должно происходить сознательно. Недопустимы такие биолягические формы регулирования, как война, эпидемия, голод.[ …]
Дифференциация тканей, появление органов, формирование нервной системы и мозга у животных привели к совершенно новой стратегии существования: вместо стабилизированного гомеостаза приоритет перешел к гибкому приспособительному поведению на основе переработки сигнальной информации и контуров регулирования. Можно сказать, что многие миллионы лет тому назад животные как бы приобрели компьютер, обладающий способностью к адаптации и огромным потенциалом самосовершенствования. В будущем у него многократно возрастет объем памяти, число обратных связей, способность к самонастройке и самообеспечению разнообразными адаптивными программами, возникнет великолепная «периферия» — органы чувств и мотивированного движения. За такой мозг, за большое число степеней свободы и широкие возможности приспособительного поведения пришлось расплачиваться необычайно возросшей напряженностью жизни, риском тактических ошибок, ситуациями стресса и частотой бифуркаций.[ …]
Можно сказать, что многие миллионы лет тому назад животные как бы приобрели компьютер, обладающий способностью к адаптации и огромным потенциалом самосовершенствования. В будущем у него многократно возрастет объем памяти, число обратных связей, способность к самонастройке и самообеспечению разнообразными адаптивными программами, возникнет великолепная «периферия» — органы чувств и мотивированного движения. За такой мозг, за большое число степеней свободы и широкие возможности приспособительного поведения пришлось расплачиваться необычайно возросшей напряженностью жизни, риском тактических ошибок, ситуациями стресса и частотой бифуркаций.[ …]
Инициированное микробное сообщество чутко реагирует на внесение минеральных удобрений. Отрицательное воздействие высоких доз минеральных удобрений оценивается по размерам зон гомеостаза и дозам, вызывающим микробный токсикоз. Величины зон гомеостаза и микробного токсикоза по азоту, калию и NPK — минимальные для дерново-подзолистой почвы и наибольшие для выщелоченного чернозема (рис. 20).[ …]
20).[ …]
Наиболее устойчивы крупные экосистемы и самая стабильная из них — биосфера, а наименее устойчивы — молодые экосистемы. Это объясняется тем, что в больших экосистемах создается саморегулирующий гомеостаз за счет взаимодействия круговоротов веществ и потоков энергии (Ю. Одум, 1975).[ …]
Гомеостаз1 (от греч. Поддержание гомеостаза — непременное условие существования как отдельных клеток и организмов, так целых биологических сообществ и экосистем.[ …]
Но для того, чтобы следы встречи с тем или иным повреждающим фактором были зафиксированы «молекулярной памятью», необходимо не менее двух условий: чтобы воздействие на организм было достаточно сильным, приводящим к выраженным нарушениям гомеостаза, и чтобы оно было довольно длительным или периодически повторяющимся. Одноразовый подъем в горы не дает долговременной адаптации к гипоксии. Дело ограничивается лишь кратковременным приспособлением. Чтобы оно стало долговременным, надо прожить в горах не менее 3—4 нед или больше. Одноразовая физическая нагрузка, сколь бы она ни была значительна, не приводит к долговременному приспособлению организма к повышенной мышечной деятельности. Для этого нужна длительная систематическая тренировка. И мало того, величина и интенсивность нагрузок в ней должны непрерывно возрастать. Ведь с увеличением тренированности выполнение данной работы, физического упражнения, все более облегчается, сопровождается все меньшими нарушениями гомеостаза. А это значит, что тренирующий эффект упражнения все более снижается. Чтобы избежать этого, надо и объем, и интенсивность нагрузки все время увеличивать, чтобы величина нарушений гомеостаза оставалась достаточно эффективной.[ …]
Одноразовая физическая нагрузка, сколь бы она ни была значительна, не приводит к долговременному приспособлению организма к повышенной мышечной деятельности. Для этого нужна длительная систематическая тренировка. И мало того, величина и интенсивность нагрузок в ней должны непрерывно возрастать. Ведь с увеличением тренированности выполнение данной работы, физического упражнения, все более облегчается, сопровождается все меньшими нарушениями гомеостаза. А это значит, что тренирующий эффект упражнения все более снижается. Чтобы избежать этого, надо и объем, и интенсивность нагрузки все время увеличивать, чтобы величина нарушений гомеостаза оставалась достаточно эффективной.[ …]
Многие генетически детерминированные реакции организмов на внешние факторы среды имеют адаптивный характер, что обеспечивает жизнь и размножение организмов в колеблющихся условиях среды. Среди адаптивных реакций различают физиологический гомеостаз и гомеостаз развития. Физиологический гомеостаз — это генетически детерминированная способность организмов противостоять колеблющимся условиям внешней среды У млекопитающих, в том числе у человека, типичным примером физиологического гомеостаза является константность осмотического давления в клетках и концентрация водородных ионов в крови вследствие функционирования почек и наличия в крови буферных субстанций. Гомеостаз развития — это генетически детерминированная способность организмов так изменять отдельные реакции, что функции организмов при этом в целом сохраняются. Например, выход из строя одной почки сопровождается тем, что остающаяся почка выполняет двойную нагрузку. Примером гомеостаза развития может быть также приобретение переболевшим организмом иммунитета против соответствующей инфекции.[ …]
Гомеостаз развития — это генетически детерминированная способность организмов так изменять отдельные реакции, что функции организмов при этом в целом сохраняются. Например, выход из строя одной почки сопровождается тем, что остающаяся почка выполняет двойную нагрузку. Примером гомеостаза развития может быть также приобретение переболевшим организмом иммунитета против соответствующей инфекции.[ …]
Важнейшим свойством экосистемы является ее устойчивость, сбалансированность обмена и происходящих в ней процессов. Способность популяций или экосистемы поддерживать устойчивое динамическое равновесие в изменяющихся условиях среды называется гомеостазом (homoios — тот же, подобный; stasis — состояние). В основе гомеостаза лежит принцип обратной связи. Для поддержания равновесия в природе не требуется внешнего управления. Пример гомеостаза — субсистема «хищник-жертва», в которой регулируются плотность популяций хищника и жертвы.[ …]
Стабильность сообщества может быть длительной лишь в том случае, если изменения среды, вызванные одними организмами, точно компенсируются деятельностью других, с противоположными экологическими требованиями. Это условие нарушается при нарушении круговорота веществ и тогда часть популяций, которые не могут выдержать конкуренции, вытесняются другими, для которых эти условия благоприятны, и гомеостаз восстанавливается.[ …]
Это условие нарушается при нарушении круговорота веществ и тогда часть популяций, которые не могут выдержать конкуренции, вытесняются другими, для которых эти условия благоприятны, и гомеостаз восстанавливается.[ …]
Суть саморегуляции у высших животных заключается в том, что при систематически меняющихся окружающих условиях сохраняется постоянство внутренней среды. Это выражается в поддержании постоялкой температуры тела, в постоянстве химического, ионного и газового состава, давления, частоты дыхания и сердечных сокращений, постоянном синтезе нужных веществ и разрушении вредных. Гомеостаз — важнейшее свойство организма — достигается благодаря совместной деятельности нервной, кровеносной, иммунной, эндокринной и пищеварительной систем.[ …]
Иммунная система человека контролирует постоянство внутренней среды организма, элиминируя различные патологические агенты биологической и ксенобиотической природы. Физиологическое предназначение иммунной системы приводит к ее постоянному контакту с поллютан-тами, попадающими в организм человека через продукты питания, воду, кожные покровы и верхние дыхательные пути (Хаитов, 1995). Способность клеток иммунной системы обеспечивать постоянный контроль гомеостаза внутренней среды организма путем элиминирования микробных и ксенобиотических агентов зависит от быстрого воспроизводства различных клеток белой крови. Таким образом, иммунная система является наиболее обновляемой и регенерирующей, что увеличивает вероятность мутаций лимфоидных клеток, часть из которых может наследоваться, особенно если токсическому воздействию подвергаются беременные женщины или работницы, которые в последующем становятся матерями.[ …]
Способность клеток иммунной системы обеспечивать постоянный контроль гомеостаза внутренней среды организма путем элиминирования микробных и ксенобиотических агентов зависит от быстрого воспроизводства различных клеток белой крови. Таким образом, иммунная система является наиболее обновляемой и регенерирующей, что увеличивает вероятность мутаций лимфоидных клеток, часть из которых может наследоваться, особенно если токсическому воздействию подвергаются беременные женщины или работницы, которые в последующем становятся матерями.[ …]
ЦП С. sumensis нормальные неполночленные. В ЦП-1 неполночлен-ность обусловлена отсутствием особей ювенильного и раннегенеративного возрастного состояния, а в ЦП-2 не обнаружены ювенильные, старые генеративные и сенильные растения. Возрастной спектр ЦП-1 трехвершинный, с преобладанием виргинильного, зрелого генеративного и сенильного онтогенетического состояния. ЦП-2 более молодая по сравнению с ЦП-1. Наличие проростков в обеих ЦП свидетельствавало о семенном размножении и поддержании гомеостаза популяций. [ …]
[ …]
Единственный выход из этого положения, как считал В. И. Вернадский, — создание ноосферы. Ноосфера — это новый этап в истории биосферы, когда определяющая роль ее развития перейдет от стихийного течения природных процессов и антропогенного воздействия на природу к гармоничному развитию природы и общества. Развитие человека и биосферы пойдет в неразрывной связи по единственному правильному пути — коэволюции, на котором не будет ни победителей, ни господства одной из сторон. Под коэволюцией следует понимать такое совместное развитие человеческого общества и биосферы, которое не выводит параметры биосферы из гомеостаза и обеспечивает ее устойчивое развитие.[ …]
Страница не найдена
Размер:
AAA
Цвет: C C C
Изображения Вкл. Выкл.
Обычная версия сайта
RUENBY
Гомельский государственный
медицинский университет
- Университет
- Университет
- История
- Руководство
- Устав и Символика
- Воспитательная деятельность
- Организация образовательного процесса
- Международное сотрудничество
- Система менеджмента качества
- Советы
- Факультеты
- Кафедры
- Подразделения
- Первичная профсоюзная организация работников
- Издания университета
- Гордость университета
- Выпускник-2021
- Первичная организация «Белорусский союз женщин»
- Одно окно
- ГомГМУ в международных рейтингах
- Структура университета
- Абитуриентам
- Приёмная комиссия
- Целевая подготовка
- Заключение, расторжение «целевого» договора
- Льготы для молодых специалистов
- Архив проходных баллов
- Карта и маршрут проезда
- Порядок приёма на 2022 год
- Специальности
- Контрольные цифры приёма в 2022 году
- Стоимость обучения
- Информация о ходе приёма документов
- Приём документов и время работы приёмной комиссии
- Порядок приёма граждан РФ, Кыргызстана, Таджикистана, Казахстана
- Pепетиционное тестирование
- Горячая линия по вопросам вступительной кампании
- Студентам
- Первокурснику
- Расписание занятий
- Расписание экзаменов
- Информация для студентов
- Студенческий клуб
- Спортивный клуб
- Общежитие
- Нормативные документы
- Практика
- Стоимость обучения
- Безопасность жизнедеятельности
- БРСМ
- Профком студентов
- Учебный центр практической подготовки и симуляционного обучения
- Многофункциональная карточка студента
- Анкетирование студентов
- Выпускникам
- Интернатура и клиническая ординатура
- Докторантура
- Аспирантура
- Магистратура
- Распределение
- Врачам и специалистам
- Профессорский консультативный центр
- Факультет повышения квалификации и переподготовки
- Иностранным гражданам
- Факультет иностранных студентов
- Стоимость обучения
- Регистрация и визы
- Полезная информация
- Правила приёма
- Информация о возможностях и условиях приема в 2022 году
- Официальные представители ГомГМУ по набору студентов
- Страхование иностранных граждан
- Приём на Подготовительное отделение иностранных граждан
- Прием иностранных граждан для обучения на английском языке / Training of foreign students in English
- Повышение квалификации и переподготовка для иностранных граждан
- Научная деятельность
- Направления научной деятельности
- Научно-педагогические школы
- Студенчеcкое научное общество
- Инновационные технологии в ГомГМУ
- Научно-исследовательская часть
- Научно-исследовательская лаборатория
- Конкурсы, гранты, стипендии
- Работа комитета по этике
- Научные мероприятия
- В помощь исследователю
- Диссертационный совет
- «Горизонт Европа»
- Патенты
- Инструкции на метод
- Совет молодых ученых
- Госпрограмма (ЧАЭС)
- Главная
Обучение и память (Раздел 4, Глава 7) Неврология в Интернете: Электронный учебник по неврологии | Кафедра нейробиологии и анатомии
Анализ анатомических и физических основ обучения и памяти является одним из больших успехов современной нейронауки. Тридцать лет назад мало что было известно о том, как работает память, но теперь мы знаем очень многое. В этой главе мы обсудим четыре вопроса, занимающих центральное место в обучении и памяти. Во-первых, каковы различные типы памяти? Во-вторых, где в мозгу находится память? Одна из возможностей состоит в том, что человеческая память похожа на микросхему памяти в персональном компьютере (ПК), которая хранит всю память в одном месте. Вторая возможность заключается в том, что наши воспоминания распределены и хранятся в разных областях мозга. В-третьих, как работает память? Какие типы изменений происходят в нервной системе при формировании и сохранении памяти, существуют ли определенные гены и белки, которые участвуют в памяти, и как память может сохраняться на всю жизнь? В-четвертых, вопрос, который важен для многих людей, особенно с возрастом: как сохранить и улучшить память и как восстановить ее, если она нарушена?
Тридцать лет назад мало что было известно о том, как работает память, но теперь мы знаем очень многое. В этой главе мы обсудим четыре вопроса, занимающих центральное место в обучении и памяти. Во-первых, каковы различные типы памяти? Во-вторых, где в мозгу находится память? Одна из возможностей состоит в том, что человеческая память похожа на микросхему памяти в персональном компьютере (ПК), которая хранит всю память в одном месте. Вторая возможность заключается в том, что наши воспоминания распределены и хранятся в разных областях мозга. В-третьих, как работает память? Какие типы изменений происходят в нервной системе при формировании и сохранении памяти, существуют ли определенные гены и белки, которые участвуют в памяти, и как память может сохраняться на всю жизнь? В-четвертых, вопрос, который важен для многих людей, особенно с возрастом: как сохранить и улучшить память и как восстановить ее, если она нарушена?
7.1 Типы памяти
Психологи и нейробиологи разделили системы памяти на две широкие категории: декларативные и недекларативные (рис. 7.1). Декларативная система памяти — это система памяти, которая, пожалуй, наиболее знакома. Это система памяти, которая имеет сознательный компонент и включает воспоминания о фактах и событиях. Такой факт, как «Париж — столица Франции», или событие, подобное предшествующему отпуску в Париже. Недекларативная память, также называемая имплицитной памятью, включает в себя типы систем памяти, которые не имеют сознательного компонента, но тем не менее чрезвычайно важны. Они включают в себя воспоминания о навыках и привычках (например, езде на велосипеде, вождении автомобиля, игре в гольф, теннис или игру на фортепиано), явление, называемое праймингом, простые формы ассоциативного обучения [например, классическое обусловливание (Павловское обусловливание)] и наконец, простые формы неассоциативного обучения, такие как привыкание и сенсибилизация. Сенсибилизация будет подробно обсуждаться далее в этой главе. Декларативная память — это «знание того, что», а недекларативная память — это «знание того, как».
7.1). Декларативная система памяти — это система памяти, которая, пожалуй, наиболее знакома. Это система памяти, которая имеет сознательный компонент и включает воспоминания о фактах и событиях. Такой факт, как «Париж — столица Франции», или событие, подобное предшествующему отпуску в Париже. Недекларативная память, также называемая имплицитной памятью, включает в себя типы систем памяти, которые не имеют сознательного компонента, но тем не менее чрезвычайно важны. Они включают в себя воспоминания о навыках и привычках (например, езде на велосипеде, вождении автомобиля, игре в гольф, теннис или игру на фортепиано), явление, называемое праймингом, простые формы ассоциативного обучения [например, классическое обусловливание (Павловское обусловливание)] и наконец, простые формы неассоциативного обучения, такие как привыкание и сенсибилизация. Сенсибилизация будет подробно обсуждаться далее в этой главе. Декларативная память — это «знание того, что», а недекларативная память — это «знание того, как».
Рисунок 7.1 |
7.2 Проверка памяти
Рисунок 7.2 |
Рисунок 7.3 |
Всем интересно узнать, насколько хорошо они помнят, поэтому давайте проведем простой тест памяти. В тесте (рис. 7.2) будет представлен список из 15 слов, затем будет пауза и вас спросят, помните ли вы некоторые из этих слов. К сожалению, вы должны положить ручку для этого теста и не читать дальше в главе, пока не выполните тест.
Этот тест памяти называется тестом DRM в честь его создателей Джеймса Диза, Генри Редигера и Кэтлин МакДермотт. Это не должно было быть уловкой, но чтобы проиллюстрировать очень интересную и важную особенность памяти. Нам нравится думать, что память подобна тому, чтобы сделать фотографию и поместить ее в ящик картотеки, чтобы потом извлечь (вызвать) как «воспоминание» именно в том виде, в каком она была помещена туда изначально (хранилась). Но память больше похожа на то, как сделать снимок, разорвать его на мелкие кусочки и разложить по разным ящикам. Затем воспоминание вызывается путем восстановления воспоминаний из отдельных фрагментов воспоминаний. Причина, по которой так много людей ошибочно полагают, что слово «сладкий» было в списке, заключается в том, что в списке было так много других слов, которые имели сладкий оттенок. «Провал» этого теста на самом деле неплохой результат. Люди с болезнью Альцгеймера обычно не говорят, что «сладкое» было в списке. Они не могут создавать нормальные ассоциации, связанные с воспроизведением воспоминаний.
Это не должно было быть уловкой, но чтобы проиллюстрировать очень интересную и важную особенность памяти. Нам нравится думать, что память подобна тому, чтобы сделать фотографию и поместить ее в ящик картотеки, чтобы потом извлечь (вызвать) как «воспоминание» именно в том виде, в каком она была помещена туда изначально (хранилась). Но память больше похожа на то, как сделать снимок, разорвать его на мелкие кусочки и разложить по разным ящикам. Затем воспоминание вызывается путем восстановления воспоминаний из отдельных фрагментов воспоминаний. Причина, по которой так много людей ошибочно полагают, что слово «сладкий» было в списке, заключается в том, что в списке было так много других слов, которые имели сладкий оттенок. «Провал» этого теста на самом деле неплохой результат. Люди с болезнью Альцгеймера обычно не говорят, что «сладкое» было в списке. Они не могут создавать нормальные ассоциации, связанные с воспроизведением воспоминаний.
Список слов дает представление об обработке и воспроизведении в памяти, но это не очень хороший тест на «сырые» способности памяти, поскольку на нее могут влиять искажения и предубеждения. Чтобы избежать этих проблем, психологи разработали другие тесты памяти. Одним из них является тест на распознавание объектов (рис. 7.3) для проверки декларативной памяти. Этот тест хорош еще и тем, что, как мы увидим позже, его можно использовать даже на животных. Тест включает в себя предоставление испытуемому двух разных объектов, и его просят запомнить эти объекты. Пауза, затем снова показываются два объекта, один из которых новый, а другой уже показывался ранее. Субъектов просят идентифицировать новый объект, и для этого им нужно вспомнить, какой из них был показан ранее. Несколько родственным тестом является тест на определение местоположения объекта, в котором испытуемых просят вспомнить местоположение объекта на двухмерной поверхности.
Чтобы избежать этих проблем, психологи разработали другие тесты памяти. Одним из них является тест на распознавание объектов (рис. 7.3) для проверки декларативной памяти. Этот тест хорош еще и тем, что, как мы увидим позже, его можно использовать даже на животных. Тест включает в себя предоставление испытуемому двух разных объектов, и его просят запомнить эти объекты. Пауза, затем снова показываются два объекта, один из которых новый, а другой уже показывался ранее. Субъектов просят идентифицировать новый объект, и для этого им нужно вспомнить, какой из них был показан ранее. Несколько родственным тестом является тест на определение местоположения объекта, в котором испытуемых просят вспомнить местоположение объекта на двухмерной поверхности.
Примеры недекларативной памяти, такие как ассоциативное обучение, можно проверить, соединив один стимул с другим, а затем проверив, научился ли субъект создавать ассоциации между двумя стимулами. Классическим примером является парадигма, разработанная русским физиологом Иваном Павловым, которую теперь называют классической или павловской обусловленностью. При классическом обусловливании (рис. 7.4) новый или слабый раздражитель (условный раздражитель, УС), такой как звук, сочетается с таким раздражителем, как пища, которая обычно вызывает рефлекторную реакцию (безусловный ответ, УР; безусловный раздражитель, УЗ), такую как слюноотделение. После достаточной тренировки с условными презентациями CS-US (которые могут быть единичным испытанием) CS способен вызывать ответ (условный ответ, CR), который часто напоминает UR (или какой-либо его аспект).
При классическом обусловливании (рис. 7.4) новый или слабый раздражитель (условный раздражитель, УС), такой как звук, сочетается с таким раздражителем, как пища, которая обычно вызывает рефлекторную реакцию (безусловный ответ, УР; безусловный раздражитель, УЗ), такую как слюноотделение. После достаточной тренировки с условными презентациями CS-US (которые могут быть единичным испытанием) CS способен вызывать ответ (условный ответ, CR), который часто напоминает UR (или какой-либо его аспект).
Рисунок 7.4 |
7.3 Локализация памяти
Теперь обратимся к вопросу о том, где находится память. Существует три основных подхода.
- Визуализация. Современные методы визуализации, такие как фМРТ (функциональная магнитно-резонансная томография) или ПЭТ (позитронно-эмиссионная томография), позволяют «увидеть» области мозга, которые активны во время определенных мозговых задач.
 Если человека поместить в сканер фМРТ и провести тест памяти, можно определить, какие области мозга активны, и эта активность предположительно связана с тем, где в мозгу обрабатывается и/или хранится память.
Если человека поместить в сканер фМРТ и провести тест памяти, можно определить, какие области мозга активны, и эта активность предположительно связана с тем, где в мозгу обрабатывается и/или хранится память.
 Если человека поместить в сканер фМРТ и провести тест памяти, можно определить, какие области мозга активны, и эта активность предположительно связана с тем, где в мозгу обрабатывается и/или хранится память.
Если человека поместить в сканер фМРТ и провести тест памяти, можно определить, какие области мозга активны, и эта активность предположительно связана с тем, где в мозгу обрабатывается и/или хранится память.
Рис. 7.5
ПЭТ-сканирование мозга во время теста на определение местоположения объекта. (из A. M. Owen, et al., J. Cog. Neurosci. 8:6, 588-602, 1996.)
На рис. 7.5 показан пример ПЭТ-сканирования человека, выполняющего тест на определение местоположения объекта. Цветовой код таков, что более яркие и красные области указывают на повышенную активность мозга. Наиболее активной областью является гиппокамп. При обсуждении памяти неоднократно упоминается гиппокамп, потому что это основная часть мозга, участвующая в декларативной функции памяти.
 Эта иллюстрация ясно показывает, что гиппокамп участвует в памяти местоположения объекта. Но, как мы скоро увидим, это не место, где хранятся все воспоминания.
Эта иллюстрация ясно показывает, что гиппокамп участвует в памяти местоположения объекта. Но, как мы скоро увидим, это не место, где хранятся все воспоминания.- Поражения головного мозга. В этой экспериментальной процедуре небольшие части мозга мышей или крыс удаляются хирургическим путем или химически инактивируются, и животных систематически исследуют, чтобы определить, повлияло ли поражение на какую-либо систему памяти.
- Болезни и травмы головного мозга. Здесь ученые используют людей, перенесших неудачные травмы головного мозга, например, в результате инсульта или опухоли головного мозга в определенной области мозга. Если у пациента обнаруживается дефицит памяти, вполне вероятно, что в этой памяти задействована область мозга, которая была повреждена.
Классическое исследование локализации памяти стало результатом операции, проведенной на Генри Молейсоне, пациенте, который в научном сообществе был известен только как «Г. М.» вплоть до своей смерти в 2008 году. Х. М. известен в литературе по неврологии, потому что его мозг дал важные сведения о локализации функции памяти. В 1950-х годах Х.М. была диагностирована неизлечимая эпилепсия, и, хотя существуют фармакологические методы лечения, в некоторых случаях единственным лечением является удаление части мозга, вызывающей припадки. Следовательно, гиппокамп H.M. был удален с двух сторон. На рис. 7.6 (справа) представлена МРТ нормального человека, показывающая область гиппокампа, тогда как на рис. 7.6 (слева) показана МРТ пациента Х.М. после удаления гиппокампа.
М.» вплоть до своей смерти в 2008 году. Х. М. известен в литературе по неврологии, потому что его мозг дал важные сведения о локализации функции памяти. В 1950-х годах Х.М. была диагностирована неизлечимая эпилепсия, и, хотя существуют фармакологические методы лечения, в некоторых случаях единственным лечением является удаление части мозга, вызывающей припадки. Следовательно, гиппокамп H.M. был удален с двух сторон. На рис. 7.6 (справа) представлена МРТ нормального человека, показывающая область гиппокампа, тогда как на рис. 7.6 (слева) показана МРТ пациента Х.М. после удаления гиппокампа.
Рисунок 7.6
Сканирование отрубей H.M. (слева) и обычный человек (справа). (Авторское право © 1997 г. Сюзанны Коркин, используется с разрешения The Wylie Agency LLC.)
Перед операцией H.M. обладал прекрасной памятью, но после операции Х.М. был очень сильный дефицит памяти. В частности, после операции способность Г. М. формировать любые новые воспоминания о фактах и событиях была серьезно нарушена; ему было очень трудно выучить любые новые словарные слова; он не мог вспомнить, что произошло накануне. Итак, если Х.М. имел интервью на следующий день после предыдущего интервью, он почти ничего не помнит об интервью или событиях во время него. Это исследование ясно показало, что гиппокамп имеет решающее значение для формирования памяти. Но тогда как Х.М. имел большие трудности с формированием новых воспоминаний о фактах и событиях, у него все еще были все его старые воспоминания о фактах и событиях. В частности, у него были все его детские воспоминания и все его воспоминания до операции. Этот тип дефицита памяти называется антероградная амнезия . (Напротив, ретроградная амнезия относится к потере старых воспоминаний.) Исследования H.M. ясно показали, что, хотя гиппокамп имеет решающее значение для формирования новых воспоминаний, он не является местом хранения старых воспоминаний. Теперь известно, что эти старые воспоминания хранятся в других частях мозга, например, в лобной коре. Процесс, посредством которого первоначально лабильная память превращается в более устойчивую форму, называется 9.
Итак, если Х.М. имел интервью на следующий день после предыдущего интервью, он почти ничего не помнит об интервью или событиях во время него. Это исследование ясно показало, что гиппокамп имеет решающее значение для формирования памяти. Но тогда как Х.М. имел большие трудности с формированием новых воспоминаний о фактах и событиях, у него все еще были все его старые воспоминания о фактах и событиях. В частности, у него были все его детские воспоминания и все его воспоминания до операции. Этот тип дефицита памяти называется антероградная амнезия . (Напротив, ретроградная амнезия относится к потере старых воспоминаний.) Исследования H.M. ясно показали, что, хотя гиппокамп имеет решающее значение для формирования новых воспоминаний, он не является местом хранения старых воспоминаний. Теперь известно, что эти старые воспоминания хранятся в других частях мозга, например, в лобной коре. Процесс, посредством которого первоначально лабильная память превращается в более устойчивую форму, называется 9. 0135 консолидация . Этот процесс включает в себя хранение памяти в другой части мозга, чем исходное место ее кодирования.
0135 консолидация . Этот процесс включает в себя хранение памяти в другой части мозга, чем исходное место ее кодирования.
Х.М. был также интересен тем, что, хотя его способность формировать новые воспоминания о фактах и событиях была серьезно нарушена, он мог формировать новые воспоминания о навыках и привычках. Хотя он мог формировать новые воспоминания о навыках и привычках, он не знал, что у него есть навыки! Он не осознавал память; он не мог заявить, что он у него есть. Это открытие ясно указывало на то, что память на навыки и привычки , а не образуются в гиппокампе. В совокупности мы узнали из этих исследований H.M. и других пациентов, что память распределена по всей нервной системе, и разные области мозга участвуют в опосредовании разных типов памяти.
На рис. 7.7 обобщены многолетние исследования анатомического локуса систем памяти. Медиальная височная доля и такие структуры, как гиппокамп, связаны с воспоминаниями о фактах и событиях; полосатое тело связано с воспоминаниями о навыках и привычках; неокортекс участвует в прайминге; миндалевидное тело связано с эмоциональными воспоминаниями; и мозжечок с простыми формами ассоциативного обучения. Нижние отделы головного мозга и спинной мозг содержат еще более простые формы обучения. Таким образом, память не хранится в одном месте мозга. Он распределяется в разных частях мозга .
Нижние отделы головного мозга и спинной мозг содержат еще более простые формы обучения. Таким образом, память не хранится в одном месте мозга. Он распределяется в разных частях мозга .
Рисунок 7.7 |
7.4 Механизмы памяти
Модельные системы для изучения механизмов памяти
Рисунок 7.8 |
Многое из того, что было изучено о нейронных и молекулярных механизмах обучения и памяти, было получено благодаря использованию так называемых «модельных систем», поддающихся клеточному анализу. Одна из таких модельных систем показана на рис. 7.8А. Аплизия калифорнийская находится в приливных бассейнах вдоль побережья Южной Калифорнии. Это около шести дюймов в длину и весит около 150 граммов. На первый взгляд это бесперспективное существо, но нейробиологи использовали технические преимущества этого животного, чтобы получить фундаментальное представление о молекулярных механизмах памяти. Действительно, пионерские открытия Эрика Кандела с использованием этого животного были отмечены получением им Нобелевской премии по физиологии и медицине в 2000 году. Aplysia имеют три технических преимущества.
7.8А. Аплизия калифорнийская находится в приливных бассейнах вдоль побережья Южной Калифорнии. Это около шести дюймов в длину и весит около 150 граммов. На первый взгляд это бесперспективное существо, но нейробиологи использовали технические преимущества этого животного, чтобы получить фундаментальное представление о молекулярных механизмах памяти. Действительно, пионерские открытия Эрика Кандела с использованием этого животного были отмечены получением им Нобелевской премии по физиологии и медицине в 2000 году. Aplysia имеют три технических преимущества.
Во-первых, он демонстрирует простые формы недекларативного (имплицитного) обучения, такие как классическое (Павловское) обусловливание, оперантное обусловливание и сенсибилизация.
Во-вторых, Аплизия имеет очень простую нервную систему. По сравнению с сотнями миллиардов нервных клеток в человеческом мозгу, вся нервная система этого животного насчитывает всего около 10 000 клеток. Эти клетки распределены по разным ганглиям, как показано на рис. 7.8В. В каждом ганглии, подобном этому, всего около 2000 клеток, но они способны опосредовать или контролировать ряд различных видов поведения. Это означает, что любое поведение может контролироваться 100 нейронами или даже меньше. Можно изучить всю нейронную цепь, лежащую в основе поведения, а затем, после обучения животного, можно изучить нейронную цепь, чтобы определить, что изменилось в цепи, лежащей в основе памяти.
Эти клетки распределены по разным ганглиям, как показано на рис. 7.8В. В каждом ганглии, подобном этому, всего около 2000 клеток, но они способны опосредовать или контролировать ряд различных видов поведения. Это означает, что любое поведение может контролироваться 100 нейронами или даже меньше. Можно изучить всю нейронную цепь, лежащую в основе поведения, а затем, после обучения животного, можно изучить нейронную цепь, чтобы определить, что изменилось в цепи, лежащей в основе памяти.
В-третьих, ганглии содержат очень большие нейроны. На рис. 7.8В показан ганглий под препаровальным микроскопом. Его диаметр составляет около 2 мм. Сферические структуры ганглиев представляют собой клеточные тела отдельных нейронов. Каждый нейрон идентифицируем и имеет уникальную локализацию и функцию. Связанное с этим преимущество заключается в том, что отдельные нейроны могут быть удалены и помещены в культуральную среду, где они могут выжить в течение многих дней. В самом деле, из ганглиев можно удалить несколько нейронов, и они восстановят свои нормальные синаптические связи, тем самым предоставив очень мощную экспериментальную систему для изучения физиологии нервных клеток и свойств связей между ними. На рис. 7.8C показан пример сенсорного нейрона (маленькая клетка справа) и моторного нейрона (большая клетка слева) в культуре. На микрофотографии можно увидеть тень микроэлектрода, проколовшего сенсорный нейрон, и тень микроэлектрода, проколовшего двигательный нейрон для проведения внутриклеточной записи.
На рис. 7.8C показан пример сенсорного нейрона (маленькая клетка справа) и моторного нейрона (большая клетка слева) в культуре. На микрофотографии можно увидеть тень микроэлектрода, проколовшего сенсорный нейрон, и тень микроэлектрода, проколовшего двигательный нейрон для проведения внутриклеточной записи.
Сенсибилизация, простая форма недекларативного обучения, поддающаяся детальному клеточному анализу
Рисунок 7.9 |
| А. | Б. | С. |
Рисунок 7.10 | ||
На рисунках 7.9 и 7.10 показано простое поведение животного и простая форма обучения, называемая сенсибилизацией. Животное тестируют, стимулируя его хвост слабым электрическим током (7.9).) или слабым механическим ударом (7.10). Эти стимулы вызывают защитный рефлекс отдергивания тела, включая хвост и близлежащие участки, такие как жабры и мясистый носик, называемый сифоном. В ответ на тестовые стимулы, подаваемые каждые пять минут, отдергивания достаточно надежны. Каждый раз они имеют примерно одинаковую продолжительность (рис. 7.9В, С, 7.10А). Но если сильный вредный раздражитель (например, удар электрическим током) воздействует на другую часть тела животного, например, на стенку его тела, последующие тестовые стимулы на хвост вызывают усиленную реакцию (рис. 7.9).Б и 7.10Б). Это пример простой формы обучения, называемой сенсибилизацией. Его определяют как усиление реакции на тестовый стимул в результате воздействия на животное сильного, как правило, вредного раздражителя. В некотором смысле животное узнает, что оно находится в «пугающей» среде. Сенсибилизация — это повсеместная форма обучения, которой обладают все животные, включая человека.
В некотором смысле животное узнает, что оно находится в «пугающей» среде. Сенсибилизация — это повсеместная форма обучения, которой обладают все животные, включая человека.
Нейронная цепь и механизмы сенсибилизации
- Нейронная цепь. Мы можем воспользоваться преимуществами больших нервных клеток Aplysia, и возможностью делать внутриклеточные записи с их помощью, чтобы проработать основную нервную цепь. На рис. 7.11 в упрощенном виде показаны ключевые компоненты лежащей в основе нейронной цепи. Стимуляция кожи активирует сенсорные нейроны (СН) (здесь показан только один из них), которые образуют глутаматергические возбуждающие синаптические связи (треугольники) с моторными нейронами (МН). Если суммарный синаптический вход в моторные нейроны достаточно велик, моторные нейроны будут активированы, и потенциалы действия будут распространяться из ганглия, вызывая возможное сокращение мышцы. Таким образом, стимуляция кожи возбуждает сенсорные нейроны, сенсорные нейроны активируют моторные нейроны, а моторные нейроны сокращают мышцы. Кроме того, должно быть очевидно, что чем больше активация моторных нейронов, тем сильнее будет последующий рефлекторный ответ. Этот рефлекс в Аплизия похожа на коленный рефлекс или рефлекс растяжения, опосредованный аналогичными схемами в спинном мозге позвоночных.
Рисунок 7.11
Нейронная цепь для оборонительного рефлекса отдергивания. - Механизмы сенсибилизации. Сенсибилизирующие стимулы приводят к высвобождению нейротрансмиттера серотонина (5-НТ) (обозначен ячейкой, помеченной IN и окрашенной в фиолетовый цвет на рис. 7.11). 5-HT модулирует силу связи между сенсорным нейроном и двигательным нейроном. Потенциал действия в сенсорном нейроне до обучения вызывает небольшой возбуждающий постсинаптический потенциал (ВПСП) в двигательном нейроне (рис. 7.12А). Но после подачи сенсибилизирующего стимула потенциал действия в сенсорном нейроне приводит к увеличению синаптического потенциала в двигательном нейроне (рис. 7.12С). Больший синаптический потенциал мотонейрона увеличивает вероятность того, что мотонейрон активируется в большей степени и вызовет большее сокращение мышцы (т. е. сенсибилизацию).
 Кроме того, должно быть очевидно, что чем больше активация моторных нейронов, тем сильнее будет последующий рефлекторный ответ. Этот рефлекс в Аплизия похожа на коленный рефлекс или рефлекс растяжения, опосредованный аналогичными схемами в спинном мозге позвоночных.
Кроме того, должно быть очевидно, что чем больше активация моторных нейронов, тем сильнее будет последующий рефлекторный ответ. Этот рефлекс в Аплизия похожа на коленный рефлекс или рефлекс растяжения, опосредованный аналогичными схемами в спинном мозге позвоночных. 7.12С). Больший синаптический потенциал мотонейрона увеличивает вероятность того, что мотонейрон активируется в большей степени и вызовет большее сокращение мышцы (т. е. сенсибилизацию).
7.12С). Больший синаптический потенциал мотонейрона увеличивает вероятность того, что мотонейрон активируется в большей степени и вызовет большее сокращение мышцы (т. е. сенсибилизацию).Один из принципов обучения и памяти, выведенный из исследований этого простого животного, и этот принцип справедлив и для нашего мозга, состоит в том, что обучение включает изменения в силе синаптических связей между нейронами . Обучение происходит не за счет реорганизации нервной системы или роста новых нейронов. Что изменилось, так это то, что изменена сила ранее существовавшего соединения.
Теперь мы можем пойти дальше в этом анализе и задаться вопросом, какие биохимические механизмы лежат в основе обучения и памяти. Мы разделим обсуждение на две временные области памяти; кратковременная память и долговременная память. Мы уже обсуждали различные типы памяти, такие как декларативная и недекларативная память. Существуют также различные временные области памяти.
Рисунок 7.12A
До сенсибилизации. Сдвиньте синий шар, чтобы управлять анимацией.Рисунок 7.12B
Во время сенсибилизации. Сдвиньте синий шар, чтобы управлять анимацией.Рисунок 7.12C
После сенсибилизации. Управляйте анимацией, перемещая синий шар.
 Кратковременная память подобна памяти телефонного номера, которая длится несколько минут, а долговременная память — это воспоминания, которые длятся дни, недели или всю жизнь.
Кратковременная память подобна памяти телефонного номера, которая длится несколько минут, а долговременная память — это воспоминания, которые длятся дни, недели или всю жизнь.- Механизмы кратковременной сенсибилизации. Механизмы кратковременной памяти для сенсибилизации проиллюстрированы на рисунке 7.12B. Сенсибилизирующий раздражитель приводит к высвобождению нейротрансмиттера 5-НТ. 5-HT связывает два типа рецепторов на сенсорном нейроне; один связан с системой DAG/PKC, а другой связан с циклической системой AMP/PKA. Это те же общие каскады, которые вы изучали в биохимии. Механизмы обучения развились, чтобы использовать некоторые биохимические механизмы, которые уже присутствуют во всех клетках, и использовать их специально для механизма памяти в нервных клетках. Протеинкиназы оказывают два типа действия. Во-первых, они регулируют свойства различных мембранных каналов (маленькие ворота на иллюстрации (рис. 7.12) представляют собой мембранные каналы, лежащие в основе инициации и реполяризации потенциала действия). Следовательно, после сенсибилизирующего стимула количество кальция, поступающего в синаптическое окончание во время потенциала действия и вызывающего высвобождение медиатора, будет увеличиваться. Кроме того, модуляция мембранных каналов приводит к повышению возбудимости сенсорного нейрона, в результате чего большее количество потенциалов действия будет вызываться тестовым раздражителем на кожу. Во-вторых, киназы регулируют другие клеточные процессы, участвующие в высвобождении медиатора, такие как размер пула синаптических пузырьков, доступных для высвобождения в ответ на приток Ca 9.0364 2+ с каждым потенциалом действия. Наконец, 5-НТ приводит к изменению свойств постсинаптического мотонейрона. В частности, 5-HT приводит к увеличению количества глутаматных рецепторов. Последствия этих процессов можно увидеть, сравнив силу синаптической связи, создаваемой одиночным потенциалом действия до (рис. 7.12А) и после (рис. 7.12С) сенсибилизации. Конкретные детали всех течений и процессов не имеют решающего значения. Однако важно знать общие принципы. Один принцип состоит в том, что обучение включает в себя использование систем вторичных мессенджеров. Здесь задействованы как системы протеинкиназы С (ПКС), так и системы протеинкиназы А (ПКА). Это достаточно общий принцип. В каждом когда-либо изученном примере обучения, будь то позвоночные или беспозвоночные, задействованы системы вторичных мессенджеров. Второй принцип заключается в том, что память включает модуляцию каналов мембраны нейронов . Они могут включать каналы, которые непосредственно регулируют высвобождение медиатора (например, Ca 2+ каналов в пресинаптическом нейроне), каналы, регулирующие возбудимость нейронов, и каналы, опосредующие синаптические ответы в постсинаптическом нейроне. Третий принцип заключается в том, что циклический AMP является одним из важнейших вторичных мессенджеров, задействованных в памяти . Зная эту информацию, вы можете начать думать о том, как можно улучшить память, основываясь на ваших знаниях лежащей в ее основе биохимии.
 5-HT связывает два типа рецепторов на сенсорном нейроне; один связан с системой DAG/PKC, а другой связан с циклической системой AMP/PKA. Это те же общие каскады, которые вы изучали в биохимии. Механизмы обучения развились, чтобы использовать некоторые биохимические механизмы, которые уже присутствуют во всех клетках, и использовать их специально для механизма памяти в нервных клетках. Протеинкиназы оказывают два типа действия. Во-первых, они регулируют свойства различных мембранных каналов (маленькие ворота на иллюстрации (рис. 7.12) представляют собой мембранные каналы, лежащие в основе инициации и реполяризации потенциала действия). Следовательно, после сенсибилизирующего стимула количество кальция, поступающего в синаптическое окончание во время потенциала действия и вызывающего высвобождение медиатора, будет увеличиваться. Кроме того, модуляция мембранных каналов приводит к повышению возбудимости сенсорного нейрона, в результате чего большее количество потенциалов действия будет вызываться тестовым раздражителем на кожу.
5-HT связывает два типа рецепторов на сенсорном нейроне; один связан с системой DAG/PKC, а другой связан с циклической системой AMP/PKA. Это те же общие каскады, которые вы изучали в биохимии. Механизмы обучения развились, чтобы использовать некоторые биохимические механизмы, которые уже присутствуют во всех клетках, и использовать их специально для механизма памяти в нервных клетках. Протеинкиназы оказывают два типа действия. Во-первых, они регулируют свойства различных мембранных каналов (маленькие ворота на иллюстрации (рис. 7.12) представляют собой мембранные каналы, лежащие в основе инициации и реполяризации потенциала действия). Следовательно, после сенсибилизирующего стимула количество кальция, поступающего в синаптическое окончание во время потенциала действия и вызывающего высвобождение медиатора, будет увеличиваться. Кроме того, модуляция мембранных каналов приводит к повышению возбудимости сенсорного нейрона, в результате чего большее количество потенциалов действия будет вызываться тестовым раздражителем на кожу. Во-вторых, киназы регулируют другие клеточные процессы, участвующие в высвобождении медиатора, такие как размер пула синаптических пузырьков, доступных для высвобождения в ответ на приток Ca 9.0364 2+ с каждым потенциалом действия. Наконец, 5-НТ приводит к изменению свойств постсинаптического мотонейрона. В частности, 5-HT приводит к увеличению количества глутаматных рецепторов. Последствия этих процессов можно увидеть, сравнив силу синаптической связи, создаваемой одиночным потенциалом действия до (рис. 7.12А) и после (рис. 7.12С) сенсибилизации. Конкретные детали всех течений и процессов не имеют решающего значения. Однако важно знать общие принципы. Один принцип состоит в том, что обучение включает в себя использование систем вторичных мессенджеров. Здесь задействованы как системы протеинкиназы С (ПКС), так и системы протеинкиназы А (ПКА). Это достаточно общий принцип. В каждом когда-либо изученном примере обучения, будь то позвоночные или беспозвоночные, задействованы системы вторичных мессенджеров.
Во-вторых, киназы регулируют другие клеточные процессы, участвующие в высвобождении медиатора, такие как размер пула синаптических пузырьков, доступных для высвобождения в ответ на приток Ca 9.0364 2+ с каждым потенциалом действия. Наконец, 5-НТ приводит к изменению свойств постсинаптического мотонейрона. В частности, 5-HT приводит к увеличению количества глутаматных рецепторов. Последствия этих процессов можно увидеть, сравнив силу синаптической связи, создаваемой одиночным потенциалом действия до (рис. 7.12А) и после (рис. 7.12С) сенсибилизации. Конкретные детали всех течений и процессов не имеют решающего значения. Однако важно знать общие принципы. Один принцип состоит в том, что обучение включает в себя использование систем вторичных мессенджеров. Здесь задействованы как системы протеинкиназы С (ПКС), так и системы протеинкиназы А (ПКА). Это достаточно общий принцип. В каждом когда-либо изученном примере обучения, будь то позвоночные или беспозвоночные, задействованы системы вторичных мессенджеров. Второй принцип заключается в том, что память включает модуляцию каналов мембраны нейронов . Они могут включать каналы, которые непосредственно регулируют высвобождение медиатора (например, Ca 2+ каналов в пресинаптическом нейроне), каналы, регулирующие возбудимость нейронов, и каналы, опосредующие синаптические ответы в постсинаптическом нейроне. Третий принцип заключается в том, что циклический AMP является одним из важнейших вторичных мессенджеров, задействованных в памяти . Зная эту информацию, вы можете начать думать о том, как можно улучшить память, основываясь на ваших знаниях лежащей в ее основе биохимии.
Второй принцип заключается в том, что память включает модуляцию каналов мембраны нейронов . Они могут включать каналы, которые непосредственно регулируют высвобождение медиатора (например, Ca 2+ каналов в пресинаптическом нейроне), каналы, регулирующие возбудимость нейронов, и каналы, опосредующие синаптические ответы в постсинаптическом нейроне. Третий принцип заключается в том, что циклический AMP является одним из важнейших вторичных мессенджеров, задействованных в памяти . Зная эту информацию, вы можете начать думать о том, как можно улучшить память, основываясь на ваших знаниях лежащей в ее основе биохимии.Мы обсудили механизм кратковременной памяти. Это «кратковременно», потому что память преходяща, и это так, потому что основные биохимические изменения преходящи. Продолжительность памяти зависит от того, как долго фосфорилируются различные белки-субстраты (например, мембранные каналы). ПКА будет активирован только на короткое время после краткого стимула, потому что циклический АМФ будет деградировать, а уровень ПКА снизится.
Рисунок 7.13
Структурные изменения сенсорных нейронов, связанные с длительной сенсибилизацией. (С изменениями из M. Wainwright et al., J. Neurosci. 22:4132-4141, 2002.)
- Механизмы длительной сенсибилизации. Между кратковременной и долговременной памятью есть два основных различия. Долгосрочные воспоминания связаны с изменениями в синтезе белка и регуляции генов, тогда как кратковременные воспоминания этого не делают. И долговременные воспоминания во многих случаях связаны со структурными модификациями. На рис. 7.13 показаны примеры отростков двух сенсорных нейронов, заполненных красителем: один от нетренированного животного, а другой от обученного. Показаны толстый аксональный отросток нейрона и множество тонких ответвлений.
Рисунок 7.
Гены, участвующие в долговременной сенсибилизации.Учитывая, что долговременная память включает изменения в экспрессии генов, основная цель нейробиологов состоит в том, чтобы определить конкретные гены и белки, которые участвуют в долговременной памяти. На рис. 7.14 показаны некоторые гены и белки, участвующие в долговременной сенсибилизации. Обратите внимание, что цАМФ, один из вторичных мессенджеров, участвующих в кратковременной памяти, также участвует в индукции долговременной памяти. Но теперь, в дополнение к своему влиянию на фосфорилирование мембранных каналов, цАМФ через PKA фосфорилирует факторы транскрипции, такие как CREB (9).0135 c AMP r esponsive e lement b inding белок). Факторы транскрипции, такие как CREB, при фосфорилировании способны регулировать экспрессию генов, что приводит к изменениям экспрессии белков, важных для индукции и поддержания долговременных изменений синаптической силы и, следовательно, долговременной памяти.
Обратите внимание, что не существует единого «волшебного гена памяти» — скорее, индукция и поддержание памяти даже в одном нейроне включает в себя участие нескольких генов и белков, которые действуют синергетически, изменяя свойства нейронов и регулируя свойства нейрона и силу синапса. Также обратите внимание, что изменения в экспрессии генов не происходят сразу — существуют разные фазы. Некоторые изменения в экспрессии генов происходят рано, некоторые даже через 24 часа после обучения.
Долговременная потенциация (LTP): вероятный синаптический механизм декларативной памяти
Считается, что устойчивая форма синаптической пластичности, называемая долговременной потенциацией (ДП), задействована во многих примерах декларативной памяти. Он присутствует в гиппокампе, который, как известно, участвует в декларативной памяти. LTP можно изучать на препаратах срезов головного мозга, где к афферентным волокнам может быть применен электрический разряд (тестовый стимул), а результирующий суммарный ВПСП может быть зарегистрирован в постсинаптическом нейроне (рис.
Подача короткой последовательности высокочастотных (100 Гц) стимулов (например, столбняка) продолжительностью 1 с к афферентному нерву вызывает два типа усиления в постсинаптических нейронах. Во-первых, существует временное облегчение, называемое посттетанической потенциацией (ПТП), которое исчезает через несколько минут. Во-вторых, за PTP следует очень устойчивое усовершенствование EPSP, называемое LTP. LTP — это своего рода механизм, необходимый для хранения долговременной памяти (рис. 7.15B).
Рисунок 7.16
Анимация индукции и экспрессии LTP.Рецептор глутамата типа NMDA имеет решающее значение для некоторых форм LTP, в частности LTP в синапсах CA3-CA1 в гиппокампе. Постсинаптические шипы нейронов СА1 имеют два типа рецепторов глутамата; Рецепторы глутамата NMDA-типа и рецепторы глутамата AMPA-типа (рис.
Даже если глутамат связывается с рецептором NMDA и вызывает конформационные изменения, оттока K + или притока Na + и Ca 2+ не происходит, потому что канал «закупорен» или заблокирован Mg 2+ . Таким образом, слабый тестовый стимул не откроет этот канал, так как он заблокирован Mg 2+ . Слабый тестовый стимул вызовет ВПСП, но этот ВПСП будет опосредован АМРА-рецептором. Как будто рецептора NMDA даже не было.
Теперь рассмотрим последствия доставки столбняка (рис. 7.16B). Во время столбняка происходит пространственное и временное суммирование ВПСП, производимых множественными афферентными синапсами общей постсинаптической клетки (рис.
7.5 Расширение памяти
Рис.
График данных улучшенной памяти у трансгенных мышей.Зная некоторые гены и белки, участвующие в памяти, мы можем использовать эту информацию, чтобы попытаться проверить роль определенных белков в памяти, а также улучшить память. Одним из экспериментальных способов решения этой проблемы является использование трансгенной технологии, при которой интересующий ген может быть сверхэкспрессирован в организме животного путем введения его в яйцеклетку. Когда потомство становится взрослым, можно проверить его результаты в тестах на память. Пример такого подхода показан на рис. 7.17. Здесь роль рецептора NMDA исследовали Джо Циен и его коллеги, работавшие в то время в Принстонском университете. Если рецепторы NMDA важны для индукции LTP, а LTP важен для декларативной памяти, можно было бы ожидать, что животные, у которых было большее количество рецепторов NMDA, обучались бы с большей готовностью. Рецепторы NMDA были сверхэкспрессированы у мышей, и мышей протестировали в тесте распознавания объектов, который обсуждался ранее в этой главе.
Чтобы оценить эффективность мыши в задаче распознавания объектов, экспериментатор измеряет количество времени в течение некоторого предопределенного периода, которое мышь тратит на изучение одного объекта, по сравнению с количеством времени, которое мышь тратит на изучение другого объекта. Если мышь вспомнит, что она уже видела один из объектов ранее, она потратит больше времени на изучение нового. Как показано на рис. 7.17, через час после первоначального предъявления объектов мыши очень хорошо справляются с тестом. Действительно, они верны примерно в 100% случаев. Они знают новый объект. Однако через день производительность памяти довольно низкая, а через три дня еще хуже. К одной неделе мыши не проявляют памяти узнавания.
А как насчет мышей, получивших дополнительные рецепторы NMDA? Теперь через день после тренировки у них отличная память! Таким образом, дополнительные рецепторы привели к улучшению работы памяти. Это хорошая новость, но плохая новость заключается в том, что через неделю память не улучшилась.
Эта анимация, созданная аспирантами Джулией Хилл и Натальей Розас Де О’Лафлин из программы магистратуры по неврологии Медицинской школы Макговерна при UTHealth, объясняет концепцию синаптической пластичности.
Проверьте свои знания
- Вопрос 1
- А
- Б
- С
- Д
- Е
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A. Трудности в изучении новых фактов
B. Трудно описать недавнее событие
C. Трудно выучить новое словарное слово
D. Трудно вспомнить детское воспоминание
E. Трудности с запоминанием лица
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A.
Гиппокамп участвует в декларативной памяти, включая память на факты.
B. Трудно описать недавнее событие
C. Трудности в изучении нового словарного слова
D. Трудно вспомнить детское воспоминание
E. Трудности с запоминанием лица
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A. Трудности в изучении новых фактов
B. Трудно описать недавнее событие Этот ответ НЕВЕРЕН.
Гиппокамп участвует в декларативной памяти, включая память на недавние события.
C. Трудности в изучении нового словарного слова
D. Трудно вспомнить детское воспоминание
E.
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A. Трудности в изучении новых фактов
B. Трудно описать недавнее событие
C. Трудности в изучении нового словарного слова Этот ответ НЕВЕРЕН.
Гиппокамп участвует в декларативной памяти, включая память на словарные слова (семантическая память).
D. Трудно вспомнить детское воспоминание
E. Трудности с запоминанием лица
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A. Трудности в изучении новых фактов
B. Трудно описать недавнее событие
C.
D. Трудно вспомнить детское воспоминание. Это ПРАВИЛЬНЫЙ ответ!
Гиппокамп участвует в формировании новых воспоминаний, но не в хранении старых воспоминаний после их консолидации.
E. Трудности с запоминанием лица
50-летний пациент с недавним повреждением гиппокампа в результате инсульта, вероятно, будет иметь все следующие дефициты, КРОМЕ:
A. Трудности в изучении новых фактов
B. Трудно описать недавнее событие
C. Трудности в изучении нового словарного слова
D. Трудно вспомнить детское воспоминание
E. Трудности с запоминанием лица Этот ответ НЕВЕРЕН.
Гиппокамп участвует в распознавании объектов.
- Вопрос 2
- А
- Б
- С
- Д
Кратковременная память может включать все следующие процессы, КРОМЕ:
A.
B. Активация систем второго мессенджера
C. Модуляция мембранных каналов
D. Модуляция выпуска передатчика
Кратковременная память может включать все следующие процессы, КРОМЕ:
A. Регуляция экспрессии генов. Ответ ПРАВИЛЬНЫЙ!
Регуляция экспрессии генов связана с долговременной памятью, а не с кратковременной памятью.
B. Активация систем второго мессенджера
C. Модуляция мембранных каналов
D. Модуляция выпуска передатчика
Кратковременная память может включать все следующие процессы, КРОМЕ:
A. Регуляция экспрессии генов
B. Активация систем второго мессенджера. Это НЕВЕРНЫЙ ответ.
Активация систем вторичных мессенджеров, таких как цАМФ, связана с кратковременной памятью.
C. Модуляция мембранных каналов
D. Модуляция выпуска передатчика
Кратковременная память может включать все следующие процессы, КРОМЕ:
A. Регуляция экспрессии генов
B. Активация систем второго мессенджера
C. Модуляция мембранных каналов Ответ НЕВЕРНЫЙ.
И потенциалозависимые, и передатчикоуправляемые каналы связаны с кратковременной памятью.
D. Модуляция выпуска передатчика
Кратковременная память может включать все следующие процессы, КРОМЕ:
A. Регуляция экспрессии генов
B. Активация систем второго мессенджера
C. Модуляция мембранных каналов
D. Модуляция срабатывания передатчика. Это НЕВЕРНЫЙ ответ.
Изменения синаптической силы связаны с кратковременной памятью.
- Вопрос 3
- А
- Б
- С
- Д
- Е
Классическое кондиционирование является примером:
А. Семантическая память
Б. Эпизодическая память
C. Неявная память
D. Декларативная память
E. Неассоциативная память
Классическое кондиционирование является примером:
A. Семантическая память Это НЕВЕРНЫЙ ответ.
Семантическая память — это тип декларативной памяти, тогда как классическая обусловленность — это тип недекларативной (имплицитной) памяти.
Б. Эпизодическая память
C. Неявная память
D. Декларативная память
E. Неассоциативная память
Классическое кондиционирование является примером:
А. Семантическая память
B. Эпизодическая память Это НЕВЕРНЫЙ ответ.
Эпизодическая память — это тип декларативной памяти, тогда как классическая обусловленность — это тип недекларативной (имплицитной) памяти.
C. Неявная память
D. Декларативная память
E. Неассоциативная память
Классическое кондиционирование является примером:
А. Семантическая память
Б. Эпизодическая память
C. Неявная память.
D. Декларативная память
E. Неассоциативная память
Классическое кондиционирование является примером:
А. Семантическая память
Б. Эпизодическая память
C. Неявная память
D. Декларативная память Это НЕВЕРНЫЙ ответ.
Классическое обусловливание является примером недекларативной памяти.
E. Неассоциативная память
Классическое кондиционирование является примером:
А. Семантическая память
Б. Эпизодическая память
C. Неявная память
D. Декларативная память
E. Неассоциативная память Это НЕВЕРНЫЙ ответ.
Классическое обусловливание — это форма ассоциативного обучения, которая отличается от примеров неассоциативной памяти, таких как сенсибилизация.
Пожертвования в пользу Neuroscience Online помогут финансировать разработку новых функций и контента.
 Протеиновые фосфатазы удаляют фосфатные группы белков-субстратов, которые «хранят» память.
Протеиновые фосфатазы удаляют фосфатные группы белков-субстратов, которые «хранят» память. Вдоль ветвей видны небольшие точечные вздутия или варикозы. Эти варикозы представляют собой пресинаптические окончания сенсорных нейронов, которые вступают в контакт с другими нейронами, такими как двигательные нейроны. (Моторные нейроны не видны, потому что красителем были заполнены только сенсорные нейроны.) В части B на рис. 7.13 показан пример сенсорного нейрона, которому был введен краситель у нетренированного животного, а в части A показан нейрон, которому был введен краситель. заполняли красителем через 24 ч после сенсибилизирующего тренинга. Между этими двумя нейронами есть большая разница. Нейрон обученного животного имеет большее количество ветвей и большее количество синаптических варикозов, чем нейрон необученного животного. Следовательно, долговременная память включает в себя изменения в структуре нейронов, в том числе рост новых отростков и синапсов. Итак, в той мере, в какой вы вспомните что-нибудь об этом материале в памяти завтра, или на следующей неделе, или в следующем году, это будет потому, что в вашем мозгу начинаются структурные изменения в синапсах!
Вдоль ветвей видны небольшие точечные вздутия или варикозы. Эти варикозы представляют собой пресинаптические окончания сенсорных нейронов, которые вступают в контакт с другими нейронами, такими как двигательные нейроны. (Моторные нейроны не видны, потому что красителем были заполнены только сенсорные нейроны.) В части B на рис. 7.13 показан пример сенсорного нейрона, которому был введен краситель у нетренированного животного, а в части A показан нейрон, которому был введен краситель. заполняли красителем через 24 ч после сенсибилизирующего тренинга. Между этими двумя нейронами есть большая разница. Нейрон обученного животного имеет большее количество ветвей и большее количество синаптических варикозов, чем нейрон необученного животного. Следовательно, долговременная память включает в себя изменения в структуре нейронов, в том числе рост новых отростков и синапсов. Итак, в той мере, в какой вы вспомните что-нибудь об этом материале в памяти завтра, или на следующей неделе, или в следующем году, это будет потому, что в вашем мозгу начинаются структурные изменения в синапсах! 14
14 
 7.15A). Если проводящий путь стимулируется неоднократно (например, каждую минуту), амплитуда ВПСП остается постоянной (рис. 7.15Б).
7.15A). Если проводящий путь стимулируется неоднократно (например, каждую минуту), амплитуда ВПСП остается постоянной (рис. 7.15Б).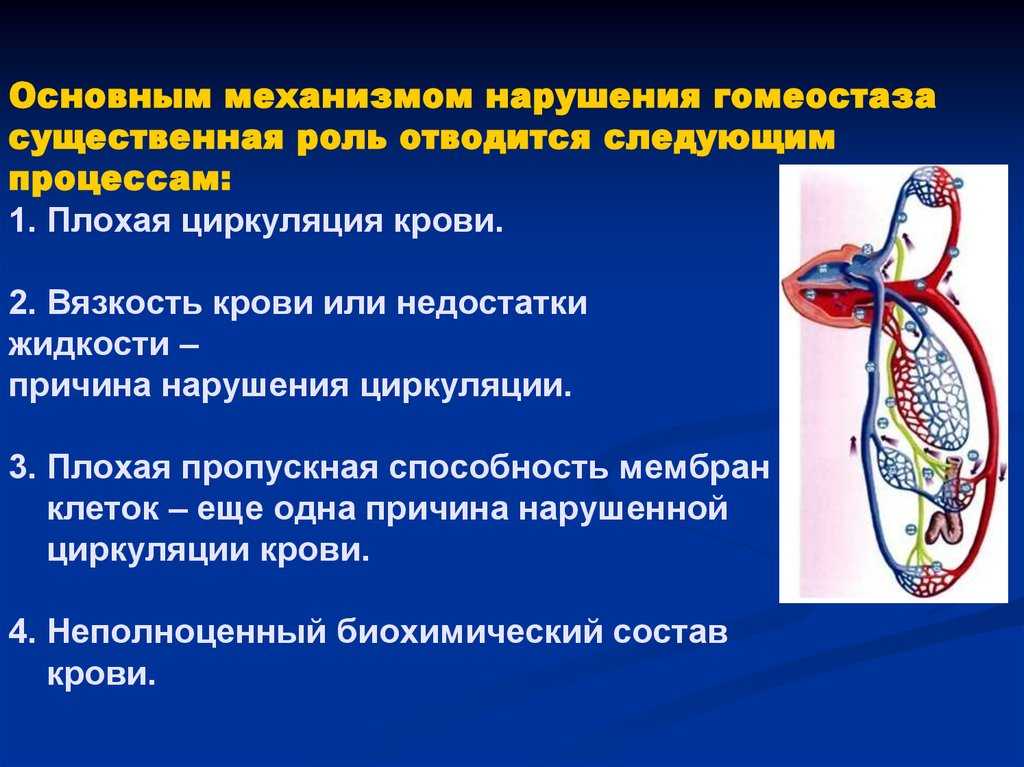 7.16A). Оба рецептора проницаемы для Na + и K + , но тип NMDA имеет две дополнительные функции. Во-первых, помимо того, что он проницаем для Na + , он также обладает значительной проницаемостью для Ca 2+ . Во-вторых, этот канал обычно блокируется Mg 2+ .
7.16A). Оба рецептора проницаемы для Na + и K + , но тип NMDA имеет две дополнительные функции. Во-первых, помимо того, что он проницаем для Na + , он также обладает значительной проницаемостью для Ca 2+ . Во-вторых, этот канал обычно блокируется Mg 2+ .  7.15А). Следовательно, мембранный потенциал постсинаптического нейрона будет значительно деполяризован, в гораздо большей степени, чем деполяризация, вызванная единичным тестовым афферентным стимулом. Поскольку внутренняя часть клетки становится положительной при большом синаптическом входе, положительно заряженный Mg 2+ отталкивается внутренней позитивностью и «выталкивается» из канала. Теперь канал отключен, и Ca 2+ может проникнуть в позвоночник через незаблокированный рецептор NMDA. Ca 2+ , попадая в клетку, активирует различные протеинкиназы, которые затем запускают долговременные изменения. Одним из компонентов долговременных изменений является внедрение новых АМРА-рецепторов в постсинаптическую мембрану (рис. 7.16С). Следовательно, после столбняка медиатор, высвобождаемый из пресинаптического нейрона тестовым стимулом, будет связываться с большим числом рецепторов постсинаптического нейрона. Если больше рецепторов связано и, следовательно, открыто, будет произведен более крупный (потенцированный) ВПСП (т.
7.15А). Следовательно, мембранный потенциал постсинаптического нейрона будет значительно деполяризован, в гораздо большей степени, чем деполяризация, вызванная единичным тестовым афферентным стимулом. Поскольку внутренняя часть клетки становится положительной при большом синаптическом входе, положительно заряженный Mg 2+ отталкивается внутренней позитивностью и «выталкивается» из канала. Теперь канал отключен, и Ca 2+ может проникнуть в позвоночник через незаблокированный рецептор NMDA. Ca 2+ , попадая в клетку, активирует различные протеинкиназы, которые затем запускают долговременные изменения. Одним из компонентов долговременных изменений является внедрение новых АМРА-рецепторов в постсинаптическую мембрану (рис. 7.16С). Следовательно, после столбняка медиатор, высвобождаемый из пресинаптического нейрона тестовым стимулом, будет связываться с большим числом рецепторов постсинаптического нейрона. Если больше рецепторов связано и, следовательно, открыто, будет произведен более крупный (потенцированный) ВПСП (т.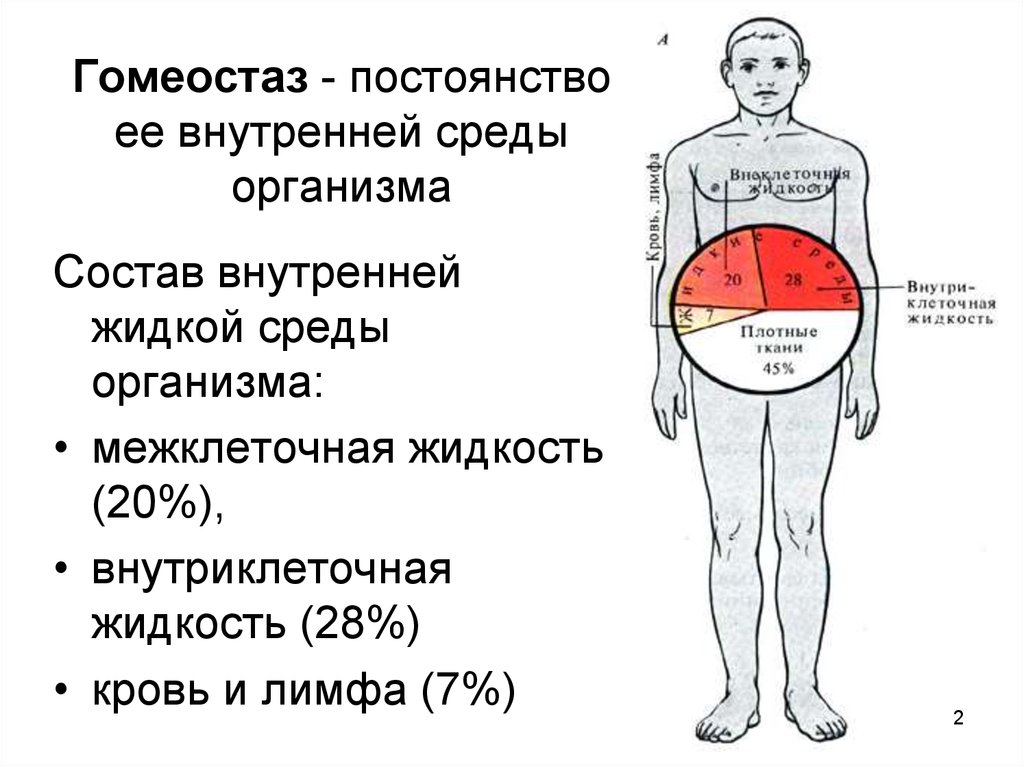 е. ДП) (рис. 7.16С). Помимо увеличения количества постсинаптических AMPA-рецепторов, есть данные, что большее количество медиатора высвобождается из пресинаптических нейронов. Комбинация пресинаптических и постсинаптических эффектов будет действовать синергетически, увеличивая размер синаптического потенциала в постсинаптических нейронах. Обратите внимание, что этот пример синаптического механизма для декларативной памяти имеет некоторое сходство с синаптическим механизмом для примера недекларативной памяти (сенсибилизация), рассмотренным ранее. Хотя конкретные детали различаются, оба включают активацию систем вторичных мессенджеров и регуляцию мембранных каналов. Таким образом, на фундаментальном механистическом уровне, по-видимому, нет существенных различий между двумя основными классами систем памяти. Основное различие, по-видимому, заключается в области мозга и нейронной цепи, в которую встроен механизм обучения.
е. ДП) (рис. 7.16С). Помимо увеличения количества постсинаптических AMPA-рецепторов, есть данные, что большее количество медиатора высвобождается из пресинаптических нейронов. Комбинация пресинаптических и постсинаптических эффектов будет действовать синергетически, увеличивая размер синаптического потенциала в постсинаптических нейронах. Обратите внимание, что этот пример синаптического механизма для декларативной памяти имеет некоторое сходство с синаптическим механизмом для примера недекларативной памяти (сенсибилизация), рассмотренным ранее. Хотя конкретные детали различаются, оба включают активацию систем вторичных мессенджеров и регуляцию мембранных каналов. Таким образом, на фундаментальном механистическом уровне, по-видимому, нет существенных различий между двумя основными классами систем памяти. Основное различие, по-видимому, заключается в области мозга и нейронной цепи, в которую встроен механизм обучения. 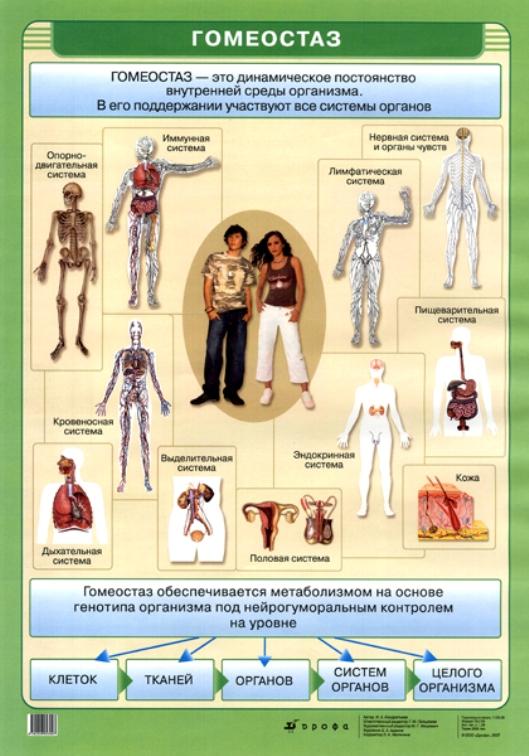 7.17
7.17 
 Этот несколько разочаровывающий вывод не должен вызывать удивления. Хотя рецепторы NMDA важны для памяти, это еще не все. Как указывалось ранее в этой главе, память включает в себя синергетическое взаимодействие нескольких генов и белков. Таким образом, для дальнейшего улучшения памяти необходимо будет манипулировать несколькими генами. В настоящее время это сделать сложно, но, вероятно, в ближайшем будущем это станет возможным. Также будет возможно сверхэкспрессировать интересующие гены в целевых областях человеческого мозга. Будущее лечения людей с нарушениями памяти выглядит многообещающим.
Этот несколько разочаровывающий вывод не должен вызывать удивления. Хотя рецепторы NMDA важны для памяти, это еще не все. Как указывалось ранее в этой главе, память включает в себя синергетическое взаимодействие нескольких генов и белков. Таким образом, для дальнейшего улучшения памяти необходимо будет манипулировать несколькими генами. В настоящее время это сделать сложно, но, вероятно, в ближайшем будущем это станет возможным. Также будет возможно сверхэкспрессировать интересующие гены в целевых областях человеческого мозга. Будущее лечения людей с нарушениями памяти выглядит многообещающим. В 2011 году он занял третье место в первом конкурсе видеороликов Общества нейронауки о мозге.
В 2011 году он занял третье место в первом конкурсе видеороликов Общества нейронауки о мозге.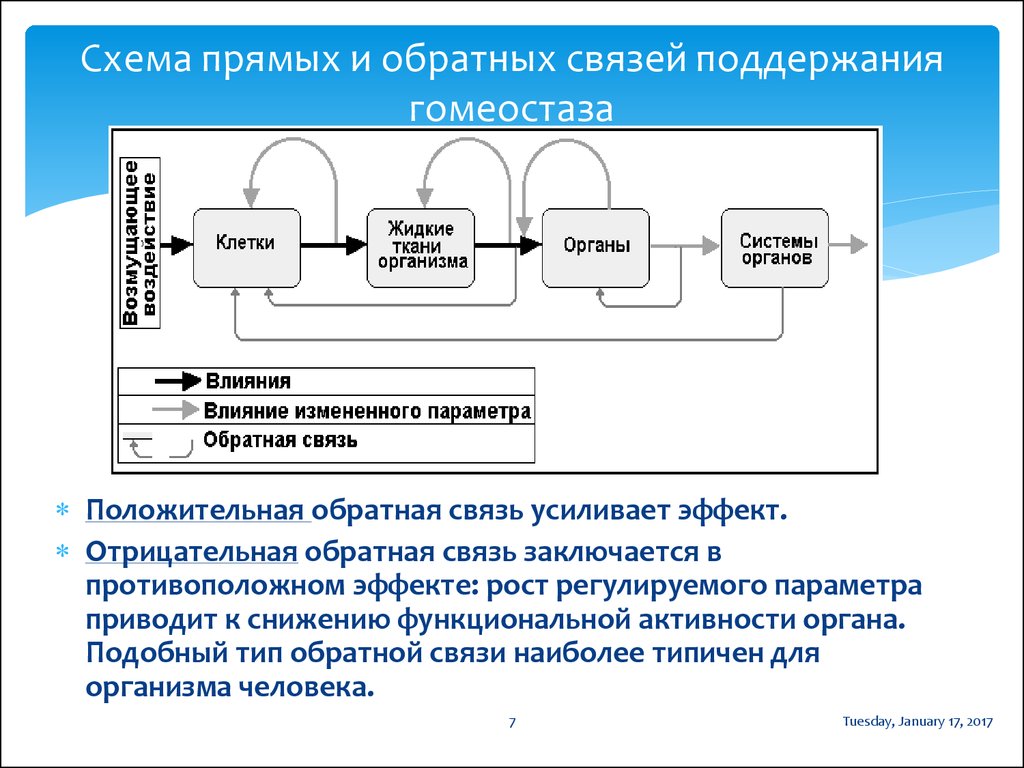 Трудности в изучении новых фактов Этот ответ НЕВЕРЕН.
Трудности в изучении новых фактов Этот ответ НЕВЕРЕН. Трудности с запоминанием лица
Трудности с запоминанием лица Трудно выучить новое словарное слово
Трудно выучить новое словарное слово Регуляция экспрессии генов
Регуляция экспрессии генов


 Это ПРАВИЛЬНЫЙ ответ!
Это ПРАВИЛЬНЫЙ ответ! 
Введение в петли обратной связи
Ментальные модели|Время чтения: 7 минут
Петли обратной связи создаются, когда реакции влияют сами на себя и могут быть положительными или отрицательными.
Рассмотрим термостат, регулирующий комнатную температуру. Это пример петли отрицательной обратной связи. При повышении температуры термостат выключает печь, позволяя комнате оставаться при заданной температуре. Когда температура падает ниже заданной температуры, печь снова зажигается, чтобы вернуть комнату в равновесное состояние. Другие примеры включают температуру тела и финансовые рынки.
Говоря о кредитных проблемах в 2008-2009 годах, вице-председатель Berkshire Hathaway Чарли Мангер объяснил: страна. Последовал свободный спад деловой активности, ускоряющийся темпами, которых я никогда прежде не видел. США — и большая часть мира — оказались в ловушке порочного круга негативной обратной связи. Страх привел к сокращению бизнеса, а это, в свою очередь, привело к еще большему страху.
* * *
В книге «Воспитание спекулянта» Виктор Нидерхоффер говорит:
Одной из общих черт, которыми обладает вся жизнь, является механизм поддержания упорядоченных условий. Эта тенденция называется гомеостазом . В теории систем это называется отрицательной обратной связью … Обычным гомеостатическим поведением у людей является регулирование температуры. Если температура поднимается выше оптимальной для нормальной человеческой деятельности 98,6 F, сенсоры на коже обнаруживают это и сигнализируют мозгу, что повышение произошло.
 Мозг передает информацию эффекторам, которые увеличивают приток крови к коже. Это вызывает потоотделение. Потеря тепла, вызванная испарением, снижает температуру тела. Когда тело охлаждается ниже определенной точки, запускается аналогичный механизм, на этот раз уменьшающий кровоток и вызывающий дрожь. Эта деятельность генерирует тепло за счет физической активности, что повышает температуру тела.
Мозг передает информацию эффекторам, которые увеличивают приток крови к коже. Это вызывает потоотделение. Потеря тепла, вызванная испарением, снижает температуру тела. Когда тело охлаждается ниже определенной точки, запускается аналогичный механизм, на этот раз уменьшающий кровоток и вызывающий дрожь. Эта деятельность генерирует тепло за счет физической активности, что повышает температуру тела.В «Универсальных принципах дизайна» Уильям Лидвелл, Джилл Батлер и Критина Холден пишут:
Каждое действие вызывает равное и противоположное противодействие. Когда реакции зацикливаются, чтобы воздействовать на самих себя, создается петля обратной связи . Все системы реального мира состоят из множества таких взаимодействующих петель обратной связи — животных, машин, предприятий и экосистем, и это лишь некоторые из них. Существует два типа контуров обратной связи: положительный и отрицательный. Положительная обратная связь усиливает производительность системы, что приводит к ее росту или упадку.
Циклы положительной обратной связи эффективны для создания изменений, но обычно приводят к негативным последствиям, если их не смягчить петлями отрицательной обратной связи. Например, в ответ на травмы головы и шеи в футболе в конце 1950-х дизайнеры создали пластиковые футбольные шлемы с внутренней прокладкой для замены кожаных шлемов. Шлемы обеспечивали большую защиту, но заставляли игроков идти на все больший риск при отборе мяча. Произошло больше травм головы и шеи (после введения пластиковых касок), чем раньше. Концентрируясь на проблеме изолированно (например, не принимая во внимание изменения в поведении игроков, дизайнеры непреднамеренно создали петлю положительной обратной связи, в которой игроки использовали голову и шею все более рискованными способами. Это привело к большему количеству травм, что привело к дополнительным изменениям конструкции, которые сделали шлем раковины более твердые и более мягкие и т.
Отрицательные петли обратной связи эффективны для сопротивления изменениям. Например, Segway Human Transporter использует отрицательную обратную связь для поддержания равновесия. Когда водитель наклоняется вперед или назад, Segway ускоряется или замедляется, чтобы удерживать систему в равновесии. Чтобы достичь этого плавно, Segway делает сотни регулировок каждую секунду. Учитывая высокую скорость регулировки, колебания вокруг точки равновесия настолько малы, что их невозможно обнаружить. Однако, если бы в секунду производилось меньше регулировок, колебания увеличивались бы в размерах, и поездка становилась бы все более прерывистой.
Ключевой урок петель обратной связи заключается в том, что все взаимосвязано — изменение одной переменной в системе повлияет на другие переменные в этой системе и других системах. Это важно, потому что это означает, что дизайнеры должны учитывать не только отдельные элементы дизайна, но и их отношение к дизайну в целом и к окружающей среде в целом.
 Отрицательная обратная связь гасит выход, стабилизирует систему вокруг точки равновесия.
Отрицательная обратная связь гасит выход, стабилизирует систему вокруг точки равновесия.  д.
д.
Из «Алгебры графов» Кортни Браун:
Циклы обратной связи обычно используются для регулирования и контроля. Петля обратной связи похожа на ввод, но ее источник находится внутри самой системы, а не снаружи системы . Во многих системах выходные данные повторно входят в систему в качестве других входных данных. Именно это происходит с микрофоном и динамиками, когда звук из динамиков возвращается в микрофон, часто вызывая громкий визг.
Санджай Бакши, приглашенный профессор MDI, написал электронное письмо одному из своих студентов по поводу положительных отзывов:
На мой взгляд, неправильно всегда рассматривать циклы положительной обратной связи в бизнесе как разрушительные, хотя они вполне могут быть таковыми. Например, набег на банк может поставить его на колени за очень короткий период времени, и он может распространиться (системный риск) на другие банки. Точно так же пузыри на фондовом рынке можно рассматривать как циклы положительной обратной связи — высокие цены подпитывают оптимизм, который подпитывает высокие цены — это не длится вечно, но может длиться очень долго.
Я думаю, вы на правильном пути, когда визуализируете петлю положительной обратной связи как механизм, встроенный в петлю отрицательной обратной связи. Чтобы проиллюстрировать, почему медвежьи рынки следуют за бычьими? Потому что в долгосрочной перспективе рынки работают внутри петли отрицательной обратной связи со встроенными корректирующими механизмами. Когда цены уходят слишком далеко от базовых значений, есть силы, которые тянут их назад. Например, когда акции становятся слишком дешевыми по сравнению со стоимостью замещения базовых активов, нет стимула для создания новых мощностей, и, вероятно, произойдет консолидация отрасли, когда сильные игроки в отрасли вместо создания новых мощностей покупают вне конкурентов.
И наоборот, когда цены на акции растут настолько высоко, что становятся намного больше, чем восстановительная стоимость базовых активов, эти высокие цены создают сильные стимулы для создания новых мощностей. Таким образом, нехватка следует за избытком, за нехваткой….
Но что в первую очередь вызвало спекулятивный пузырь? Почему люди внезапно впадают в эйфорию по поводу отрасли или сектора и инвестируют в них в унисон? Эта часть ответа лучше объясняется циклами положительной обратной связи.
Также важно рассматривать петли положительной обратной связи как средство объяснения некоторых экстремальных успехов в бизнесе. Я привел два примера в классе доминирующей газеты и Wal-Mart. Но можно думать и о других. Например, в некоторых отраслях первопроходец имеет большое преимущество. Он идет и захватывает очень большую часть рынка и получает эффект масштаба. И как только он это сделал, его становится очень трудно вытеснить.

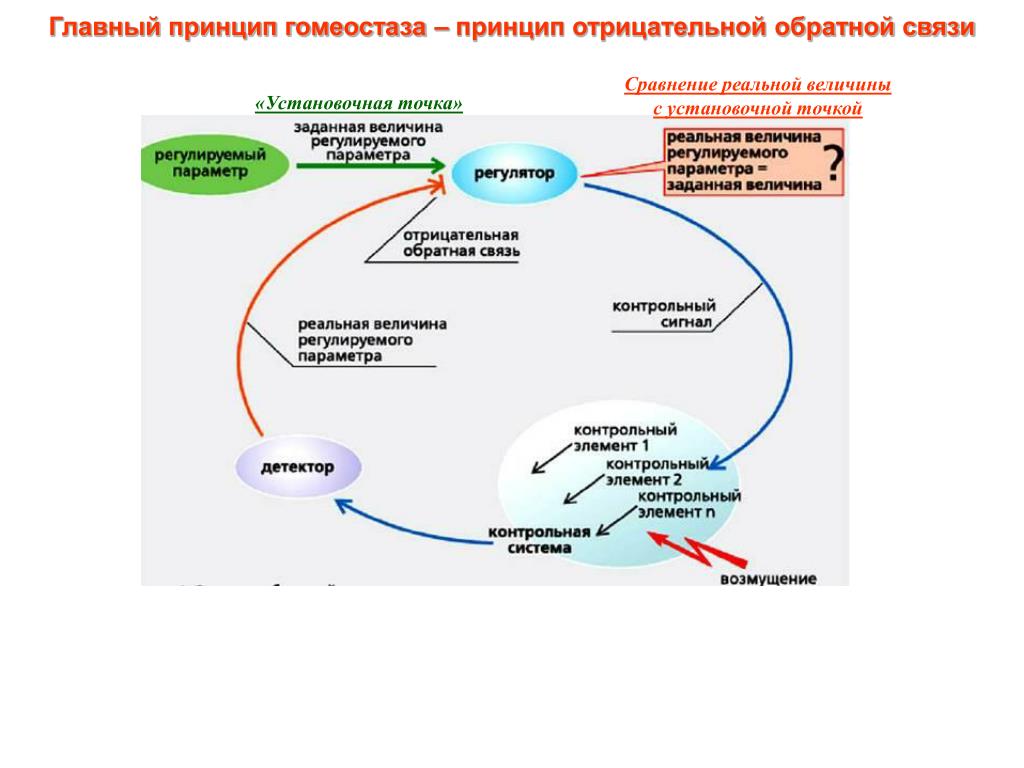 – отсюда отрицательная обратная связь.
– отсюда отрицательная обратная связь.В 2005 году Майкл Мобуссен, главный инвестиционный стратег Legg Mason Capital Management, предложил 1 :
Последняя идея имеет отношение к системам, которые демонстрируют суперлинейное поведение по сравнению с теми, которые демонстрируют сублинейное поведение.
Сублинейные системы связаны с максимизацией эффективности на единицу продукции в относительно зрелых отраслях. Использование капитала для инноваций может быть не лучшим путем для таких компаний: вместо этого они должны работать над эффективностью.
 В сверхлинейных системах, когда вы добавляете больше, вы создаете больше: существует положительная обратная связь. Одним из наиболее прямых применений этого на рынках являются сетевые эффекты: ценность товара или услуги возрастает по мере того, как все больше людей используют этот товар или услугу. Типичным примером может служить eBay. Они начали со своего основного аукционного бизнеса, который имеет очень явные сетевые эффекты. Затем они перешли на PayPal, который занимается платежами. Он также извлек выгоду из сетевых эффектов. На прошлой неделе они объявили о приобретении Skype. Опять же, это бизнес с сетевыми эффектами.
В сверхлинейных системах, когда вы добавляете больше, вы создаете больше: существует положительная обратная связь. Одним из наиболее прямых применений этого на рынках являются сетевые эффекты: ценность товара или услуги возрастает по мере того, как все больше людей используют этот товар или услугу. Типичным примером может служить eBay. Они начали со своего основного аукционного бизнеса, который имеет очень явные сетевые эффекты. Затем они перешли на PayPal, который занимается платежами. Он также извлек выгоду из сетевых эффектов. На прошлой неделе они объявили о приобретении Skype. Опять же, это бизнес с сетевыми эффектами.В 2002 году Mauboussin привел пример муравьев и положительные отзывы:
Сила этого коллективного воздействия не угасла в природе.
Когда исследователи поняли эту коллективную способность, они решили подшутить над муравьями. В контролируемых условиях ученые разместили два источника пищи на одинаковом расстоянии от гнезда. Как оказалось, муравьи в итоге использовали только один из путей, хотя тот, который они выбрали, был случайным. Почему? Поскольку они следуют химическим следам, еще пара муравьев, идущих по одному пути, привлечет других муравьев, запуская петлю положительной обратной связи. Таким образом, вместо того, чтобы найти оптимальное решение, у муравьев есть один переполненный путь и равноудаленный пустой путь.
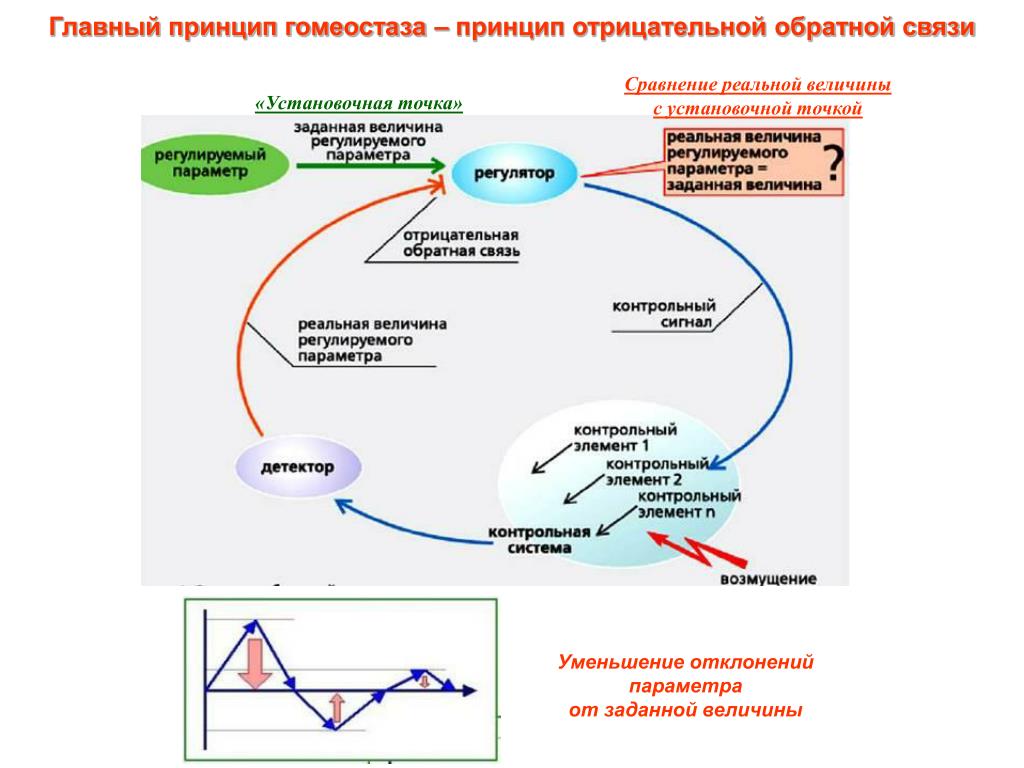 Вот тут-то и появляются рассказы Джонсона о муравьях. Как муравьи это делают? Муравьи-собиратели покидают гнездо с единственной целью — найти и добыть пищу. У них также есть способность уходить и следовать по химическим следам. Сначала они расходятся хаотично. Когда муравьи, нашедшие пищу, возвращаются в гнездо, они оставляют химический след, по которому могут следовать их сестры. Исследования показывают, что этот процесс позволяет муравьям последовательно находить кратчайший путь к еде.
Вот тут-то и появляются рассказы Джонсона о муравьях. Как муравьи это делают? Муравьи-собиратели покидают гнездо с единственной целью — найти и добыть пищу. У них также есть способность уходить и следовать по химическим следам. Сначала они расходятся хаотично. Когда муравьи, нашедшие пищу, возвращаются в гнездо, они оставляют химический след, по которому могут следовать их сестры. Исследования показывают, что этот процесс позволяет муравьям последовательно находить кратчайший путь к еде. Удивительно, но природа предвидела и эту проблему. Как оказалось, муравьи периодически сбиваются с основного пути и снова начинают случайный процесс поиска. Муравьи запрограммированы на поиск баланса между использованием известного источника пищи и поиском следующего источника пищи. Муравьи запрограммированы на поиск разнообразия.
Удивительно, но природа предвидела и эту проблему. Как оказалось, муравьи периодически сбиваются с основного пути и снова начинают случайный процесс поиска. Муравьи запрограммированы на поиск баланса между использованием известного источника пищи и поиском следующего источника пищи. Муравьи запрограммированы на поиск разнообразия. В Earth in the Balance Эл Гор пишет:
Когда я пролетал над тропическим лесом Амазонки в небольшом месте, меня поразило то, что произошло сразу после того, как гроза прошла через участок леса: как только дождь прекратился, с деревьев стали подниматься тучи влаги, образуя новые дождевые тучи, которые двигались на запад, гонимые ветром, где давали воду для нового дождя, выпадающего из новых гроз. Любое прерывание этого естественного процесса может оказать огромное влияние. Когда большие площади тропических лесов сжигаются, количество осадков, перерабатываемых в прилегающие районы, резко сокращается, лишая эти районы дождя, необходимого для поддержания их здорового состояния.
 Если обезлесенная территория достаточно велика, количество осадков, удаленных с прилегающих территорий, будет достаточным, чтобы вызвать усиливающийся цикл засухи, который медленно убивает больше деревьев, тем самым еще больше сокращая повторное использование осадков и ускоряя гибель леса, в свою очередь.
Если обезлесенная территория достаточно велика, количество осадков, удаленных с прилегающих территорий, будет достаточным, чтобы вызвать усиливающийся цикл засухи, который медленно убивает больше деревьев, тем самым еще больше сокращая повторное использование осадков и ускоряя гибель леса, в свою очередь.От Томаса Гетца в Wired:
… Итак, петли обратной связи работают. Почему? Почему представление собственных данных каким-то образом заставляет нас действовать? Отчасти это связано с тем, что обратная связь связана с чем-то ключевым в человеческом опыте, даже в нашем биологическом происхождении. Как и любой организм, люди являются саморегулирующимися существами с множеством систем, работающих для достижения гомеостаза. Сама эволюция, в конце концов, представляет собой петлю обратной связи, хотя и настолько растянутую, что она незаметна для отдельного человека. Петли обратной связи — это то, как мы учимся, называем ли мы это методом проб и ошибок или корректировкой курса.
 Во многих сферах жизни мы добиваемся успеха, когда у нас есть некоторое представление о том, где мы находимся, и какая-то оценка нашего прогресса. Действительно, мы склонны жаждать такого рода информации; это то, что мы интуитивно хотим знать, хорошо это или плохо. Как выразился Бандура из Стэнфорда, «люди — это активные, целеустремленные организмы». Обратная связь затрагивает эти устремления.
Во многих сферах жизни мы добиваемся успеха, когда у нас есть некоторое представление о том, где мы находимся, и какая-то оценка нашего прогресса. Действительно, мы склонны жаждать такого рода информации; это то, что мы интуитивно хотим знать, хорошо это или плохо. Как выразился Бандура из Стэнфорда, «люди — это активные, целеустремленные организмы». Обратная связь затрагивает эти устремления.* * *
Ричард Кох:
В отсутствие петель обратной связи естественное распределение явлений было бы 50/50 — входы с заданной частотой приводили бы к соизмеримым результатам. Только из-за положительной и отрицательной обратной связи причины не имеют одинаковых результатов. Тем не менее, также кажется верным, что мощные петли положительной обратной связи влияют только на небольшое меньшинство входных данных.
* * *
Чем отличники отличаются от посредственных?
Практические занятия бесполезны без полезной обратной связи о результатах.
Петли обратной связи являются частью Решетки ментальных моделей Фарнам-стрит.
Учебные блоки | Управление медицинского образования, Медицинский факультет
Основы медицины (ФОМ)
Этот блок знакомит с фундаментальными науками, лежащими в основе медицинской практики. Блок начинается с углубленного введения в анатомию человека и переходит к принципам клеточного строения и биологии, лежащим в основе основных патологических процессов болезни. Темы наследственности, иммунитета, метаболизма и микробиологии освещаются, чтобы подготовить студентов к последующей курсовой работе по патофизиологии. Этот блок длится с августа по март первого года обучения.
Директора блоков
Синтия Лэнс-Джонс, доктор философии
Ассоциированный профессор
Кафедра нейробиологии
clancej@pitt.edu
Martin C. Schmidt, PhD
Профессор
Кафедра микробиологии и молекулярной генетики
mcs2@pitt. edu
edu
Курсы
- Медицинская анатомия
- Генетика человека
- Метаболизм топлива
- Иммунология в области здравоохранения и болезней
- Фармакология
- Ткани в области здоровья и болезней
- Медицинская микробиология
Блок патофизиологии систем органов (OSP)
Блок систем органов основан на принципах, представленных в блоке фундаментальных наук. Курсы начинаются с тщательного изучения физиологии систем человека, за которым следуют материалы по распространенным патологическим состояниям, соответствующей фармакологии и введение в медицину каждой системы. Разделы состоят из курсов по системам, которые неразрывно связаны по функциям или концептуально связаны по основным физиологическим принципам. Блок систем органов начинается в марте первого года обучения и продолжается в течение всего второго года обучения.
Заведующий отделением
Луи Рапкин, доктор медицины
Клинический директор, онкология
Отделение педиатрии
louis. rapkin@chp.edu
rapkin@chp.edu
Курсы
- Неврология
- Введение в психиатрию
- Гомеостаз жидкости организма: сердечно-сосудистая система
- Гомеостаз биологических жидкостей: почечный
- Гомеостаз жидкости организма: легочный
- Заболевания кожи и опорно-двигательного аппарата
- Пищеварение и питание
- Гематология
- Эндокринный
- Репродуктивная биология и биология развития
- Комплексные тематические исследования
Блок доказательств и обнаружения (E&D)
Навыки чтения биомедицинской литературы, интерпретации данных и оценки клинических исследований находятся в центре внимания курсов этого блока. Курсы объединяют темы клинической экспертизы и научных рассуждений. Студентам напоминают об основных статистических методах и узнают, как клинические испытания, медицинские базы данных и трансляционная медицина являются основой доказательной медицины и ориентированной на пациента помощи. Цель блока E&D — помочь студентам-медикам стать практикующими врачами/исследователями, которые в курсе последних достижений медицины. Курсы этого блока проходят в течение всего первого и второго года обучения.
Цель блока E&D — помочь студентам-медикам стать практикующими врачами/исследователями, которые в курсе последних достижений медицины. Курсы этого блока проходят в течение всего первого и второго года обучения.
Директор блока
Питер Дрейн, доктор философии
Ассоциированный профессор
Кафедра клеточной биологии и физиологии
drop@pitt.edu
Курсы
- Доказательная медицина. Основы
- Доказательная медицина — прикладная
- Расследование и открытие
- Продольный исследовательский проект
Блок для пациентов, врачей и общества (PPS)
Это серия курсов, направленных на развитие навыков, необходимых для эффективного взаимодействия с пациентами, и на углубление понимания учащимися многих ролей, которые врачи играют в обществе, в медицинских профессиях и в жизни своих пациентов. Блок включает интегрированное обучение по сквозным темам, таким как этика, профессионализм, культурная компетентность, качество, безопасность пациентов, системы здравоохранения и правовые аспекты здравоохранения. По возможности в эти курсы включаются контакты с симулированными или реальными пациентами. Студенты начинают иметь непосредственный контакт с пациентами в течение первой недели учебной программы, на курсе «Введение в работу врача». Курсы PPS проводятся в течение всего первого и второго года обучения.
По возможности в эти курсы включаются контакты с симулированными или реальными пациентами. Студенты начинают иметь непосредственный контакт с пациентами в течение первой недели учебной программы, на курсе «Введение в работу врача». Курсы PPS проводятся в течение всего первого и второго года обучения.
Директор блока
Julie Childers, MD, MS
Доцент
Медицинский факультет
childersjw2@upmc.edu
Курсы
- Введение в профессию врача
- Этика, право и профессионализм
- Поведенческая медицина
- Здоровье населения
Введение в блок ухода за пациентами (IPC)
Основы взаимодействия врача и пациента находятся в центре внимания этого блока, который проходит на протяжении всего первого и второго года обучения. Студенты переживают градуированные встречи при разработке интервью с пациентом и проведении основных физических осмотров. По мере роста их навыков включаются элементы физической диагностики отклонений от нормы.