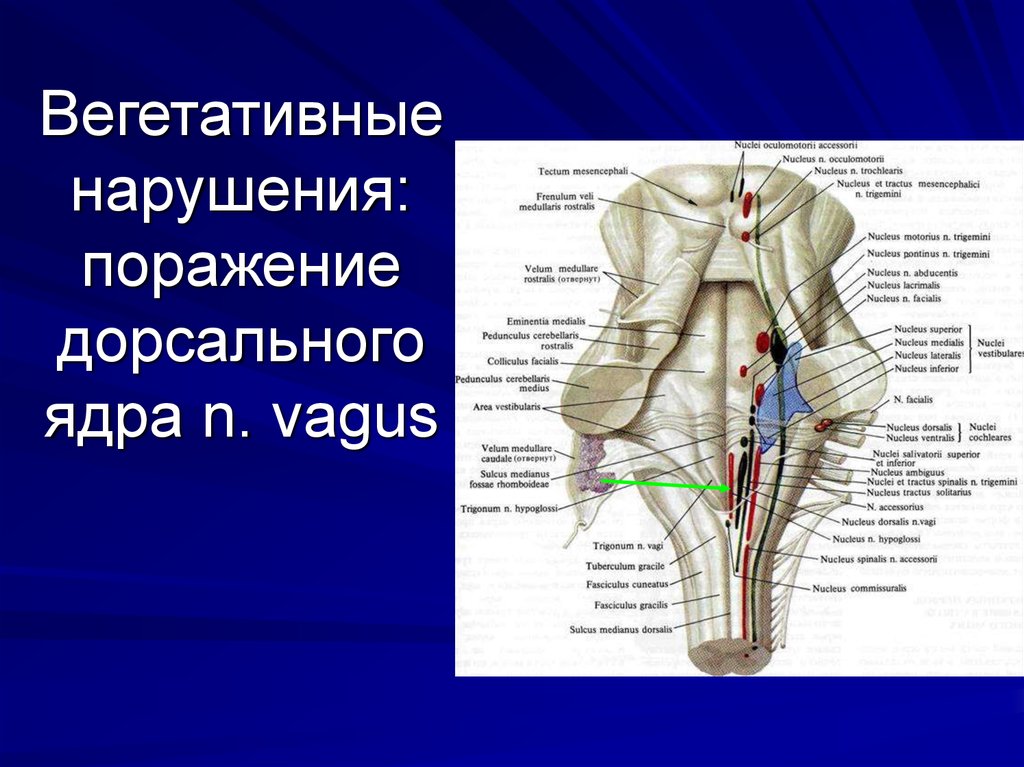
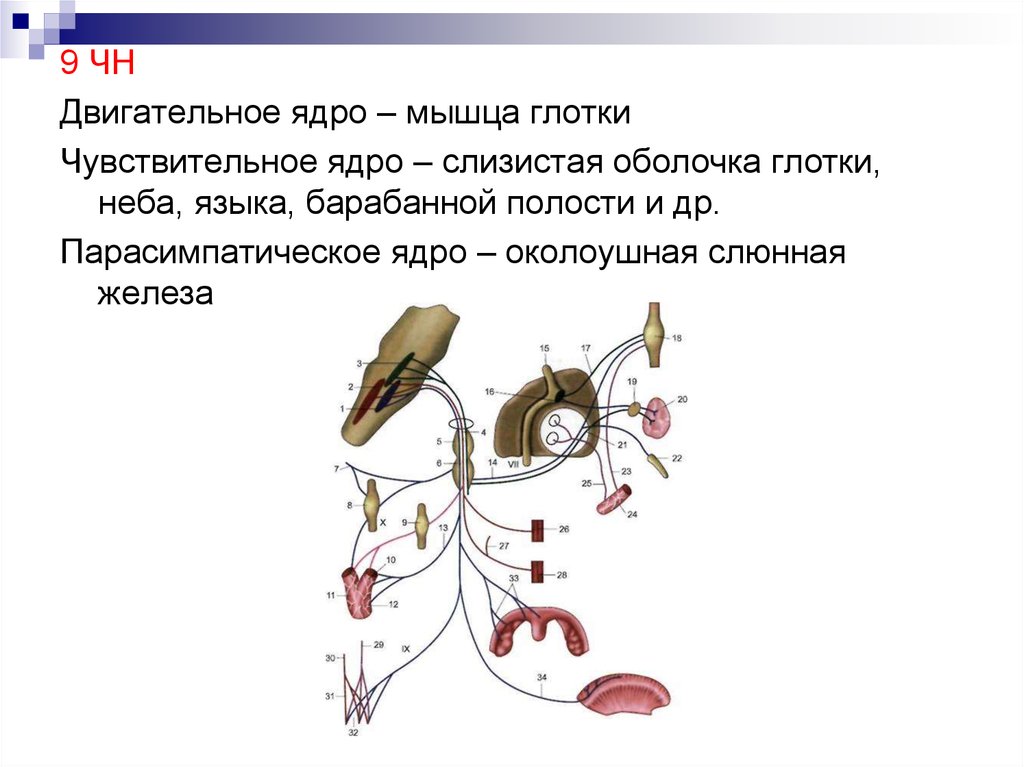
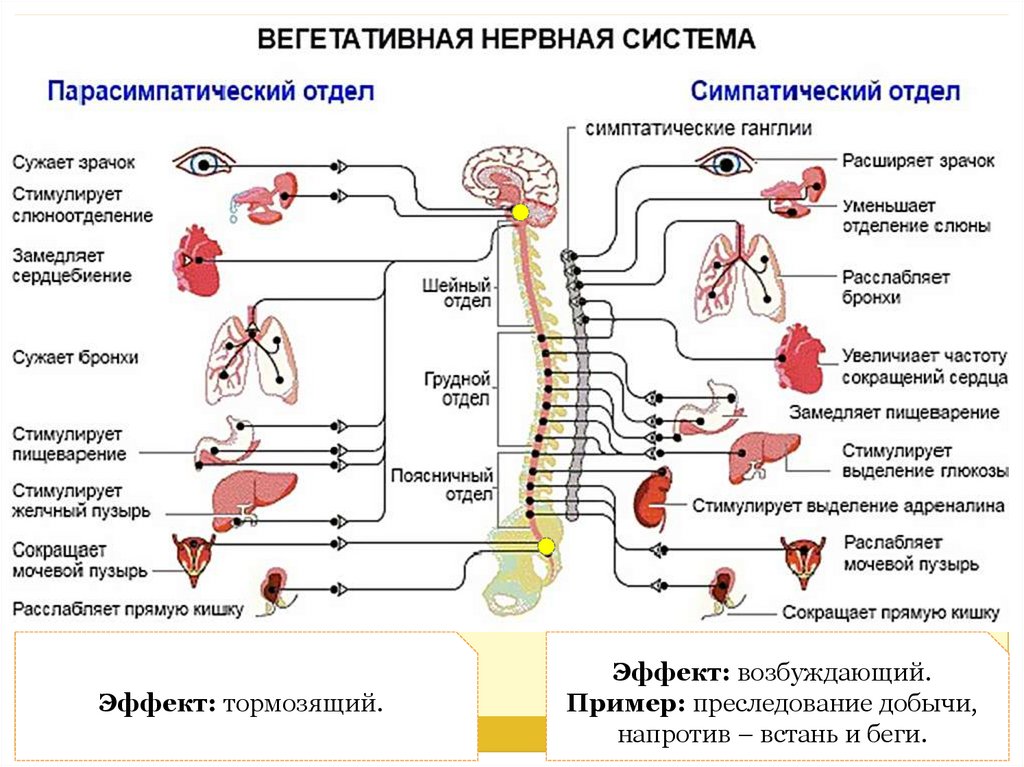
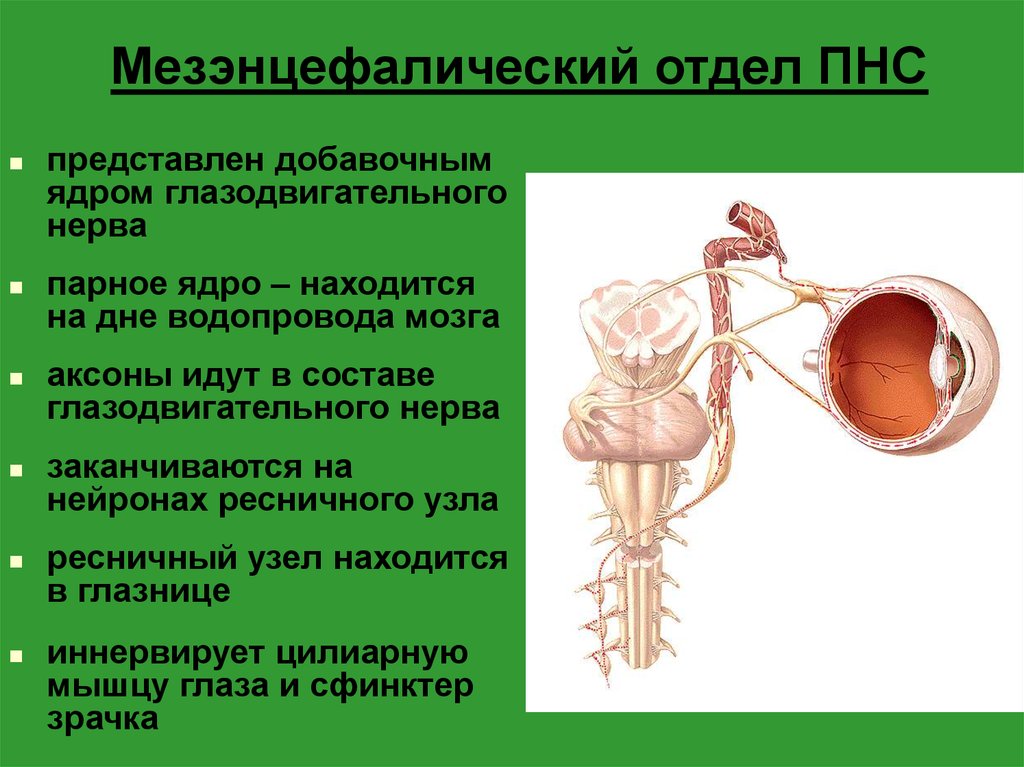
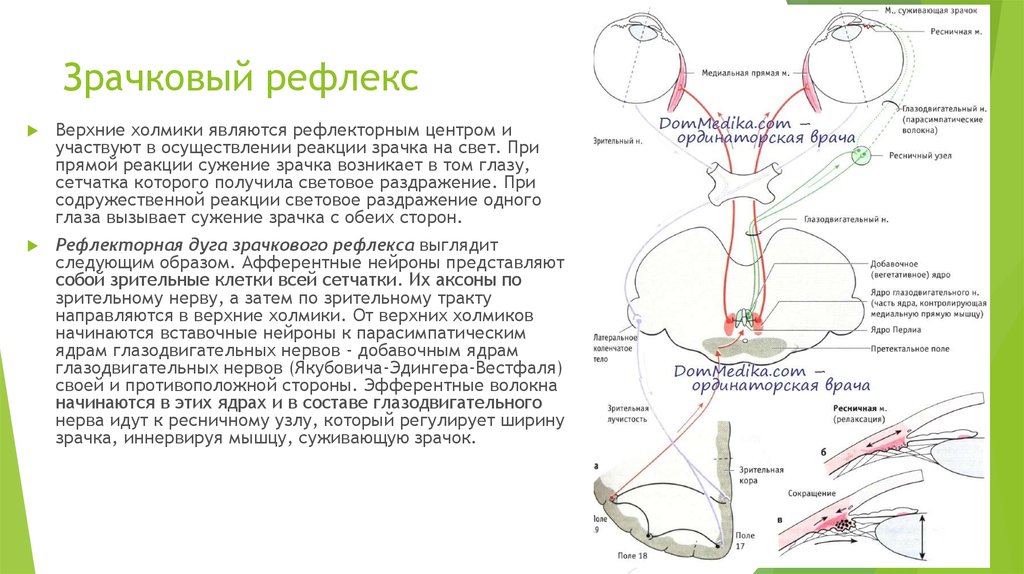
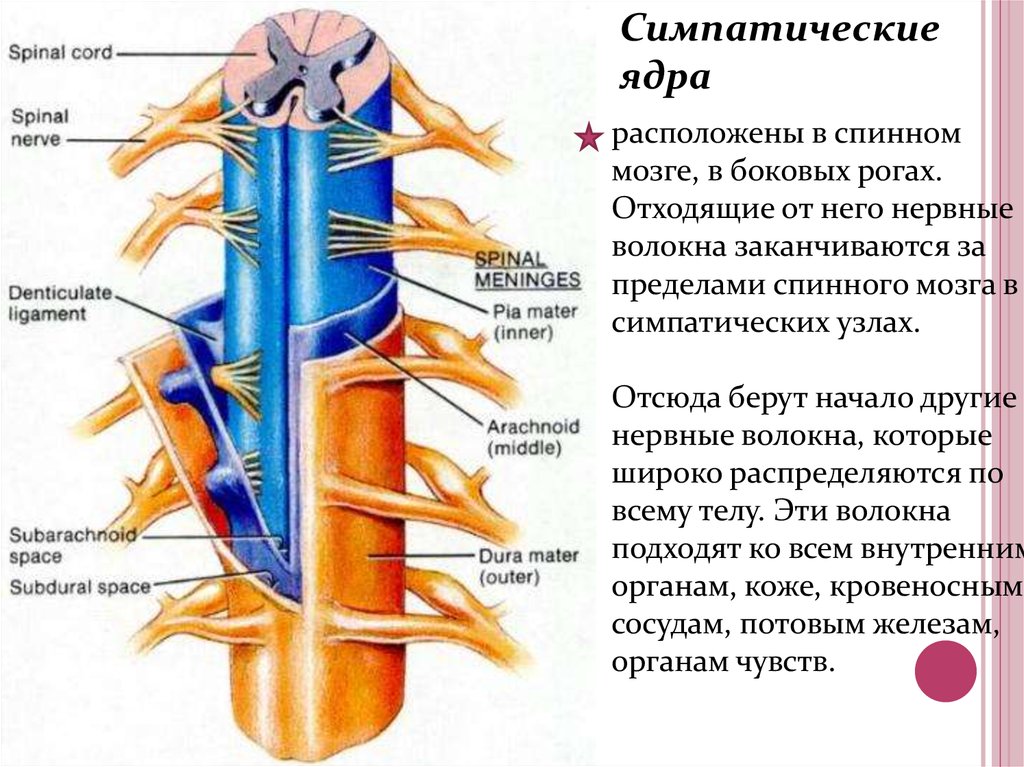
Вегетативное ядро
Вегетативное размножение. Сначала ядро, лежащее в районе перешейка, делится, образуя два ядра, которые расходятся к центрам обеих полукле-ток. После того как ядра передвинулись в по-луклетки, в середине перешейка образуется перегородка, тесно примыкающая своими краями к внутреннему слою оболочки (рис. 254, 1). Затем область перешейка вытягивается, в результате чего две старые полуклетки отделяются одна от другой (рис. 254, 2). Верхняя и нижняя части перешейка, лежащие по обеим сторонам поперечной перегородки, начинают раздуваться в виде пузырька, увеличиваться в размерах, после чего наступает расщепление самой перегородки (рис. 254, 3). Образовавшиеся из раздувшихся частей перешейка молодые полуклетки начинают быстро расти и достигают нормального облика, оставаясь соединенными друг с другом своими вершинами (рис. 254, 4, 5). Когда клетки достигают зрелого возраста, их оболочка, являющая собой временное образование, отбрасывается и отпадает, наподобие линяющей кожи. Тогда клетки окончательно разъединяются и начинают самостоятельное существование.[ …]
Тогда клетки окончательно разъединяются и начинают самостоятельное существование.[ …]
В большинстве случаев ядро клетки-трубки (так называемое «вегетативное» ядро) движется впереди генеративной клетки (или спормиои) и первым доходит до женского гаметофнта. Однако во многих случаях ядро клетки-трубки не только не доходит до женского гаметофита, но даже не попадает в пыльцевую трубку н часто дегенерирует еще в пыльцевом зерне. Кроме того, нередко ядро клетки-трубки идет не впереди генеративной клетки или спермиев, а позади них или между ними, иногда сбоку от них.[ …]
У одноклеточных организмов вегетативное размножение представлено такими формами, как деление, множественное деление и почкование. Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних присуще бактериям и сине-зеленым водорослям (цианобактериям). Напротив, размножение делением бурых и зеленых водорослей, а также одноклеточных животных (саркодовых, жгутиковых и инфузорий) происходит путем митотического деления ядра с последующей перетяжкой цитоплазмы. [ …]
[ …]
Вслед за делением первичного ядра пыльцевого зерна начинается цитокинез с образованием двух клеток: большой — вегетативной с жидкой вакуолизированной цитоплазмой и крупным округлым ядром и меньшей по размеру — генеративной, имеющей густую цитоплазму с высоким содержанием РНК и более плотное, богатое ДНК ядро. Различия в структуре этих двух клеток пыльцевого зерна и их ядер проявляются вскоре после-их образования, Сначала генеративная клетка одной своей стороной плотно прижата к оболочке пыльцевого зерна, но в дальнейшим она растет внутрь полости пыльцевого зерна, принимая веретенообразную форму. При этом поверхность ее соприкосновения с оболочкой пыльцевого зерна уменьшается. Постепенно генеративная клетка отделяется от оболочки пыльцевого зерна и переходит внутрь цитоплазмы вегетативной клетки (рис. 96).[ …]
Б. Нормальное развитие. После деления ядра одно из дочерних ядер перемещается в тот конец клетки, где цитоплазма характеризуется наибольшей плотностью, и становится генеративным. Другое дочернее ядро становится вегетативным.[ …]
Другое дочернее ядро становится вегетативным.[ …]
Как показывают морфометрические подсчеты, в ядрах вегетативных апексов количество гетерохроматина, приходящееся на один поперечный срез ядра, в среднем составляет 89 уел. ед., а эвокированных — 49,9.[ …]
Клетки всех бурых водорослей содержат по одному ядру и большей частью по нескольку мелких дисковидных хлоропластов. Форму хлоропластов иногда используют в качестве признака для разграничения родов. В то же время у бурых наблюдается изменение формы хлоропластов по мере старения клеток.[ …]
Необходимо отметить важную физиологическую роль вегетативного ядра во время формирования спермиев в пыльцевом зерне и пыльцевой трубке. Оно участвует в накоплении нуклео-плазмы в пыльцевом зерне, обеспечивает нормальную жизнедеятельность вегетативной клетки и принимает участие в росте пыльцевой трубки и развитии спермиев.[ …]
В некоторых случаях при прорастании зиготы, а также при вегетативном делении клеток наблюдаются сильные отклонения формы клеток от нормального типа. В результате получаются различные уродливые (тератологические) формы. Наблюдения тератологических форм показало, что они могут возникать от различных причин. Так, при неполном клеточном делении происходит только деление ядра, а разделительная поперечная перегородка между нолуклетками не образуется, в результате чего возникают уродливые клетки, состоящие из трех частей. Крайние части представляют собой нормальные полуклетки, а посередине между ними находится уродливая вздутая часть различной формы. Особенностью некоторых видов является образование аномальных форм с неодинаковыми очертаниями вполне развившихся полуклеток и совершенно нормальной оболочкой. У рода клостериум, например, часто наблюдаются сигмоидные формы, у которых одна полуклетка повернута к другой на 180°.[ …]
В результате получаются различные уродливые (тератологические) формы. Наблюдения тератологических форм показало, что они могут возникать от различных причин. Так, при неполном клеточном делении происходит только деление ядра, а разделительная поперечная перегородка между нолуклетками не образуется, в результате чего возникают уродливые клетки, состоящие из трех частей. Крайние части представляют собой нормальные полуклетки, а посередине между ними находится уродливая вздутая часть различной формы. Особенностью некоторых видов является образование аномальных форм с неодинаковыми очертаниями вполне развившихся полуклеток и совершенно нормальной оболочкой. У рода клостериум, например, часто наблюдаются сигмоидные формы, у которых одна полуклетка повернута к другой на 180°.[ …]
Хлорелла (Chlorella, рис. 211, 5—7) широко распространена и хорошо известна. Ее вегетативные клетки устроены очень просто. Это зеленые шарики, обычно не превышающие в диаметре 15 мкм, протопласт которых имеет один чашевидный, вернее, колоколообразный хлоропласт с одним пиреноидом в утолщенной части. Ядро одно, но в живой клетке без специальной обработки оно невидимо. Покрыт протопласт хорошо выраженной двухконтурной оболочкой. Размножается хлорелла исключительно автоспорами, возникающими обычно по 4—8 в одной клетке. Хлорелла нетребовательна к условиям обитания и способна весьма интенсивно размножаться, поэтому очень широко распространена и встречается почти повсеместно. В водоемах — это типичный планктер, но встречается она и в бентосе, перифитоне и нейсто-не, а также на наземных субстратах и в почве. Она входит в состав лишайников, вступает в симбиоз с разными гидробионтами, образуя так называемые зоохлореллы. Клетка хлореллы оказалась удобным объектом для разносторонних исследований. По количеству посвященных ей работ, в том числе и научно-популярных, хлорелле принадлежит первое место среди всех водорослей нашей планеты. Она стала основным объектом массового культивирования водорослей для практического использования в самых различных направлениях.[ …]
Ядро одно, но в живой клетке без специальной обработки оно невидимо. Покрыт протопласт хорошо выраженной двухконтурной оболочкой. Размножается хлорелла исключительно автоспорами, возникающими обычно по 4—8 в одной клетке. Хлорелла нетребовательна к условиям обитания и способна весьма интенсивно размножаться, поэтому очень широко распространена и встречается почти повсеместно. В водоемах — это типичный планктер, но встречается она и в бентосе, перифитоне и нейсто-не, а также на наземных субстратах и в почве. Она входит в состав лишайников, вступает в симбиоз с разными гидробионтами, образуя так называемые зоохлореллы. Клетка хлореллы оказалась удобным объектом для разносторонних исследований. По количеству посвященных ей работ, в том числе и научно-популярных, хлорелле принадлежит первое место среди всех водорослей нашей планеты. Она стала основным объектом массового культивирования водорослей для практического использования в самых различных направлениях.[ …]
Нормальные пыльцевые зерна хорошо выполнены цитоплазмой и имеют два спермия и вегетативное ядро (см.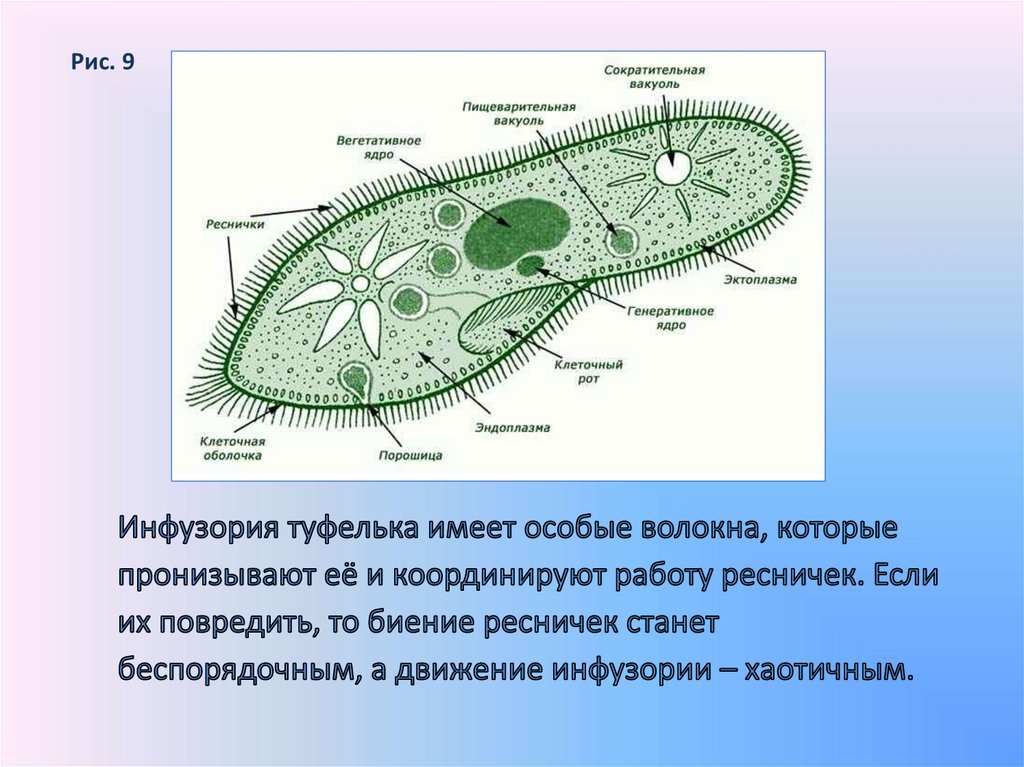 рис. 84).[ …]
рис. 84).[ …]
Второе деление протекает почти без нарушений. Пыльцевые зерна нормального строения: они имеют два спермия, вегетативное ядро и хорошо выполнены цитоплазмой; исключение составляют единичные пыльцевые зерна, которые не окрашиваются ацетокармином. Фертильность растений высокая.[ …]
Процесс дробления начинается с появления бороздки в анимальном полюсе, но глубина ее незначительна. Лежащее под бороздкой ядро делится и расходится в обе стороны от бороздки и таким образом пол , чаются первые два шара дробления, но лишь поверхностного. Через полчаса заметна уже вторая бороздка, перпендикулярная к первой. И так это продолжается далее. Но уже после появления четырех сегментов деление идет глубже, распространяясь на все яйцо, и в то время когда образуется вторая бороздка, первая разделяет все яйцо на две половины или два сегмента. Яйцо остается таковым довольно долго, пока наверху не образуются восемь сегментов, после чего начинаешься деление в экваториальном направлении, тогда как .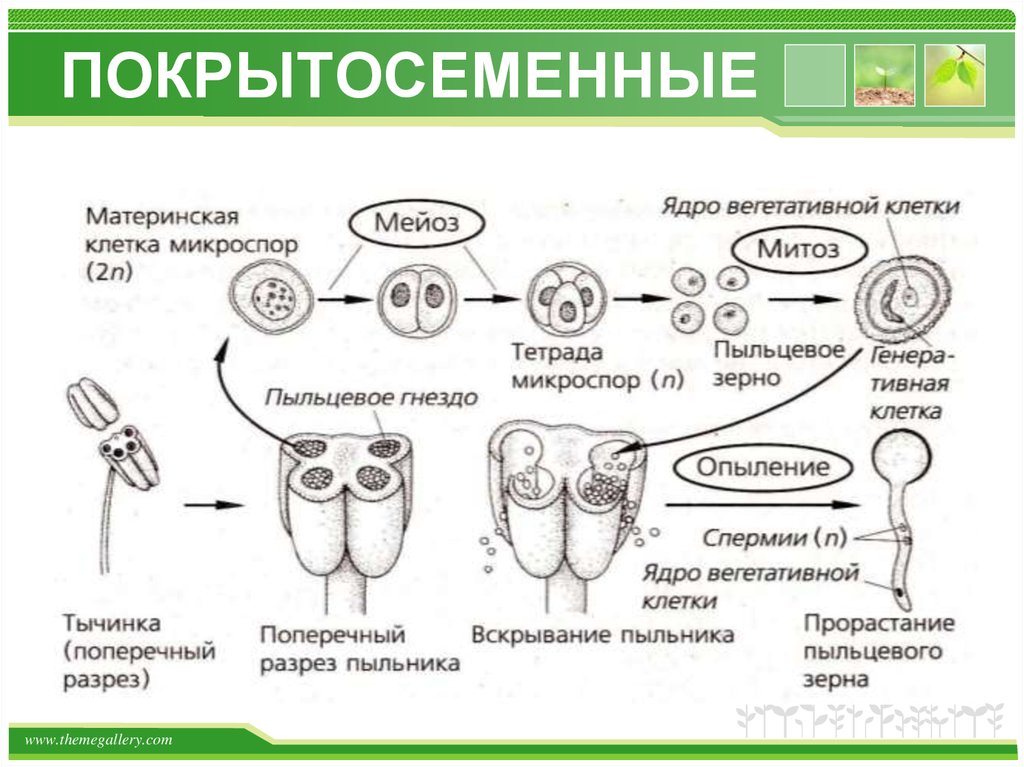 доселе все бороздки и, стало быть, деление шли в меридианальном направлении. Эти бороздки вглубь не доходят до конца, они только поверхностные, так что внутри яйца вся масса его еще нераздельна. Полное же отделение сегментов происходит лишь после появления наружных и внутренних экваториальных бороздок. Хотя меридианальные бороздки постепенно удлиняются к вегетативному полюсу яйца, но не все с одинаковой скоростью, и поэтому в то время, когда в анимальном полюсе уже 12 сегментов, в вегетативном — ■всего шесть. Наконец, после появления экваториальных борозд сегментация •продолжается в обоих направлениях, и вскоре мы видим яйцо состоящим из массы сегментов, из коих вверху лежат более мелкие и сильнее пигментированные, чем внизу.[ …]
доселе все бороздки и, стало быть, деление шли в меридианальном направлении. Эти бороздки вглубь не доходят до конца, они только поверхностные, так что внутри яйца вся масса его еще нераздельна. Полное же отделение сегментов происходит лишь после появления наружных и внутренних экваториальных бороздок. Хотя меридианальные бороздки постепенно удлиняются к вегетативному полюсу яйца, но не все с одинаковой скоростью, и поэтому в то время, когда в анимальном полюсе уже 12 сегментов, в вегетативном — ■всего шесть. Наконец, после появления экваториальных борозд сегментация •продолжается в обоих направлениях, и вскоре мы видим яйцо состоящим из массы сегментов, из коих вверху лежат более мелкие и сильнее пигментированные, чем внизу.[ …]
Нормальная, хорошо окрашивающаяся пыльца, как у родительских, так и гибриднх форм, является трехъядерной, в ней ясно видны одно вегетативное и два генеративных ядра.[ …]
Размеры ядер даже в пределах одного таллома могут существенно колебаться. Так, у одного вида бриопсиса (Bryopsis halymeniae) из сифоновых ядра спорангия имеют диаметр, равный 30—45 мкм, а ядра вегетативных клеток таллома — 3,0—4,5 мкм. У другого вида бриопсиса (Bryopsis hypnoides) ядра в проростке зиготы оказываются крупнее ядра зиготы почти в 1700 раз. У водяной сеточки в молодых клетках ядра не превышают в диаметре 2,8 мкм, а в старых иногда достигают 16 мкм. Интересно, что перед делением диаметр ядер обычно увеличивается более чем в 2 раза. В этот же период происходит и его перемещение, как правило, в центр (эдогониум) или в переднюю часть клетки (эвглена).[ …]
У другого вида бриопсиса (Bryopsis hypnoides) ядра в проростке зиготы оказываются крупнее ядра зиготы почти в 1700 раз. У водяной сеточки в молодых клетках ядра не превышают в диаметре 2,8 мкм, а в старых иногда достигают 16 мкм. Интересно, что перед делением диаметр ядер обычно увеличивается более чем в 2 раза. В этот же период происходит и его перемещение, как правило, в центр (эдогониум) или в переднюю часть клетки (эвглена).[ …]
Прежде всего резко различны происхождение и характер запасающих тканей. Запасающая ткань в сомони голосеменных представляет собой вегетативную часть женского гамето-фита и имеет, следовательно, гаплоидный характер. Эндосперм цветковых формируется, как ужо отмечалось, в результате слияния одного из спермиев с диплоидным ядром центральной клетки зародышевого мешка и у подавляющего большинства растений имеет трип-лоидный характер. В семени некоторых цветковых растений сохраняется в большей или меньшей степени ткань нуцеллуса, преобразующаяся в диплоидную запасающую ткань — перисперм. [ …]
[ …]
В люминесцентном микроскопе при окраске акридиновым оранжевым (люминесцирующий краситель) наблюдается многокрасочное изображение. Их ядра ярко люминесцируют зеленым светом (табл. 32, схема 1). Ядерные тяжи в спорогенной зоне сначала зеленые, затем окружаются веществом с красной флюоресценцией. Далее флюоресцирующая красная зона принимает овальную форму, а хроматиновый, расположенный в центре тяж светится желтым светом. При созревании спора резко меняет цвет и начинает светиться ярким зеленым светом. У зрелых спор с хорошо выраженным кортексом (корой) сердцевина споры темная, зеленым светится только оболочка. Меняется также и цвет цитоплазмы материнской клетки (спорангия). Сначала она слабо-зеленого цвета, затем, с момента оформления проспоры, начинает светиться оранжевым светом.[ …]
Химический состав и роль споровых структур. Сердцевина споры, окруженная слоем кортекса, представляет собой протопласт с собственной мембраной, ядром и цитоплазмой (схема 3 на табл. 32). Сердцевина у зрелых спор является покоящейся вегетативной клеткой. Она характеризуется очень низким уровнем метаболизма. Хотя в ней имеются все необходимые ферменты, их активность каким-то образом подавлена.[ …]
Сердцевина у зрелых спор является покоящейся вегетативной клеткой. Она характеризуется очень низким уровнем метаболизма. Хотя в ней имеются все необходимые ферменты, их активность каким-то образом подавлена.[ …]
Перед самым оплодотворением сперматоген-ная клетка делится, образуя два обычно одинаковых спермия. При оплодотворении в архего-нии вступают не только спермии, но и вегетативное ядро пыльцевой трубки, а также клетка-ножка. Если спермии различны по величине, то с ядром яйцеклетки сливается более крупный.[ …]
Размножение простейших происходит как бесполым, так и половым путем. Бесполое размножение заключается в делении орга-нияма на две половины, причем ему предшествует деление ядра. Иногда деление тела и деление ядра происходят одновременно. Размножение половым путем осуществляется с помощью синга-мии (слияния двух гамет), конъюгации (обмена гаметическими ядрами) и аутогамии, заключающейся в формировании гаплоидных ядер и слияния их в синкарионы. Некоторые простейшие размножаются одновременно бесполым и половым путем. Например, в размножении малярийных плазмодиев бесполая фаза встречается, когда они находятся в организме позвоночных, половая, когда находятся в организме комаров. Установлено, что Еир1оЪеа га11«т продуцирует ферромоны полипептидной природы, которые индуцируют как половое, так и вегетативное размножение простейших этого вида.[ …]
Например, в размножении малярийных плазмодиев бесполая фаза встречается, когда они находятся в организме позвоночных, половая, когда находятся в организме комаров. Установлено, что Еир1оЪеа га11«т продуцирует ферромоны полипептидной природы, которые индуцируют как половое, так и вегетативное размножение простейших этого вида.[ …]
На месте будущего бесполого концептакула из нижних клеток периталлия образуются, чередуясь с обычными вегетативными, фертильные (плодоносные) нити. Каждая такая пить (у мелобезии она может быть очень короткой) оканчивается молодым тетраспорангием, над которым еще в самом начале его образования формируется маленькая клетка с густым содержимым. Вегетативные нити, окружающие спорангии, продолжают расти, но по мере того, как спорангий вытягивается в длину, они дегенерируют и исчезают и таким образом образуется полость концептакула. Нетронутыми остаются только верхние части стерильных нитей, расположенные выше тетраспорангиев. Эта ткань формирует крышу концептакула. Мелкие клетки над тет-раснорангиями постепенно теряют цитоплазму и ядро и, оказываясь расположенными среди нитей крыши концентакула, приобретают вид слизистых пробок, закрывающих многочисленные поры, через которые выходят зонально поделенные спорангии, выдавливая при этом пробки (рис. 172, 4).[ …]
Мелкие клетки над тет-раснорангиями постепенно теряют цитоплазму и ядро и, оказываясь расположенными среди нитей крыши концентакула, приобретают вид слизистых пробок, закрывающих многочисленные поры, через которые выходят зонально поделенные спорангии, выдавливая при этом пробки (рис. 172, 4).[ …]
Он слагается из нескольких этапов: слияния ядер гетеро-кариона и образования диплоидного гетерозиготного ядра; размножения таких гетерозиготных ядер в гетерокариотическом мицелии вместе с гаплоидными; митотической рекомбинации (при митозе) во время размножения диплоидных ядер; вегетативной гаплоидизации диплоидных ядер в результате утраты ими хромосом. Парасексуальный процесс известен сейчас у многих несовершенных грибов, однако его наблюдают у них только в условиях лаборатории; какова же его роль в естественных условиях обитания, пока не установлено.[ …]
Тот факт, что неэквивалентное деление вызывается полярными различиями в цитоплазме, можно проиллюстрировать на примере развития пыльцевых зерен, у которых веретено деления ориентируется таким образом, что одно из дочерних ядер перемещается в конец клетки с более густой цитоплазмой и становится генеративным, тогда как дочернее ядро перемещается в зону клетки с менее густой цитоплазмой и становится вегетативным (рис. 1.14,Б). При определенных обстоятельствах плоскость веретена может оказаться ориентированной поперек оси материнской пыльцевой клетки, и тогда две образующиеся клетки будут одинаковыми, а дальнейшее развитие пыльцевого зерна нарушается.[ …]
1.14,Б). При определенных обстоятельствах плоскость веретена может оказаться ориентированной поперек оси материнской пыльцевой клетки, и тогда две образующиеся клетки будут одинаковыми, а дальнейшее развитие пыльцевого зерна нарушается.[ …]
После оплодотворения икринки набухают, в них происходит обводнение желтка с образованием вокруг него перивителлинового пространства, заполненного жидкостью. Перивителлиновое пространство образуется и в неоплодотворенных икринках, после чего они обычно погибают. Яйцеклетки костистых рыб относятся к те-лолецитальному типу. В них ядро и плазма клетки находятся на анимальном полюсе, а желток — на противоположной части (вегетативном полюсе). В результате дробление охватывает только верхнюю часть клетки, начинаясь с образования бластодиска. Дальше развитие идет по общей схеме: образование бластулы, морулы, гаст-рулы. При гаструляции клетки анимального полюса окружают жел-ток с образованием двух зародышевых листков (экто- и энтодермы). Далее развитие переходит в стадию органогенеза, подвижного эмбриона, заканчиваясь выклевом предличинок. Продолжительность инкубации икры у разных видов рыб различная и зависит от температуры воды. Например, при оптимальных температурах (20— 22 °С) икра карпа развивается в течение 3-4 сут, а радужной форели при температуре около 10 °С — в течение 45-60 сут.[ …]
Продолжительность инкубации икры у разных видов рыб различная и зависит от температуры воды. Например, при оптимальных температурах (20— 22 °С) икра карпа развивается в течение 3-4 сут, а радужной форели при температуре около 10 °С — в течение 45-60 сут.[ …]
В процессе развития происходит постепенная диффереици-ровка органов и тканей, что приводит к возникновению большого разнообразия типов клеток. Однако не все гены, входящие в состав генома, активны в каждый данный момент и в каждой данной части растения. Так, гены, контролирующие развитие цветков, обычно не экспрессируются ни у зародышей, ни во время чисто вегетативной фазы развития. Вместе с тем мы знаем, что клетки таких вегетативных органов, как лист, содержат гены для развития цветков, поскольку из клеток листьев некоторых видов могут регенерировать новые растения, способные к цветению. Следовательно, дифференцировка у растений не связана с генетическими (т. е. наследственными) различиями между ядрами различного типа клеток и тканей. В таком случае она должна определяться различиями в экспрессии генов в тех или иных частях растения или на тех или иных стадиях его жизненного цикла.[ …]
В таком случае она должна определяться различиями в экспрессии генов в тех или иных частях растения или на тех или иных стадиях его жизненного цикла.[ …]
Род дербезия (ОегЬез1а) включает нитчатые водоросли, распространенные не только в тропических, но и в умеренных морях. Слоевище их состоит из стелющихся и вертикальных разветвленных нитей. В месте отхождения ветвей, так же как и в основании спорангиев, образуются своеобразные перегородки. Они могут быть простыми или двойными, и тогда между ними остается короткий сегмент с хлоропластами и ядрами. Вегетативное размножение, кроме фрагментации, заключается в образовании на вершине ветвей своеобразных выводковых почек, которые, отделяясь, превращаются в новые растения. Для бесполого размножения служат зооспоры (изредка апланоспоры). Это крупные шаровидные клетки с венцом жгутиков на переднем конце (рис. 230, 2—3), содержащие несколько ядер и хлоропластов. При их образовании обычные вегетативные веточки меняют свою форму на шаровидную или грушевидную, и, отделяясь от слоевища перегородкой, превращаются в спорангии. [ …]
[ …]
Второе деление мейоза также протекает с большими отклонениями от нормы, особенно это сказывается на образовании микроядер, причем некоторые из них образованы многими хромосомами. Пыльца пустая, абортивная, но среди пустой пыльцы наблюдаются отдельные пыльцевые зерна, выполненные цито-тшазмой. При просмотре пыльцы под микроскопом в ацетокармине такие пыльцевые зерна хорошо красятся и в них можно видеть вегетативное ядро и два спермия. Но эти нормальные пыльцевые зерна остаются недеятельными, так как пыльники с малым количеством нормальной пыльцы кожистые и не растрескиваются. При вскрытии их пинцетом или препаровальной иглой и нанесении содержимого на рыльце цветков пшеницы завязываются семена, но в небольшом числе. Однако у отдельных гибридных растений иногда наблюдается растрескивание пыльников, в которых находится до 60% нормально выполненных цитоплазмой, хорошо окрашивающихся в ацетокармине пыльцевых зерен.[ …]
У одних представителей этого семейства наблюдается типичный для высших аскомицетов половой процесс (например, у Мопавсив ригригеив). Однако у многих эуроциевых происходит его морфологическая редукция. В этих случаях в дикарионы объединяются ядра самого аскогона.[ …]
Однако у многих эуроциевых происходит его морфологическая редукция. В этих случаях в дикарионы объединяются ядра самого аскогона.[ …]
Женский репродуктивный орган у красных водорослей — оогоний называют карпого-ном. Это особая клетка, густо заполненная цитоплазмой и лишенная хлоропласта. Карпогон отличается весьма характерной, одинаковой для всех флоридеевых формой (рис. 160, 1). Нижняя, или брюшная, часть клетки имеет конусовидную форму, верхняя вытягивается, образуя более или менее длинный трубчатый вырост. Благодаря трихогине карпогон легко отличить от остальных клеток слоевища. Ядро карпогона, как правило, находится в брюшной части. Трихогина может быть короткой и кеглевидной или длинной, узкой и спирально закрученной. У бангиевых собственно трихогина еще отсутствует, а иногда карпогоны имеют лишь короткие сосочкообразные выросты — прообраз будущей трихогины.[ …]
Дазикладовые образуют хорошо очерченную естественную группу морских водорослей, насчитывающую около 10 родов. Их слоевище, представляющее собой одну гигантскую клетку, имеет весьма сложную морфологию. В основе их организации лежит радиальная симметрия. У всех дазикладовых слоевище состоит из не-разветвленной центральной оси, покрытой мутовками коротких ветвей и прикрепляющейся к субстрату системой разветвленных ризоидов. За редкими исключениями, для этих водорослей характерно более или менее полное обызвествление стенок слоевища. В отличие от других водорослей класса сифоновых слоевище у дазикладовых одноядерное. В вегетативном состоянии ядро располагается в ризоидах. Перед гаметообразованием первичное ядро подвергается многочисленным делениям и дочерние ядра мигрируют в развивающиеся гаметангии. Хлоропласты многочисленные, дисковидные или эллипсоидные; пиреноид есть или отсутствует.[ …]
Их слоевище, представляющее собой одну гигантскую клетку, имеет весьма сложную морфологию. В основе их организации лежит радиальная симметрия. У всех дазикладовых слоевище состоит из не-разветвленной центральной оси, покрытой мутовками коротких ветвей и прикрепляющейся к субстрату системой разветвленных ризоидов. За редкими исключениями, для этих водорослей характерно более или менее полное обызвествление стенок слоевища. В отличие от других водорослей класса сифоновых слоевище у дазикладовых одноядерное. В вегетативном состоянии ядро располагается в ризоидах. Перед гаметообразованием первичное ядро подвергается многочисленным делениям и дочерние ядра мигрируют в развивающиеся гаметангии. Хлоропласты многочисленные, дисковидные или эллипсоидные; пиреноид есть или отсутствует.[ …]
Настоящие волоски багрянок следует отличать от волосовидных ветвей или ложных волосков. У видов с однорядным нитчатым слоевищем можно видеть, как конечные клетки боковых веточек удлиняются и обесцвечиваются, приобретая волосовидное строение (рис. 151,7). Это и есть ложные волоски. Настоящие волоски красных водорослей делятся на два типа: одноклеточные и многоклеточные. Одноклеточные волоски никогда не ветвятся. У однорядных нитчатых форм они образуются из верхушечных клеток ветвей, у многорядных — из поверхностных клеток коры. Клетка будущего волоска отделяется от материнской клетки поперечной перегородкой и сильно вытягивается, достигая в длину нередко миллиметра и более (рис. 156). Она содеряшт ядро и небольшое количество цитоплазмы. Хлоропласт по мере роста волоска исчезает, и волоски становятся бесцветными. Обычно материнская клетка волоска ничем не отличается от соседних вегетативных клеток, но иногда она намного крупнее и остается хорошо заметной после отпадения волоска. У кораллиновых волоски не являются самостоятельными клетками, а представляют собой только выросты специальных клеток, от которых опи не отделяются перегородкой.[ …]
151,7). Это и есть ложные волоски. Настоящие волоски красных водорослей делятся на два типа: одноклеточные и многоклеточные. Одноклеточные волоски никогда не ветвятся. У однорядных нитчатых форм они образуются из верхушечных клеток ветвей, у многорядных — из поверхностных клеток коры. Клетка будущего волоска отделяется от материнской клетки поперечной перегородкой и сильно вытягивается, достигая в длину нередко миллиметра и более (рис. 156). Она содеряшт ядро и небольшое количество цитоплазмы. Хлоропласт по мере роста волоска исчезает, и волоски становятся бесцветными. Обычно материнская клетка волоска ничем не отличается от соседних вегетативных клеток, но иногда она намного крупнее и остается хорошо заметной после отпадения волоска. У кораллиновых волоски не являются самостоятельными клетками, а представляют собой только выросты специальных клеток, от которых опи не отделяются перегородкой.[ …]
При анализе особенностей прохождения ежегодно повторяющихся половых циклов в водоемах различных широт было обнаружено, что в южных водоемах, в частности Куре и Кубани, самки судака зимуют с незрелыми половыми клетками. Процесс интенсивного вителлогенеза у них, в отличие от судака, обитающего в более северных водоемах, протекает незадолго до периода размножения судака в водоеме. Анализ полового цикла самок курин-ского судака показал, что особи в этом водоеме зимуют с половыми железами в III стадии зрелости яичников. На гистологических срезах яичников судака из р. Куры, которые были получены в ноябре, можно было видеть, что основная масса ооцитов в яйцене-сущих пластинках вступает лишь в начальный период трофоплаз-матического роста и находится в различных фазах вакуолизации и, в частности, в конечной фазе этого процесса, и лишь некоторые яйцеклетки в самом начале отложения питательных веществ (рис. 31, Б). В преднерестовый период ооциты генерации текущего года находятся в однородных фазах зрелости (рис. 31, А). При этом, так же как и у других исследованных нами представителей окуневых, у судака происходят изменения в строении и расположении запасных питательных веществ в ооцитах, обнаружены изменения в оболочках, ядре и т, д.
Процесс интенсивного вителлогенеза у них, в отличие от судака, обитающего в более северных водоемах, протекает незадолго до периода размножения судака в водоеме. Анализ полового цикла самок курин-ского судака показал, что особи в этом водоеме зимуют с половыми железами в III стадии зрелости яичников. На гистологических срезах яичников судака из р. Куры, которые были получены в ноябре, можно было видеть, что основная масса ооцитов в яйцене-сущих пластинках вступает лишь в начальный период трофоплаз-матического роста и находится в различных фазах вакуолизации и, в частности, в конечной фазе этого процесса, и лишь некоторые яйцеклетки в самом начале отложения питательных веществ (рис. 31, Б). В преднерестовый период ооциты генерации текущего года находятся в однородных фазах зрелости (рис. 31, А). При этом, так же как и у других исследованных нами представителей окуневых, у судака происходят изменения в строении и расположении запасных питательных веществ в ооцитах, обнаружены изменения в оболочках, ядре и т, д. В частности, по мере развития ооцита, происходит слияние мелких жировых вакуолей в более крупные, а затем образуется одна жировая капля, расположенная ближе к анимальному полюсу яйцеклетки. Ядро ооцита перед овуляцией смещается к вегетативному полюсу. Желток образует сплошную гомогенную массу. Многие из перечисленных особенностей развития яйцеклеток являются характерными для всего семейства в целом и подробно были описаны нами на примере развития половых клеток у окуня, как представителя семейства с наиболее четкими и последовательными картинами всего прохождения гаметогенеза и половых циклов.[ …]
В частности, по мере развития ооцита, происходит слияние мелких жировых вакуолей в более крупные, а затем образуется одна жировая капля, расположенная ближе к анимальному полюсу яйцеклетки. Ядро ооцита перед овуляцией смещается к вегетативному полюсу. Желток образует сплошную гомогенную массу. Многие из перечисленных особенностей развития яйцеклеток являются характерными для всего семейства в целом и подробно были описаны нами на примере развития половых клеток у окуня, как представителя семейства с наиболее четкими и последовательными картинами всего прохождения гаметогенеза и половых циклов.[ …]
Одноклеточные организмы — что это, определение и ответ
Подцарство Простейшие (одноклеточные)
Подцарство Простейшие включает в себя животных, тело которых состоит из одной клетки. Клетка – целый организм, роль органов выполняют органоиды.
Разнообразие Простейших
Тип Жгутиковые
Тип Саркодовые
Тип Инфузории
Тип Споровики
I. Тип Жгутиковые
Тип Жгутиковые
Представитель: Эвглена Зеленая
Строение и описание жизнедеятельности:
Эвглена, как типичный представитель данного типа, осуществляет передвижение за счет движения (вращения) ЖГУТИКА
На переднем конце тела расположен клеточный рот и светочувствительный глазок (=стигма)
В передней части тела мы также можем увидеть сократительную вакуоль (органоид выделения жидких продуктов обмена)
В цитоплазме содержатся зеленые ХЛОРОПЛАСТЫ, несущие зеленый пигмент – хлорофилл
Питание = может питаться как готовыми органическими веществами, так и собственной органикой, синтезированной в процессе фотосинтеза
Есть плотная оболочка, форма тела постоянна (НЕ КЛЕТОЧНАЯ СТЕНКА)
Размножается простым делением надвое
Особенности данного организма:
Эвглена сочетает в себе признаки:
Эвглена зеленая
Лямблия, трипаносома
II. Тип Саркодовые
Тип Саркодовые
Представитель: Амеба Обыкновенная
Строение и описание жизнедеятельности:
Непостоянная форма тела из-за отсутствия плотной оболочки, способны образовывать ПСЕВДОПОДИИ, или ЛОЖНОНОЖКИ
При неблагоприятных условиях образует ЦИСТУ
Питается фагоцитозно, нет клеточного рта
Размножается простым делением надвое
Строение Амебы обыкновенной
Фораминиферы
Особенности:
обитатели моря;
подавляющее большинство фораминифер образует известковую раковину, служащую экзоскелетом. Через отверстие раковины во внешнюю среду могут выпячиваться ложноножки;
раковины фораминифер участвуют в образовании значительной части морских отложений (осадочных пород).
Разнообразие фораминифер
III. Тип Инфузории (Ресничные)
Тип Инфузории (Ресничные)
Представитель: Инфузория туфелька
Строение и описание жизнедеятельности:
Большое количество РЕСНИЧЕК
Есть клеточный рот, глотка
Удаление непереваренной пищи с помощью ПОРОШИЦЫ
Наличие двух сократительных вакуолей (2 «солнышка»)
КОНЪЮГАЦИЯ = половой процесс
Особенности данного организма:
1. Два ядра
| Макронуклеус (большое ядро) | Микронуклеус (маленькое ядро) |
|---|---|
Вегетативное ядро = Отвечает за все процессы жизнедеятельности, кроме полового процесса | Генеративное ядро = Отвечает за половой процесс |
Строение Инфузории-туфельки
2. Конъюгация – половой процесс одноклеточных животных, при котором те обмениваются своим наследственным материалом.
Конъюгация – половой процесс одноклеточных животных, при котором те обмениваются своим наследственным материалом.
Размножение (бесполое и половое)
Сближение двух инфузорий (МИ – микронуклеус; МА – макронуклеус) и образование между ними цитоплазматического мостика.
Первое деление микронуклеуса (как результат, образование двух ядер).
Второе деление микронуклеуса (как результат, образование четырех ядер).
Трое из четырёх образовавшихся микронуклеусов погибают. Также погибает макронуклеус.
Третье деление микронуклеуса (как результат, образование двух ядер).
Обмен ядрами (одно из двух образовавшихся ядер переходит к клетку другой инфузории).
Слияние двух новых микронуклеусов в одно ядро.
Деление ядра надвое.
Из двух имеющихся ядер, одно становится макронуклеусом, другое – микронуклеусом.
IV. Тип Споровики
Тип Споровики
Представитель: Малярийный плазмодий
На экзамене вас будут много спрашивать про жизненный цикл малярийного плазмодия!
Основной хозяин = хозяин, в котором происходит ПОЛОВОЕ размножение
Промежуточный = БЕСПОЛОЕ
Малярийный плазмодий:
Основной хозяин = КОМАР!
Промежуточный хозяйн = ЧЕЛОВЕК!
Вызывает малярию:
Возбудитель = малярийный плазмодий
Переносчик = малярйный комар
Цикл развития малярийного плазмодия:
В организм человека возбудители малярии попадают вместе со слюной заражённого комара в виде тонких веретеновидных клеточек – так называемых спорозоитов
Кровеносная система разносит спорозоиты по всему телу, и часть их через печёночную артерию попадает в печень и там внедряется в печёночные клетки
В клетках печени паразиты разрастаются, а затем распадаются на мелкие дольки – мерозоиты, которые попадают в общее кровяное русло и уже в таком виде внедряются в красные кровяные тельца – эритроциты
Он заполняет собой весь объем поражённого им кровяного тельца, а затем дробится (шизогония = бесполое размножение) сразу на мелкие дольки – мерозоиты.
 Тогда кровяные тельца, в которых развивались паразиты, окончательно разрушаются, молодые мерозоиты попадают в кровяную жидкость, а затем проникают в новые кровяные тельца
Тогда кровяные тельца, в которых развивались паразиты, окончательно разрушаются, молодые мерозоиты попадают в кровяную жидкость, а затем проникают в новые кровяные тельцаЛихорадка
Укус комара, паразит попадает в кишечник, где происходит половое размножение.
Внедрение паразита в слюнные железы комара
 Тогда кровяные тельца, в которых развивались паразиты, окончательно разрушаются, молодые мерозоиты попадают в кровяную жидкость, а затем проникают в новые кровяные тельца
Тогда кровяные тельца, в которых развивались паразиты, окончательно разрушаются, молодые мерозоиты попадают в кровяную жидкость, а затем проникают в новые кровяные тельцаЖизненный цикл малярийного плазмодия
Внимание! Лихорадка наступает по причине разрушения эритроцитов.
Токсоплазма
Токсоплазма – внутриклеточный паразит. Токсоплазмоз протекает достаточно легко, однако у лиц с иммунодефицитом и беременных женщин последствия могут быть фатальными (у беременных плод может погибнуть).
Токсоплазма
Опосредованное LINC-комплексом позиционирование вегетативного ядра участвует в передаче сигналов кальция и АФК в пыльцевых трубках Arabidopsis
. 2020 дек;11(1):149-163.
дои: 10.1080/19491034.2020.1783783.
Морган Мозер 1 , Эндрю Киркпатрик 1 , Норман Рейд Гроувс 1 2 , Ирис Мейер 1 2 3
Принадлежности
- 1 Кафедра молекулярной генетики, Университет штата Огайо, Колумбус, Огайо, США.
- 2 Центр прикладных наук о растениях, Университет штата Огайо, Колумбус, Огайо, США.
- 3 Центр биологии РНК, Университет штата Огайо, Колумбус, Огайо, США.
- PMID: 32631106
- PMCID: PMC7529407
- DOI: 10.1080/19491034.2020.1783783
Бесплатная статья ЧВК
Морган Мозер и др. Ядро. 2020 Декабрь
Бесплатная статья ЧВК. 2020 дек;11(1):149-163.
дои: 10.1080/19491034.2020.1783783.
Авторы
Морган Мозер 1 , Эндрю Киркпатрик 1 , Норман Рейд Гроувс 1 2 , Ирис Мейер 1 2 3
Принадлежности
- 1 Кафедра молекулярной генетики, Университет штата Огайо, Колумбус, Огайо, США.
- 2 Центр прикладных наук о растениях, Университет штата Огайо, Колумбус, Огайо, США.
- 3 Центр биологии РНК, Университет штата Огайо, Колумбус, Огайо, США.
- PMID: 32631106
- PMCID: PMC7529407
- DOI: 10.1080/19491034.2020.1783783
Абстрактный
Движение и расположение ядер играют роль в процессах развития на протяжении всей жизни. Ядерное движение и позиционирование опосредуются, прежде всего, линкером комплексов нуклеоскелета и цитоскелета (LINC).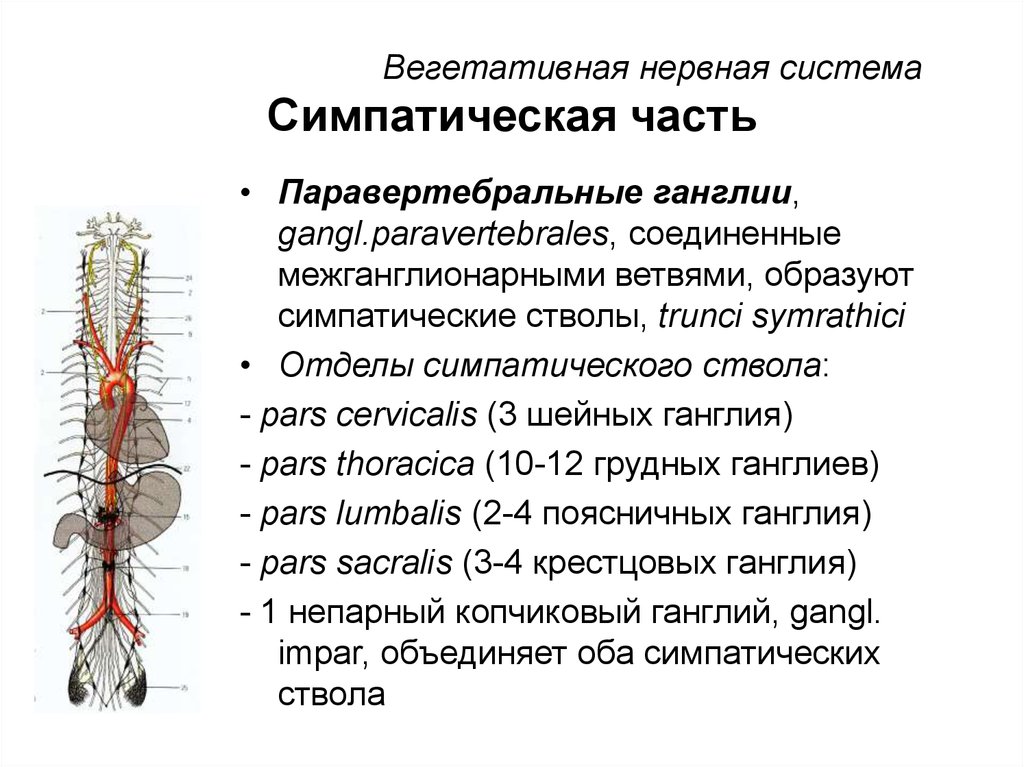
 У мутанта wit12 ядерные пики Ca 2+ снижаются в ответ на экзогенные АФК, но эти пики не коррелируют с разрывом пыльцевых трубок. Это исследование показало, что близость ВЯ к кончику пыльцевой трубки необходима как для ответа на экзогенные АФК, так и для внутреннего ядерного Ca 9.0007 2+ колебания.
У мутанта wit12 ядерные пики Ca 2+ снижаются в ответ на экзогенные АФК, но эти пики не коррелируют с разрывом пыльцевых трубок. Это исследование показало, что близость ВЯ к кончику пыльцевой трубки необходима как для ответа на экзогенные АФК, так и для внутреннего ядерного Ca 9.0007 2+ колебания.Ключевые слова: комплекс ЛИНК; мужская фертильность; ядерный кальций; ядерная оболочка; окончание пыльцевой трубки; активные формы кислорода.
Заявление о конфликте интересов
Авторы не сообщили о потенциальном конфликте интересов.
Цифры
Рисунок 1.
Ca 2+ АФК-зависимая пыльца…
Рисунок 1.
Ca 2+ -зависимый Взрыв пыльцевой трубки, индуцированный АФК, коррелирует с положением ядра.
Рисунок 1.Ca 2+ -зависимый взрыв пыльцевой трубки, индуцированный АФК, коррелирует с положением в ядре.
Рисунок 1.
Ca 2+ АФК-зависимая пыльца…
Рисунок 1.
Ca 2+ -зависимый Взрыв пыльцевой трубки, индуцированный АФК, коррелирует с положением ядра.
Рисунок 1.Ca 2+ -зависимый взрыв пыльцевой трубки, индуцированный АФК, коррелирует с положением в ядре.
Рисунок 2.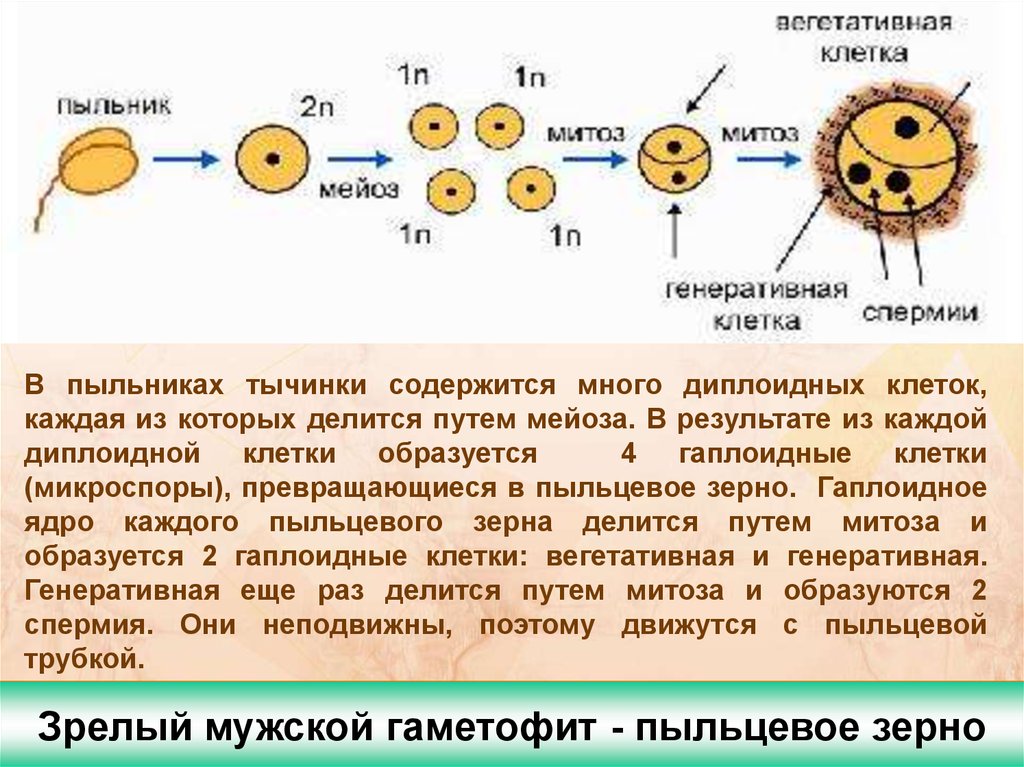
Цитоплазматический Ca 2+ колебания…
Рисунок 2.
Цитоплазматический Ca 2+ флуктуации не нарушены в w12 пыльцевых трубках.
Фигура 2.Цитоплазматический Ca 2+ флуктуации не нарушены в w12 пыльцевых трубках.
Рисунок 2.
Цитоплазматический Ca 2+ колебания…
Рисунок 2.
Цитоплазматический Ca 2+ флуктуации не нарушены в w12 пыльцевых трубках.
Фигура 2. Цитоплазматический Ca 2+ колебания не нарушены в wit12 пыльцевые трубки.
Рисунок 3.
Пыльца ядерная Ca 2+ флуктуации…
Рисунок 3.
Пыльца ядерная Ca 2+ колебания коррелируют с положением ядра.
Рисунок 3.Пыльца ядерная Ca 2+ колебаний коррелируют с положением ядра.
Рисунок 3.
Пыльца ядерная Ca 2+ флуктуации…
Рисунок 3.
Пыльца ядерная Ca 2+ колебания коррелируют с положением ядра.
Рисунок 3. Пыльца ядерная Ca 2+ колебания коррелируют с положением ядра.
Рисунок 4.
Пост-АФК Ca 2+ колебания.
Рисунок 4.
Пост-АФК Ca 2+ колебания.
Рисунок 4.Пост-АФК Са 2+ колебания.
Рисунок 4.
Пост-АФК Ca 2+ колебания.
Рисунок 4.
Пост-АФК Ca 2+ колебания.
Рисунок 4.Post-ROS Ca 2+ колебания.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
SUN закрепляет пыльцевые комплексы WIP-WIT на вегетативной ядерной оболочке и необходим для нацеливания на пыльцевые трубки и фертильности.
Чжоу X, Гроувс Н.Р., Мейер И. Чжоу С и др. J Опытный бот. 2015 декабрь; 66 (22): 7299-307. дои: 10.1093/jxb/erv425. Epub 2015 25 сентября. J Опытный бот. 2015. PMID: 26409047 Бесплатная статья ЧВК.
Продукция апопластных АФК при опылении RbohH и RbohJ у арабидопсиса.
Кая Х., Ивано М., Такеда С., Канаока М.М., Кимура С., Абэ М., Кучицу К. Кая Х и др. Поведение сигналов растений. 2015;10(2):e989050. дои: 10.4161/15592324.2014.989050. Поведение сигналов растений. 2015. PMID: 25751652 Бесплатная статья ЧВК.
NAD(P)H-оксидазы пыльцевых трубок действуют как регулятор скорости, чтобы ослабить колебания скорости роста во время роста поляризованных клеток.
Лассиг Р., Гутермут Т.
, Бей Т.Д., Конрад К.Р., Ромейс Т.
Лассиг Р. и соавт.
Плант Дж. 2014 Апрель; 78 (1): 94-106. doi: 10.1111/tpj.12452. Epub 2014 12 марта.
Завод Дж. 2014.
PMID: 24506280Заводской комплекс ЛИНК у ядерной оболочки.
Tatout C, Evans DE, Vanrobays E, Probst AV, Graumann K. Tatout C и соавт. Хромосомный Рез. 2014 июнь;22(2):241-52. дои: 10.1007/s10577-014-9419-7. Хромосомный Рез. 2014. PMID: 24801343 Обзор.
СВЯЗЬ ядерной оболочки с архитектурой сперматозоидов.
Манфревола Ф., Гийу Ф., Фазано С., Пьерантони Р., Кьянезе Р. Манфревола Ф. и др. Гены (Базель). 2021 27 апреля; 12 (5): 658. doi: 10.3390/genes12050658. Гены (Базель). 2021. PMID: 33925685 Бесплатная статья ЧВК. Обзор.
 , Бей Т.Д., Конрад К.Р., Ромейс Т.
Лассиг Р. и соавт.
Плант Дж. 2014 Апрель; 78 (1): 94-106. doi: 10.1111/tpj.12452. Epub 2014 12 марта.
Завод Дж. 2014.
PMID: 24506280
, Бей Т.Д., Конрад К.Р., Ромейс Т.
Лассиг Р. и соавт.
Плант Дж. 2014 Апрель; 78 (1): 94-106. doi: 10.1111/tpj.12452. Epub 2014 12 марта.
Завод Дж. 2014.
PMID: 24506280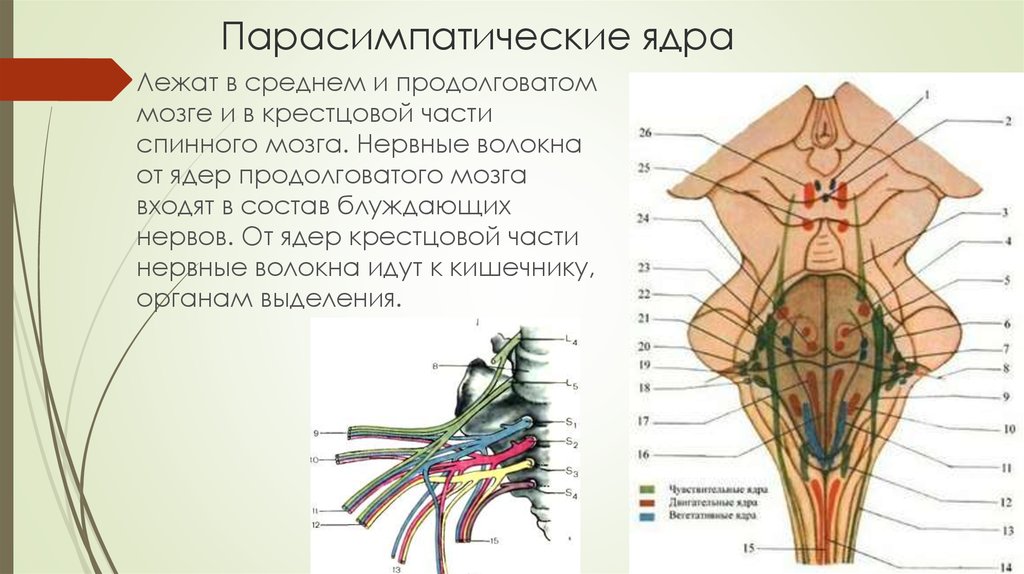
Посмотреть все похожие статьи
Цитируется
Мутация компонента комплекса ядерных пор, aladin1, нарушает асимметричное деление клеток у Zea mays (кукуруза).
Лучший NB, Addo-Quaye C, Kim BS, Weil CF, Schulz B, Johal G, Dilkes BP. Бест Н.Б. и др. G3 (Бетесда). 2021 14 июля; 11 (7): jkab106. doi: 10.1093/g3journal/jkab106. G3 (Бетесда). 2021. PMID: 36351283 Бесплатная статья ЧВК.
Различные роли белков KASH SINE1 и SINE2 в реорганизации актина замыкающих клеток, колебаниях кальция и ремоделировании вакуолей.
Биль А., Мозер М., Гровс Н.Р., Мейер И. Биль А. и соавт. Фронт завод науч. 2022 6 мая; 13:784342. doi: 10.3389/fpls.2022.784342. Электронная коллекция 2022.
Фронт завод науч. 2022.
PMID: 35599883
Бесплатная статья ЧВК.Связанное со стрессом репрограммирование развития у протонем мхов путем синтетической активации общего пути симбиоза.
Клейст Т.Дж., Бортолаццо А., Кейзер З.П., Перера А.М., Ирвинг Т.Б., Венкатешваран М., Атанджауи Ф., Танг Р.Дж., Маеда Дж., Картрайт Х.Н., Кристиансон М.Л., Лемо П.Г., Луан С., Ане Дж.М. Клейст Т.Дж. и соавт. iНаука. 2022 11 января; 25 (2): 103754. doi: 10.1016/j.isci.2022.103754. Электронная коллекция 2022 18 февраля. iНаука. 2022. PMID: 35146383 Бесплатная статья ЧВК.
Глоссарий структур растительных клеток: текущие идеи и вопросы на будущее.
Канг Б.Х., Андерсон К.Т., Аримура С.И., Байер Э., Безанилла М., Ботелла М.
А., Брандиззи Ф., Берч-Смит Т.М., Чепмен К.Д., Дюнсер К., Гу И., Джайлай И., Кирххофф Х., Отеги М.С., Росадо А., Тан Ю., Кляйне-Вен Дж., Ван П., Золман Б.К.
Канг Б.Х. и др.
Растительная клетка. 2022 янв. 20;34(1):10-52. doi: 10.1093/plcell/koab247.
Растительная клетка. 2022.
PMID: 34633455
Бесплатная статья ЧВК.
Обзор.Регуляция и физиологическое значение формы ядра у растений.
Гото К., Хара-Нисимура И., Тамура К. Гото С и др. Фронт завод науч. 2021 10 июня; 12:673905. doi: 10.3389/fpls.2021.673905. Электронная коллекция 2021. Фронт завод науч. 2021. PMID: 34177991 Бесплатная статья ЧВК. Обзор.
 Фронт завод науч. 2022.
PMID: 35599883
Бесплатная статья ЧВК.
Фронт завод науч. 2022.
PMID: 35599883
Бесплатная статья ЧВК.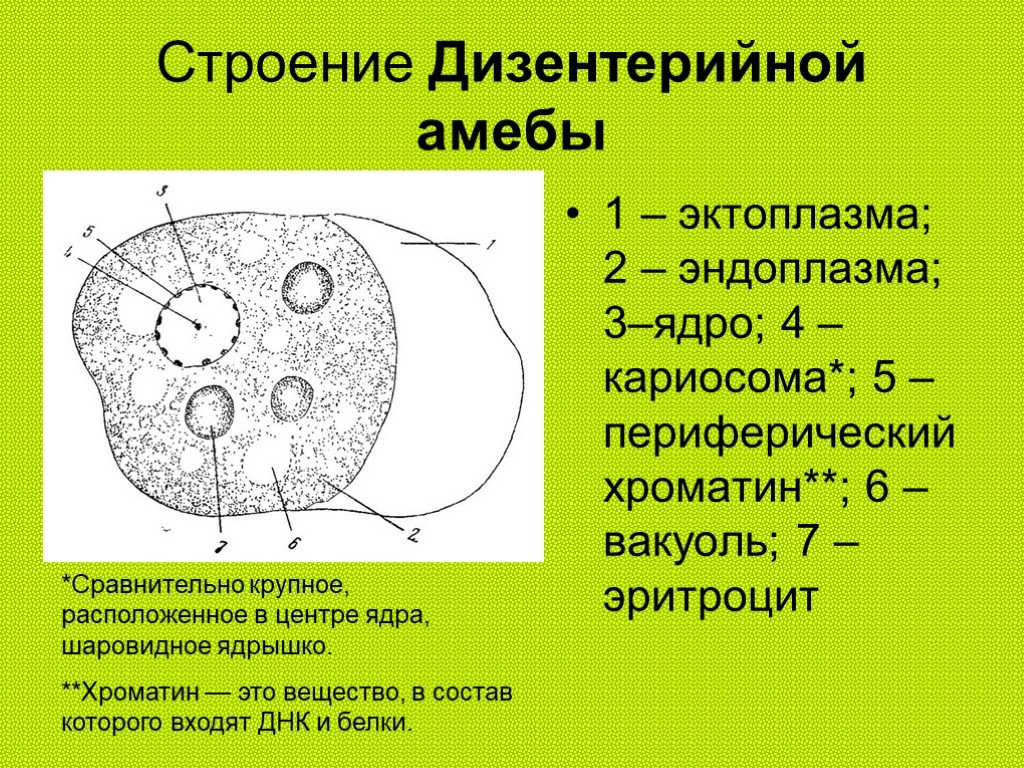 А., Брандиззи Ф., Берч-Смит Т.М., Чепмен К.Д., Дюнсер К., Гу И., Джайлай И., Кирххофф Х., Отеги М.С., Росадо А., Тан Ю., Кляйне-Вен Дж., Ван П., Золман Б.К.
Канг Б.Х. и др.
Растительная клетка. 2022 янв. 20;34(1):10-52. doi: 10.1093/plcell/koab247.
Растительная клетка. 2022.
PMID: 34633455
Бесплатная статья ЧВК.
Обзор.
А., Брандиззи Ф., Берч-Смит Т.М., Чепмен К.Д., Дюнсер К., Гу И., Джайлай И., Кирххофф Х., Отеги М.С., Росадо А., Тан Ю., Кляйне-Вен Дж., Ван П., Золман Б.К.
Канг Б.Х. и др.
Растительная клетка. 2022 янв. 20;34(1):10-52. doi: 10.1093/plcell/koab247.
Растительная клетка. 2022.
PMID: 34633455
Бесплатная статья ЧВК.
Обзор.Просмотреть все статьи «Цитируется по»
Рекомендации
- Bone CR, Starr DA. События ядерной миграции на протяжении всей разработки. Дж. Клеточные науки. 2016; 129:1951–1961.
—
ЧВК
—
пабмед
- Bone CR, Starr DA.
- Старр Д.А., Фридольфссон Х.Н. Взаимодействия между ядрами и цитоскелетом опосредованы мостиками SUN-KASH между ядерной оболочкой. Annu Rev Cell Dev Biol. 2010;26:421–444. — ЧВК — пабмед
- Чжоу С. , Грауманн К., Эванс Д.Э. и соавт. Новые заводские мосты SUN-KASH участвуют в закреплении RanGAP и определении формы ядер. Джей Селл Биол. 2012;196:203–211.
—
ЧВК
—
пабмед
- Чжоу С.
- Чжоу С., Грауманн К., Виртмюллер Л. и др. Идентификация уникальных белков ядерной оболочки, взаимодействующих с SUN, с разнообразными функциями у растений. Джей Селл Биол. 2014; 205: 677–692. — ЧВК — пабмед
- Пуле А. , Пробст А.В., Грауманн К. и соавт. Изучение эволюции белков ядерной оболочки растений. Ядро. 2017; 8:46–59.
—
ЧВК
—
пабмед
- Пуле А.
 События ядерной миграции на протяжении всей разработки. Дж. Клеточные науки. 2016; 129:1951–1961.
—
ЧВК
—
пабмед
События ядерной миграции на протяжении всей разработки. Дж. Клеточные науки. 2016; 129:1951–1961.
—
ЧВК
—
пабмед , Грауманн К., Эванс Д.Э. и соавт. Новые заводские мосты SUN-KASH участвуют в закреплении RanGAP и определении формы ядер. Джей Селл Биол. 2012;196:203–211.
—
ЧВК
—
пабмед
, Грауманн К., Эванс Д.Э. и соавт. Новые заводские мосты SUN-KASH участвуют в закреплении RanGAP и определении формы ядер. Джей Селл Биол. 2012;196:203–211.
—
ЧВК
—
пабмед , Пробст А.В., Грауманн К. и соавт. Изучение эволюции белков ядерной оболочки растений. Ядро. 2017; 8:46–59.
—
ЧВК
—
пабмед
, Пробст А.В., Грауманн К. и соавт. Изучение эволюции белков ядерной оболочки растений. Ядро. 2017; 8:46–59.
—
ЧВК
—
пабмедТипы публикаций
термины MeSH
вещества
Грантовая поддержка
Эта работа была поддержана грантом Национального научного фонда для IM [NSF-1613501].
Конформация и движение вегетативного ядра пыльцевой трубки покрытосеменных: ассоциация с актиновым цитоскелетом | Журнал клеточной науки
Пропустить пункт назначения навигации
СТАТЬЯ В ЖУРНАЛЕ|
01 июня 1989 г.
Дж. ХЕСЛОП-ХАРРИСОН,
Ю. ХЕСЛОП-ХАРРИСОН
Информация об авторе и статье
Полученная редакция: 04 января 1989 г.
Принял: 16 марта 1989
Номер в сети: 1477-9137
Номер для печати: 0021-9533
https://doi.org/10.1242/jcs.93.2.299
История статьи
Редакция Получено:
4 января 1989 г.
Принято:
16 марта 1989 г.
- Разделенный экран
- Взгляды
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Делиться
- Фейсбук
- Твиттер
- MailTo
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
Цитата
Дж.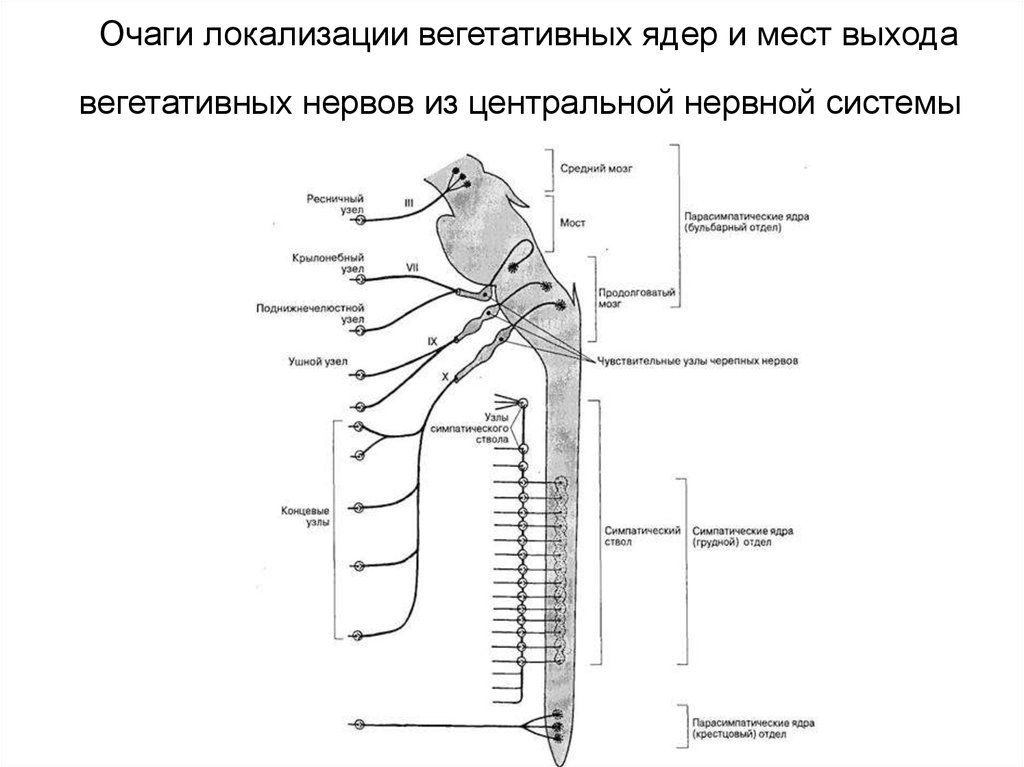 ХЕСЛОП-ХАРРИСОН, Ю. ХЕСЛОП-ХАРРИСОН; Конформация и движение вегетативного ядра пыльцевой трубки покрытосеменных: ассоциация с актиновым цитоскелетом. J Cell Sci 1 июня 1989 г.; 93 (2): 299–308. Дои: https://doi.org/10.1242/jcs.93.2.299
ХЕСЛОП-ХАРРИСОН, Ю. ХЕСЛОП-ХАРРИСОН; Конформация и движение вегетативного ядра пыльцевой трубки покрытосеменных: ассоциация с актиновым цитоскелетом. J Cell Sci 1 июня 1989 г.; 93 (2): 299–308. Дои: https://doi.org/10.1242/jcs.93.2.299
Скачать файл цитаты:
- Рис (Зотеро)
- Диспетчер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- КонецПримечание
- РефВоркс
- Бибтекс
Расширенный поиск
Актин присутствует в цитоплазме вегетативной клетки пыльцы покрытосеменных в виде многочисленных веретеновидных, колосовидных или тороидальных телец, а также в виде оболочки, обволакивающей вегетативное ядро. При активации после гидратации компактные цитоплазматические тельца превращаются в мотки протяженных фибрилл, и в цитоплазме начинаются циркуляторные движения.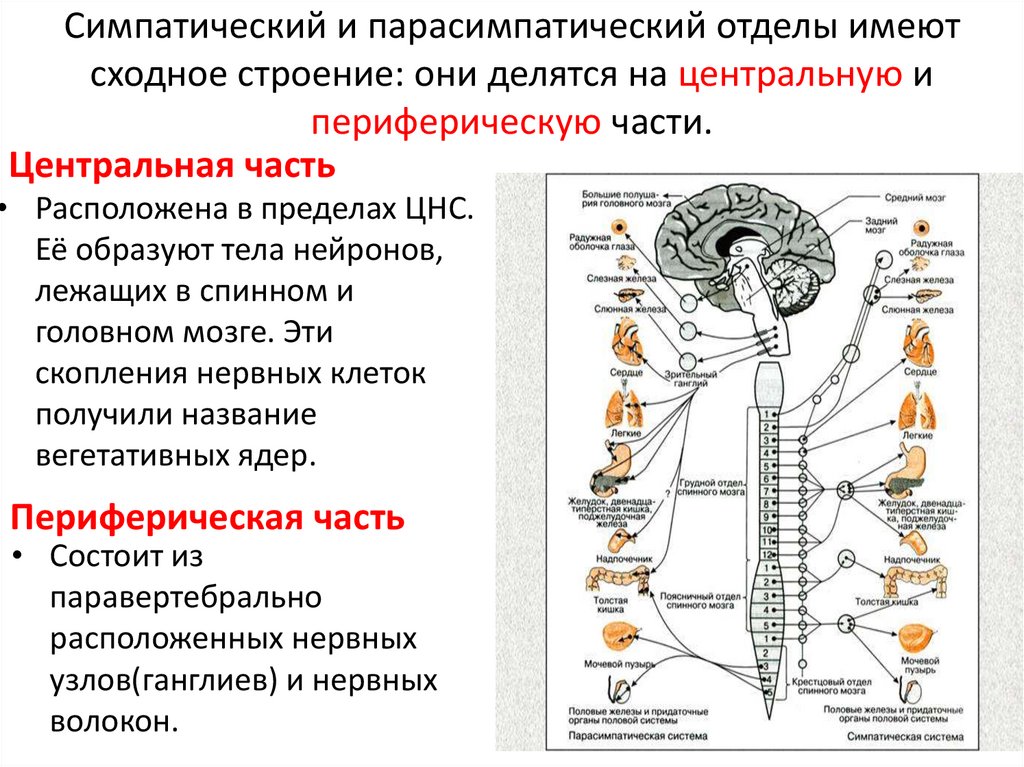 На протяжении всего этого периода вегетативное ядро с фибриллярным актином, связанным теперь с поверхностью, претерпевает непрерывное изменение формы. В вытягивающейся трубке после прорастания актиновый цитоскелет состоит из многочисленных преимущественно продольно ориентированных фибрилл. После входа в трубку вегетативное ядро остается связанным с фибриллами, обычно сильно вытягивающимися в длину и образующими утонченные, часто заостренные отростки. Наблюдаемые конформации, которые постоянно меняются, позволяют предположить, что при прохождении через трубку к вегетативному ядру применяются различные локальные напряжения. Цитохалазин D разрушает систему актиновых фибрилл и вызывает быстрое сокращение ядра, в то же время устраняя эластические растяжения ядерной оболочки. Ядра, изолированные физически из незафиксированных трубочек, также сокращаются в длину по мере отсоединения фибриллярных компонентов цитоскелета. Эти данные указывают на то, что движение вегетативного ядра зависит от локальных ассоциаций ядерной оболочки с актиновым цитоскелетом вегетативной клетки.
На протяжении всего этого периода вегетативное ядро с фибриллярным актином, связанным теперь с поверхностью, претерпевает непрерывное изменение формы. В вытягивающейся трубке после прорастания актиновый цитоскелет состоит из многочисленных преимущественно продольно ориентированных фибрилл. После входа в трубку вегетативное ядро остается связанным с фибриллами, обычно сильно вытягивающимися в длину и образующими утонченные, часто заостренные отростки. Наблюдаемые конформации, которые постоянно меняются, позволяют предположить, что при прохождении через трубку к вегетативному ядру применяются различные локальные напряжения. Цитохалазин D разрушает систему актиновых фибрилл и вызывает быстрое сокращение ядра, в то же время устраняя эластические растяжения ядерной оболочки. Ядра, изолированные физически из незафиксированных трубочек, также сокращаются в длину по мере отсоединения фибриллярных компонентов цитоскелета. Эти данные указывают на то, что движение вегетативного ядра зависит от локальных ассоциаций ядерной оболочки с актиновым цитоскелетом вегетативной клетки.